
Cortical Activation in Mental Rotation and the Role of the Corpus Callosum: Observations in Healthy Subjects and Split-Brain Patients

 and
and
Abstract
:1. Introduction
2. Materlials and Methods
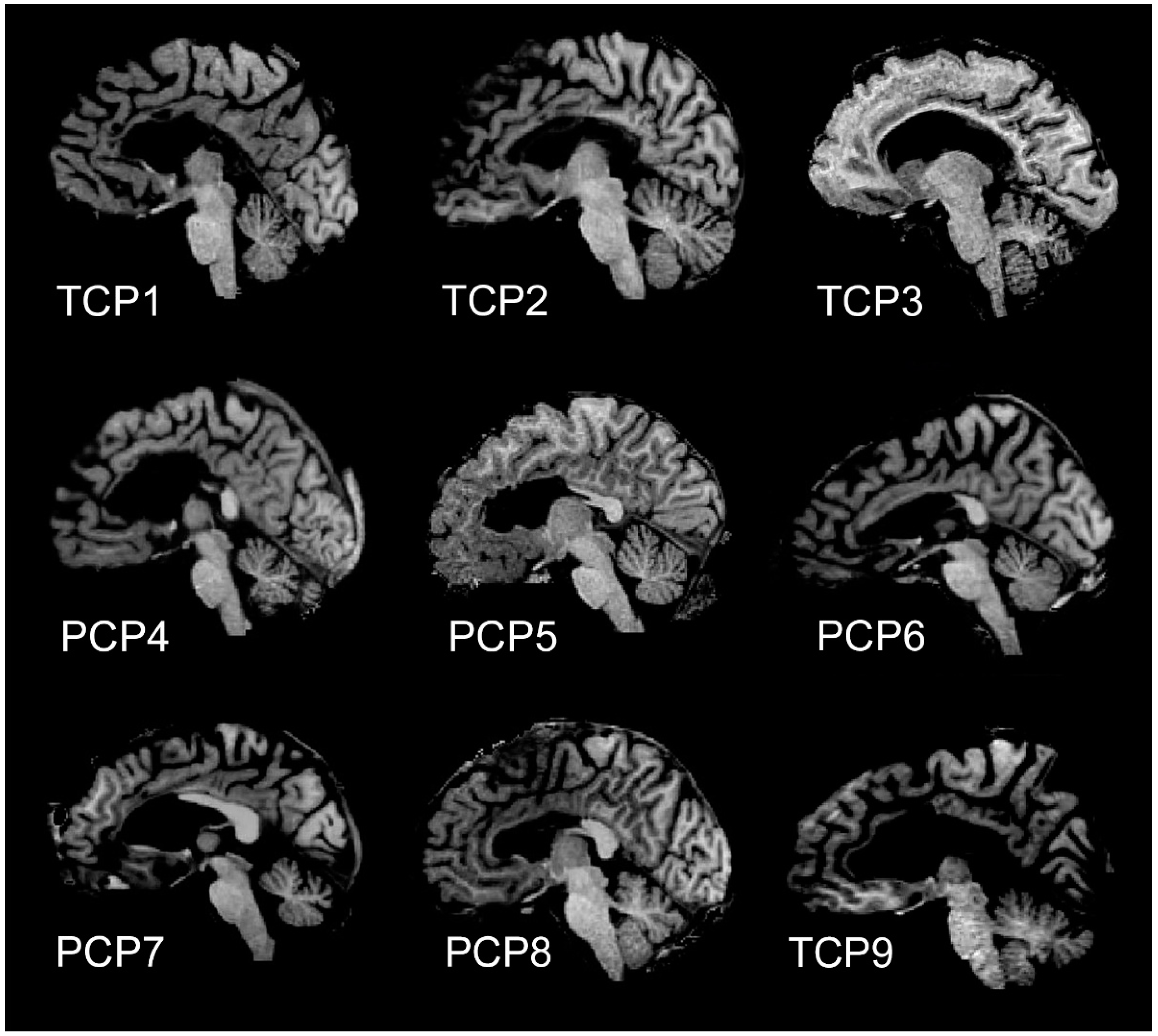
2.1. Participants
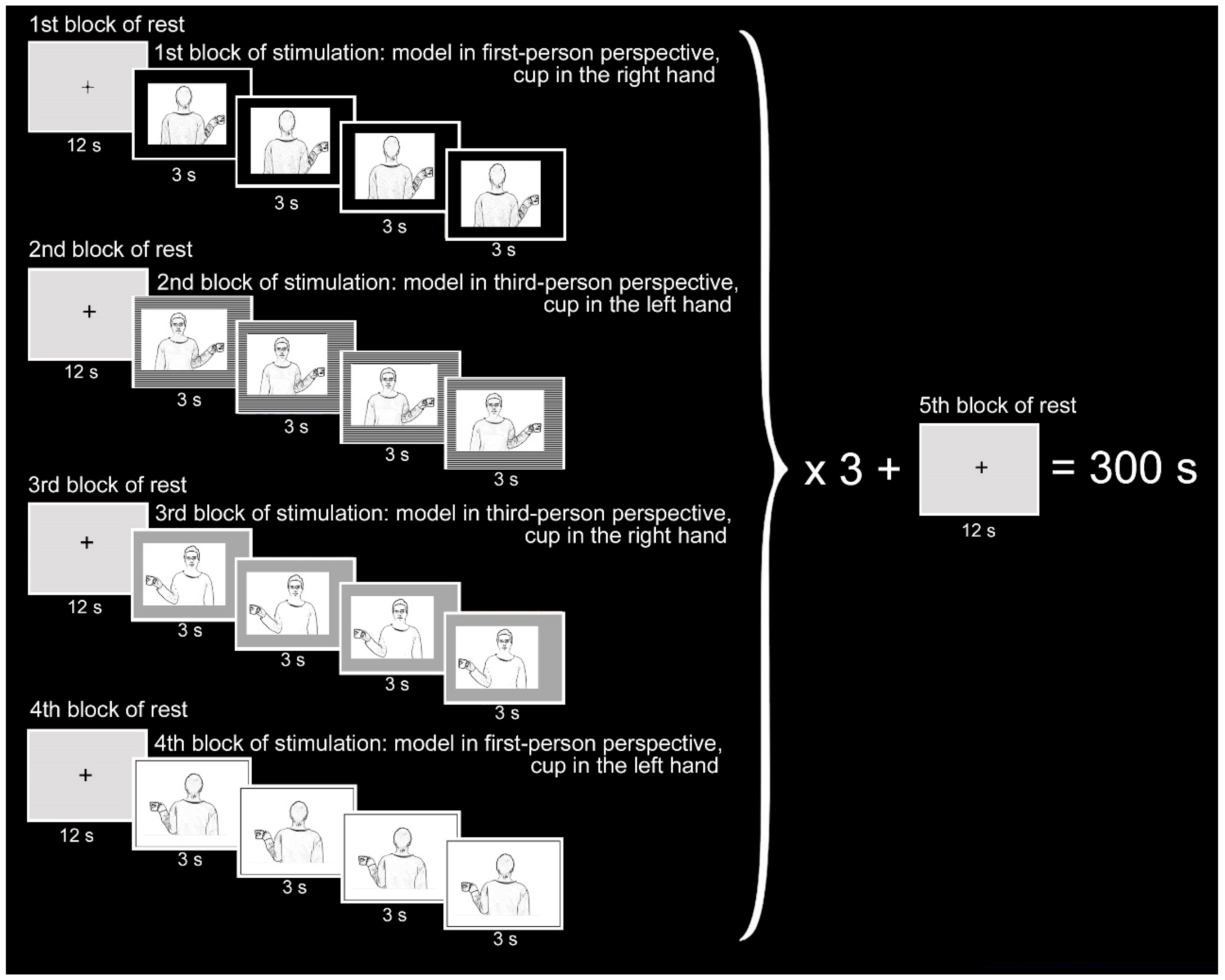
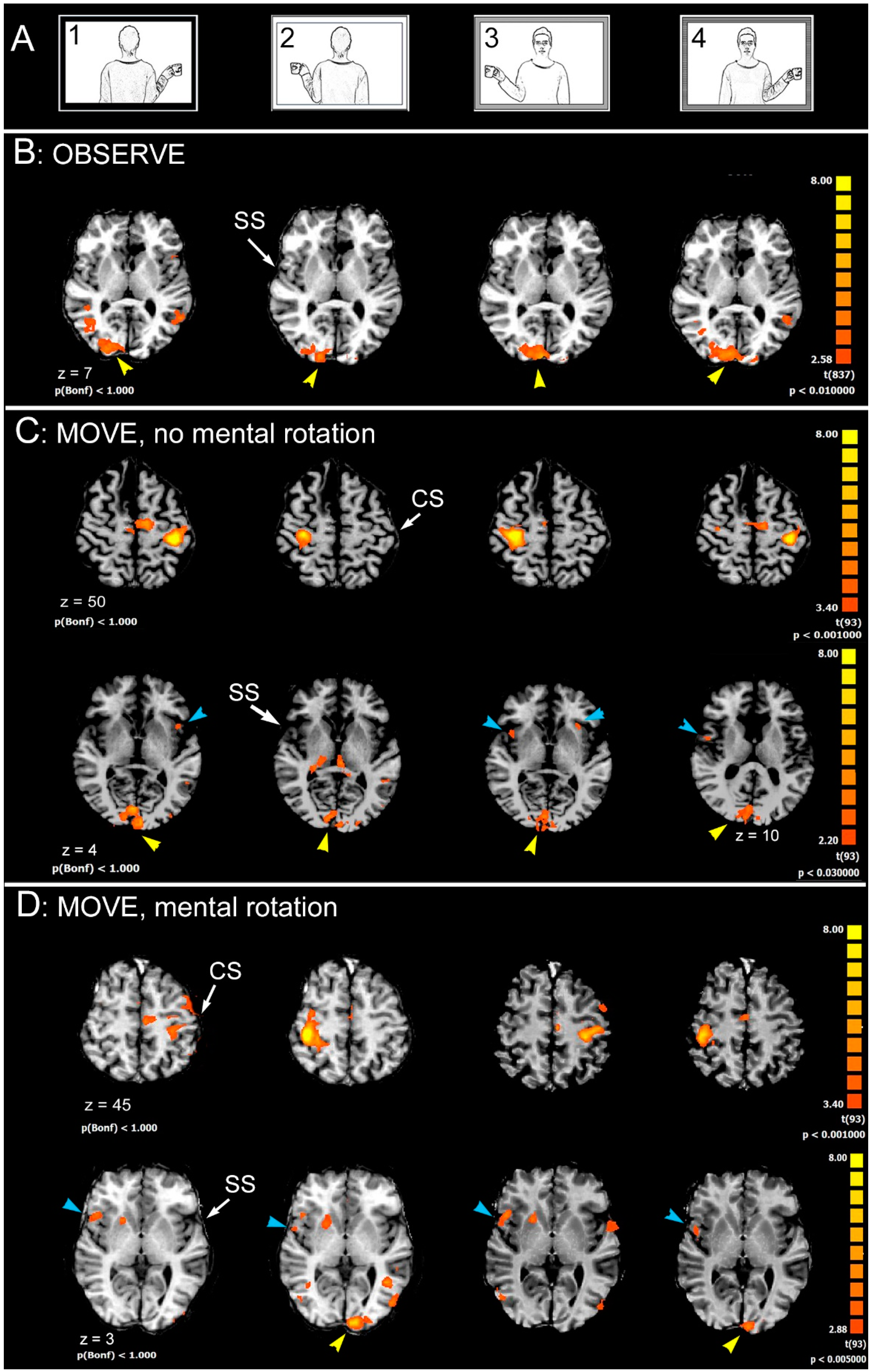
2.2. Stimuli and Task
- Observation (OBSERVE): the subject was required to carefully observe the pictures. The instruction was: “Please, simply look at the pictures”.
- Hand movement (MOVE): the subject observed the pictures and was requested to move their right (or left) hand, when the cup was in the model’s right (or left) hand. The instruction was “Please, as soon as you see the picture, move your right hand if the cup is in the model’s right hand, or move your left hand if the cup is in the model’s left hand”.
- Thinking (THINK): this condition, being more difficult to control, could be analyzed only in control subjects and in one patient. Subjects were asked to think in which hand of the model the cup was held, in their judice. The instruction was “Please, as soon as you see the picture, think “right” if the cup is in the model’s right hand, or think “left” if the cup is in the model’s left hand”.
2.3. Functional MRI Stimulation Protocol
2.4. Functional MRI Data Acquisition
- anatomical 3-planes localizer (2D SPGR, TR 120 ms, TE 15 ms, Flip Angle 70°, FOV 23 × 23 cm, slice thickness 5 mm, Matrix 256 × 256, 1 Nex, scan time 31 s);
- 3D data set (IR Prep Fast SPGR; TR 15.2 ms, TE 6.9 ms, TI 500 ms, Flip Angle 15°, FOV 29 × 29 cm, slice thickness 1 mm, Matrix 288 × 288, 1 Nex, scan time 8:20 min);
- 20 contiguous 5-mm-thick axial (or oblique) functional images, single-shot T2*-weighted gradient-echo EPI sequence (TR 3000 ms, TE 60 ms, Flip Angle 90°, FOV 28 × 21 cm, Matrix 96 × 64, 1 Nex, scan time 5:12 min);
- high-resolution axial (or oblique) anatomical images from 20 selected planes (2D SPGR, TR 100 ms, TE 12 ms, Flip Angle 70°, FOV 28 × 21 cm, slice thickness 5 mm, Matrix 256 × 256, 1 Nex, scan time 2:25 min) to superimpose functional activation images onto anatomical landmarks, allowing to show blood vessels, possible sources of BOLD signals.
2.5. Functional MRI Data Analysis
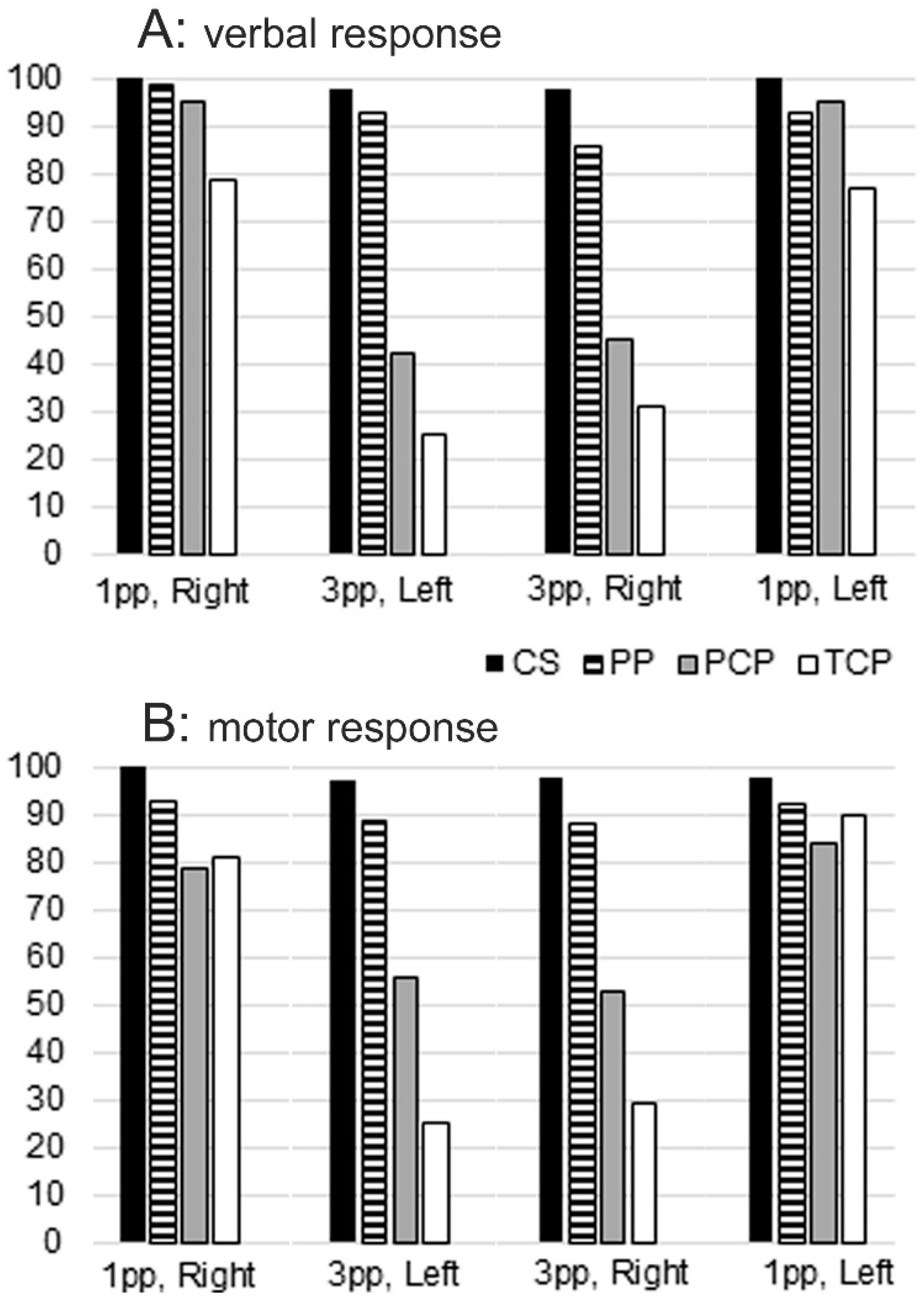
3. Results
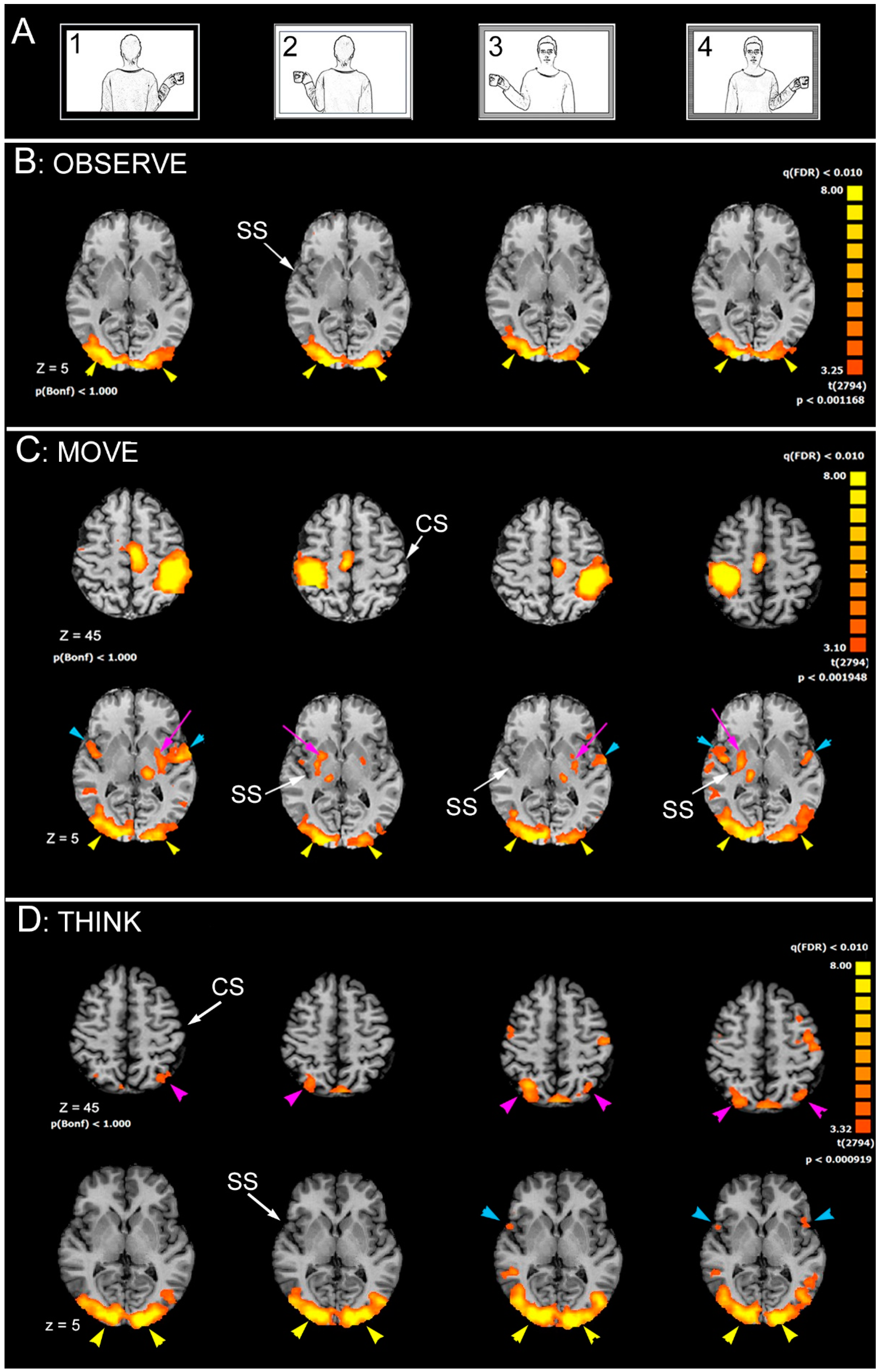
- c1 = model in 1st person, cup in the right hand
- c2 = model in 1st person, cup in the left hand
- c3 = model in 3rd person, cup in the right hand
- c4 = model in 3rd person, cup in the left hand
4. Control Subjects
4.1. Brain Areas Activated during OBSERVE Task
4.2. Brain Areas Activated during MOVE Task
4.3. Brain Areas Activated during THINK Task
4.4. Callosotomized Patients
4.4.1. Brain Areas Activated during OBSERVE Task
4.4.2. Brain Areas Activated during MOVE Task
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corballis, M.C. Mental Rotation and the Right Hemisphere. Brain Lang. 1997, 57, 100–121. [Google Scholar] [CrossRef]
- Kosslyn, S.M. Aspects of a cognitive neuroscience of mental imagery. Science 1988, 240, 1621–1626. [Google Scholar] [CrossRef] [PubMed]
- Tomasino, B.; Borroni, P.; Isaja, A.; Rumiati, R.I. The role of the primary motor cortex in mental rotation: A TMS study. Cogn. Neuropsychol. 2005, 22, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Zacks, J.M.; Michelon, P. Transformations of Visuospatial Images. Behav. Cogn. Neurosci. Rev. 2005, 4, 96–118. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, M. Spatial representation in mind and brain. In What Deficits Reveal about the Human Mind/Brain: A Handbook of Cognitive Neuropsychology; Rapp, B., Ed.; Psychology Press: Philadelphia, PA, USA, 2001; pp. 101–132. [Google Scholar]
- Shepard, R.N.; Metzler, J. Mental Rotation of Three-Dimensional Objects. Science 1971, 171, 701–703. [Google Scholar] [CrossRef] [Green Version]
- Cooper, L.A.; Shepard, R.N. The time required to prepare for a rotated stimulus. Memory Cogn. 1973, 1, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Zacks, J.M. Neuroimaging Studies of Mental Rotation: A Meta-analysis and Review. J. Cogn. Neurosci. 2008, 20, 1–19. [Google Scholar] [CrossRef]
- Cooper, L.A.; Shepard, R.N. Mental transformation in the identification of left and right hands. J. Exp. Psychol. Hum. Percept. Perform. 1975, 1, 48–56. [Google Scholar] [CrossRef]
- Parsons, L.M. Temporal and kinematic properties of motor behavior reflected in mentally simulated action. J. Exp. Psychol. Hum. Percept. Perform. 1994, 20, 709–730. [Google Scholar] [CrossRef]
- de Lange, F.P.; Helmich, R.C.; Toni, I. Posture influences motor imagery: An fMRI study. NeuroImage 2006, 33, 609–617. [Google Scholar] [CrossRef]
- Jansen, P.; Kaltner, S. Object-based and egocentric mental rotation performance in older adults: The importance of gender differences and motor ability. Aging, Neuropsychol. Cogn. 2013, 21, 296–316. [Google Scholar] [CrossRef]
- Linn, M.C.; Petersen, A.C. Emergence and Characterization of Sex Differences in Spatial Ability: A Meta-Analysis. Child Dev. 1985, 56, 1479. [Google Scholar] [CrossRef]
- Voyer, D.; Voyer, S.; Bryden, M.P. Magnitude of sex differences in spatial abilities: A meta-analysis and consideration of criticalvariables. Psychol. Bull. 1995, 117, 250. [Google Scholar] [CrossRef]
- Parsons, L.M. Superior parietal cortices and varieties of mental rotation. Trends Cogn. Sci. 2003, 7, 515–517. [Google Scholar] [CrossRef]
- Tomasino, B.; Vorano, L.; Skrap, M.; Gigli, G.; Rumiati, R.I. Effects of strategies on mental rotation performed by unilateral brain damaged patients. Cortex 2004, 40, 197–199. [Google Scholar] [CrossRef]
- Serrati, C.; Finocchi, C.; Calautti, C.; Bruzzone, G.L.; Colucci, M.; Gandolfo, C.; Del Sette, M.; Lantieri, P.B.; Favale, E. Absence of Hemispheric Dominance for Mental Rotation Ability: A Transcranial Doppler Study. Cortex 2000, 36, 415–425. [Google Scholar] [CrossRef]
- Nishizawa, H.; Kimura, T.; Goh, A.-C. The effect of different imitation models on the accuracy and speed of imitation of movement. J. Phys. Ther. Sci. 2015, 27, 3417–3420. [Google Scholar] [CrossRef] [Green Version]
- Meltzoff, A.N.; Moore, M.K. Imitation, memory, and the representation of persons. Infant Behav. Dev. 1994, 17, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Pierpaoli, C.; Ferrante, L.; Manzoni, T.; Fabri, M. Anatomical or mirror mode imitation? A behavioral approach. Arch. Ital. Biol. 2014, 152, 20–31. [Google Scholar] [PubMed]
- Pierpaoli, C.; Foschi, N.; Cagnetti, C.; Ferrante, L.; Manzoni, T.; Polonara, G.; Fabri, M. Imitation strategies in callo-sotomeized patients. Arch. Ital. Biol. 2018, 156, 12–26. [Google Scholar] [PubMed]
- Sansonetti, R.; Pierpaoli, C.; Ferrante, L.; Fabri, M.; Nardi, B. Imitation strategies in subjects with schizophrenia: A behavioural approach. Arch. Ital. Biol. 2020, 158, 3–16. [Google Scholar] [CrossRef]
- Fujino, J.; Takahashi, H.; Miyata, J.; Sugihara, G.; Kubota, M.; Sasamoto, A.; Fujiwara, H.; Aso, T.; Fukuyama, H.; Murai, T. Impaired empathic abilities and reduced white matter integrity in schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 48, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahab, S.; Stefanik, L.; Foussias, G.; Lai, M.-C.; Anderson, K.K.; Voineskos, A.N. Sex and Diffusion Tensor Imaging of White Matter in Schizophrenia: A Systematic Review Plus Meta-analysis of the Corpus Callosum. Schizophr. Bull. 2017, 44, 203–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierpaoli, C.; Ferrante, L.; Foschi, N.; Lattanzi, S.; Sansonetti, R.; Polonara, G.; Mari, M.; Nardi, B.; Fabri, M. Mental Rotation Ability: Right or Left Hemisphere Competence? What We Can Learn from Callosotomized and Psychotic Patients. Symmetry 2020, 12, 1137. [Google Scholar] [CrossRef]
- Fabri, M.; Ombrosi, M.; Goushi, M.; Pierpaoli, C.; Foschi, N.; Lattanzi, S.; Polonara, G. Cortical activation in mental rotation: Observations in healthy subjects and split-brain patients. In Proceedings of the 69th Congress of the Italian Physiological Society, Firenze, Italy, 19–21 September 2018. [Google Scholar]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Talairach, J.; Turnoux, P. Co-Planar Stereotaxic Atlas of the Human Brain; Thieme Medical Publishers: New York, NY, USA, 2008. [Google Scholar]
- Goebel, R. Brainvoyager: A program for analyzing and visualizing functional and structural magnetic resonance data sets. NeuroImage 1996, 3, S604. [Google Scholar] [CrossRef]
- Milivojevic, B.; Hamm, J.P.; Corballis, M.C. Functional Neuroanatomy of Mental Rotation. J. Cogn. Neurosci. 2009, 21, 945–959. [Google Scholar] [CrossRef]
- Thirioux, B.; Mercier, M.R.; Jorland, G.; Berthoz, A.; Blanke, O. Mental Imagery of Self-Location during Spontaneous and Active Self-Other Interactions: An Electrical Neuroimaging Study. J. Neurosci. 2010, 30, 7202–7214. [Google Scholar] [CrossRef] [Green Version]
- Tomasino, B.; Gremese, M. Effects of Stimulus Type and Strategy on Mental Rotation Network: An Activation Likelihood Estimation Meta-Analysis. Front. Hum. Neurosci. 2016, 9, 693. [Google Scholar] [CrossRef] [Green Version]
- Jackson, P.L.; Meltzoff, A.N.; Decety, J. Neural circuits involved in imitation and perspective-taking. NeuroImage 2006, 31, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Swoden, S.; Catmur, C. The Role of the Right Temporoparietal Junction in the Control of Imitation. Cereb. Cortex 2015, 25, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Mengotti, P.; Corradi-Dell’Acqua, C.; Rumiati, R.I. Imitation components in the human brain: An fMRI study. NeuroImage 2012, 59, 1622–1630. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, A.; Króliczak, G. Left extrastriate body area is sensitive to the meaning of symbolic gesture: Evidence from fMRI repetition suppression. Sci. Rep. 2016, 6, 31064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengotti, P.; Ticini, L.F.; Waszak, F.; Schütz-Bosbach, S.; Rumiati, R.I. Imitating others’ actions: Transcraniam magnetic stimulation on the parietal opercula reveals the processes underlying automatic imitation. Eur. J. Neurosci. 2013, 37, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.-P.; Cho, K.-H.; Yeh, C.-H.; Chou, K.-H.; Chen, J.-H.; Lin, C.-P. Probabilistic topography of human corpus callosum using cytoarchitectural parcellation and high angular resolution diffusion imaging tractography. Hum. Brain Mapp. 2009, 30, 3172–3187. [Google Scholar] [CrossRef] [PubMed]
- Fabri, M.; Polonara, G. Functional Topography of Human Corpus Callosum: An FMRI Mapping Study. Neural Plast. 2013, 2013, 251308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonara, G.; Mascioli, G.; Foschi, N.; Salvolini, U.; Pierpaoli, C.; Manzoni, T.; Fabri, M.; Barbaresi, P. Further evidence for the topography and connectivity of the corpus callosum: An FMRI study of patients with partial callosal resection. J. Neuroimaging 2014, 25, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Mascioli, G.; Berlucchi, G.; Pierpaoli, C.; Salvolini, U.; Barbaresi, P.; Fabri, M.; Polonara, G. Functional MRI cortical activations from unilateral tactile-taste stimulations of the tongue. Physiol. Behav. 2015, 151, 221–229. [Google Scholar] [CrossRef]
- Fabri, M.; Polonara, G.; Quattrini, A.; Salvolini, U.; Del Pesce, M.; Manzoni, T. Role of the corpus callosum in the somatosensory activation of the ipsilateral cerebral cortex: An fMRI study of callosotomized patients. Eur. J. Neurosci. 1999, 11, 3983–3994. [Google Scholar] [CrossRef]
- Frazier, T.W.; Hardan, A.Y. A Meta-Analysis of the Corpus Callosum in Autism. Biol. Psychiatry 2009, 66, 935–941. [Google Scholar] [CrossRef] [Green Version]
- Paul, L.K. Developmental malformation of the corpus callosum: A review of typical callosal development and examples of developmental disorders with callosal involveme. J. Neurodevelop. Disord. 2011, 3, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Frederiksen, K.S. Corpus callosum in aging and dementia. Dan. Med. J. 2013, 60, B4721. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Subject | Gender | Age | Handedness (Oldfield Score) | fMRI | DTI |
|---|---|---|---|---|---|
| CS1 | F | 33 | Right (14) | yes | yes |
| CS2 | F | 27 | Right (11) | yes | yes |
| CS3 | M | 24 | Right (23) | yes | yes |
| CS4 | M | 25 | Right (15) | yes | yes |
| CS5 | F | 25 | Right (20) | yes | yes |
| CS6 | M | 27 | Right (12) | yes | yes |
| CS7 | F | 27 | Right (12) | yes | yes |
| CS8 | M | 24 | Right (12) | yes | yes |
| CS9 | M | 33 | Left (45) | Yes | yes |
| CS10 | F | 25 | Right (15) | Yes | yes |
| Patient | Gender | Age | IQ | Handedness (Oldfield Score) | Callosotomy | fMRI | DTI |
|---|---|---|---|---|---|---|---|
| TCP1 | M | 49 | 81 | Right (10) | total | yes | yes |
| TCP2 | M | 39 | 83 | Right (21) | total | yes | yes |
| TCP3 | F | 37 | 70 | Right (10) | total | yes | yes |
| PCP4 | M | 45 | 70 | Right (10) | partial anterior | yes | yes |
| PCP5 | F | 40 | 70 | Right (10) | partial anterior | yes | yes |
| PCP6 | M | 49 | 87 | Right (10) | partial anterior | yes | yes |
| PCP7 | M | 55 | 80 | Right (10) | partial anterior | yes | yes |
| PCP8 | M | 45 | 70 | Right (10) | partial anterior | yes | yes |
| TCP9 | F | 57 | 93 | Right (10) | total | yes | yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pierpaoli, C.; Ghoushi, M.; Foschi, N.; Lattanzi, S.; Fabri, M.; Polonara, G. Cortical Activation in Mental Rotation and the Role of the Corpus Callosum: Observations in Healthy Subjects and Split-Brain Patients. Symmetry 2021, 13, 1953. https://doi.org/10.3390/sym13101953
Pierpaoli C, Ghoushi M, Foschi N, Lattanzi S, Fabri M, Polonara G. Cortical Activation in Mental Rotation and the Role of the Corpus Callosum: Observations in Healthy Subjects and Split-Brain Patients. Symmetry. 2021; 13(10):1953. https://doi.org/10.3390/sym13101953
Chicago/Turabian StylePierpaoli, Chiara, Mojgan Ghoushi, Nicoletta Foschi, Simona Lattanzi, Mara Fabri, and Gabriele Polonara. 2021. "Cortical Activation in Mental Rotation and the Role of the Corpus Callosum: Observations in Healthy Subjects and Split-Brain Patients" Symmetry 13, no. 10: 1953. https://doi.org/10.3390/sym13101953
APA StylePierpaoli, C., Ghoushi, M., Foschi, N., Lattanzi, S., Fabri, M., & Polonara, G. (2021). Cortical Activation in Mental Rotation and the Role of the Corpus Callosum: Observations in Healthy Subjects and Split-Brain Patients. Symmetry, 13(10), 1953. https://doi.org/10.3390/sym13101953