
Are We Right about the Right TPJ? A Review of Brain Stimulation and Social Cognition in the Right Temporal Parietal Junction

{kind=link}
Abstract
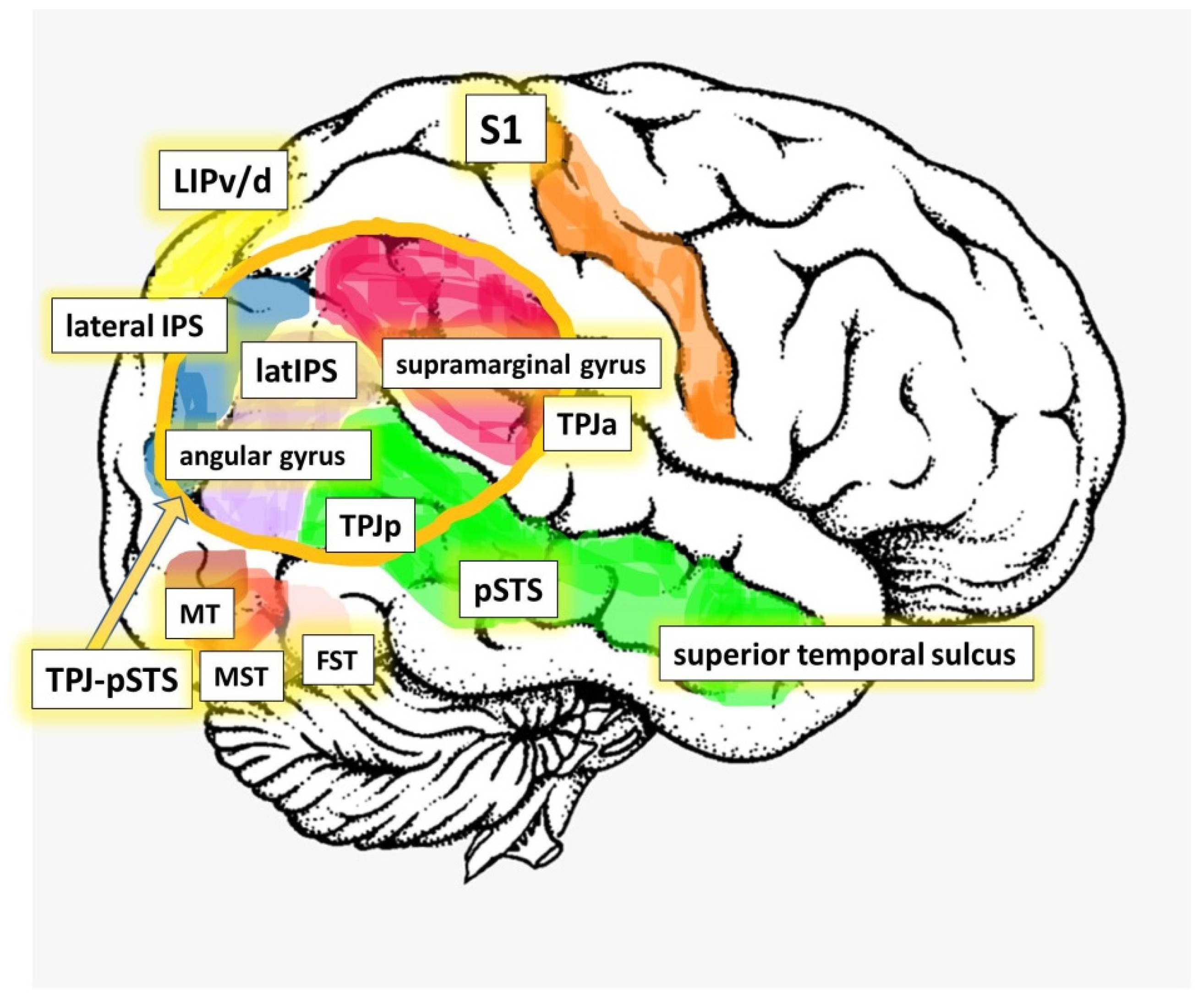
:1. The Right Temporal Parietal Junction
2. TMS and rTMS
3. tDCS
4. Patients and Brain Stimulation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mars, R.B.; Sallet, J.; Schüffelgen, U.; Jbabdi, S.; Toni, I.; Rushworth, M.F. Connectivity-based subdivisions of the human right “temporoparietal junction area”: Evidence for different areas participating in different cortical networks. Cereb. Cortex 2012, 22, 1894–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silani, G.; Lamm, C.; Ruff, C.C.; Singer, T. Right supramarginal gyrus is crucial to overcome emotional egocentricity bias in social judgments. J. Neurosci. 2013, 33, 15466–15476. [Google Scholar] [CrossRef] [PubMed]
- Morishima, Y.; Schunk, D.; Bruhin, A.; Ruff, C.C.; Fehr, E. Linking brain structure and activation in temporoparietal junction to explain the neurobiology of human altruism. Neuron 2012, 75, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strombach, T.; Weber, B.; Hangebrauk, Z.; Kenning, P.; Karipidis, I.I.; Tobler, P.N.; Kalenscher, T. Social discounting involves modulation of neural value signals by temporoparietal junction. Proc. Natl. Acad. Sci. USA 2015, 112, 1619–1624. [Google Scholar] [CrossRef] [Green Version]
- Saxe, R.; Kanwisher, N. People thinking about thinking people. The role of the temporo-parietal junction in theory of mind. Neuroimage 2003, 19, 1835–1842. [Google Scholar] [CrossRef]
- Chang, C.F.; Hsu, T.Y.; Tseng, P.; Liang, W.K.; Tzeng, O.J.; Hung, D.L.; Juan, C.H. Right temporoparietal junction and attentional reorienting. Hum. Brain Map. 2013, 34, 869–877. [Google Scholar] [CrossRef]
- Corbetta, M.; Patel, G.; Shulman, G.L. The reorienting system of the human brain: From environment to theory of mind. Neuron 2008, 58, 306–324. [Google Scholar] [CrossRef] [Green Version]
- Walsh, V.; Cowey, A. Transcranial magnetic stimulation and cognitive neuroscience. Nat. Rev. Neurosci. 2000, 1, 73–79. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Bartres-Faz, D.; Keenan, J.P. Transcranial magnetic stimulation: Studying the brain-behaviour relationship by induction of virtual lesions. Philos Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Maeda, F.; Keenan, J.P.; Tormos, J.M.; Topka, H.; Pascual-Leone, A. Modulation of corticospinal excitability by repetitive transcranial magnetic stimulation. Clin. Neurophysiol. 2000, 111, 800–805. [Google Scholar] [CrossRef]
- Barker, A.T.; Jalinous, R.; Freeston, I.L. Non-invasive magnetic stimulation of human motor cortex. Lancet 1985, 1, 1106–1107. [Google Scholar] [CrossRef]
- Bardi, L.; Six, P.; Brass, M. Repetitive TMS of the temporo-parietal junction disrupts participant’s expectations in a spontaneous Theory of Mind task. Soc. Cognit. Affect. Neurosci. 2017, 12, 1775–1782. [Google Scholar] [CrossRef] [Green Version]
- Maeda, F.; Keenan, J.P.; Pascual-Leone, A. Interhemispheric asymmetry of motor cortical excitability in major depression as measured by transcranial magnetic stimulation. Br. J. Psychiatry 2000, 177, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Maeda, F.; Keenan, J.P.; Tormos, J.M.; Topka, H.; Pascual-Leone, A. Interindividual variability of the modulatory effects of repetitive transcranial magnetic stimulation on cortical excitability. Exp. Brain Res. 2000, 133, 425–430. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Walsh, V.; Rothwell, J. Transcranial magnetic stimulation in cognitive neuroscience-virtual lesion, chronometry, and functional connectivity. Curr. Opin. Neurobiol. 2000, 10, 232–237. [Google Scholar] [CrossRef]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.J.; Fried, P.J.; Pascual-Leone, A. Transcranial magnetic stimulation: Neurophysiological and clinical applications. Handb. Clin. Neurol. 2019, 163, 73–92. [Google Scholar] [PubMed]
- Poreisz, C.; Boros, K.; Antal, A.; Paulus, W. Safety aspects of transcranial direct current stimulation concerning healthy subjects and patients. Brain Res. Bull. 2007, 72, 208–214. [Google Scholar] [CrossRef]
- Luber, B.; Fisher, C.; Appelbaum, P.S.; Ploesser, M.; Lisanby, S.H. Non-invasive brain stimulation in the detection of deception: Scientific challenges and ethical consequences. Behav. Sci. Law 2009, 27, 191–208. [Google Scholar] [CrossRef]
- McLaren, M.E.; Nissim, N.R.; Woods, A.J. The effects of medication use in transcranial direct current stimulation: A brief review. Brain Stimul. 2018, 11, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, Y.; Oizumi, R. Electric stimulation of the right temporo-parietal junction induces a task-specific effect in deceptive behaviors. Neurosci. Res. 2018, 128, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridgers, S.L. The safety of transcranial magnetic stimulation reconsidered: Evidence regarding cognitive and other cerebral effects. Electroencephalogr. Clin. Neurophysiol. Suppl. 1991, 43, 170–179. [Google Scholar] [PubMed]
- Gilbert, D.L.; Garvey, M.A.; Bansal, A.S.; Lipps, T.; Zhang, J.; Wassermann, E.M. Should transcranial magnetic stimulation research in children be considered minimal risk? Clin. Neurophysiol. 2004, 115, 1730–1739. [Google Scholar] [CrossRef]
- López-Ibor, J.J.; López-Ibor, M.I.; Pastrana, J.I. Transcranial magnetic stimulation. Curr. Opin. Psychiatry 2008, 21, 640–644. [Google Scholar] [CrossRef] [PubMed]
- San-Juan, D.; Dávila-Rodríguez, D.O.; Jiménez, C.R.; González, M.S.; Carranza, S.M.; Hernández Mendoza, J.R.; Anschel, D.J. Neuromodulation techniques for status epilepticus: A review. Brain Stimul. 2019, 12, 835–844. [Google Scholar] [CrossRef]
- Weissman-Fogel, I.; Granovsky, Y. The virtual lesion approach to transcranial magnetic stimulation: Studying the brain-behavioral relationships in experimental pain. Pain Rep. 2019, 4, e760. [Google Scholar] [CrossRef] [PubMed]
- Igelström, K.M.; Graziano, M.S.A. The inferior parietal lobule and temporoparietal junction: A network perspective. Neuropsychologia 2017, 105, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Schurz, M.; Tholen, M.G.; Perner, J.; Mars, R.B.; Sallet, J. Specifying the brain anatomy underlying temporo-parietal junction activations for theory of mind: A review using probabilistic atlases from different imaging modalities. Hum. Brain Map. 2017, 38, 4788–4805. [Google Scholar] [CrossRef]
- Patel, G.H.; Sestieri, C.; Corbetta, M. The evolution of the temporoparietal junction and posterior superior temporal sulcus. Cortex 2019, 118, 38–50. [Google Scholar] [CrossRef]
- Tsakiris, M.; Costantini, M.; Haggard, P. The role of the right temporo-parietal junction in maintaining a coherent sense of one’s body. Neuropsychologia 2008, 46, 3014–3018. [Google Scholar] [CrossRef]
- Jäncke, L.; Schlaug, G.; Huang, Y.; Steinmetz, H. Asymmetry of the planum parietale. Neuroreport 1994, 5, 1161–1163. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, H.; Herzog, A.; Schlaug, G.; Huang, Y.; Jäncke, L. Brain (A) symmetry in monozygotic twins. Cereb. Cortex 1995, 5, 296–300. [Google Scholar] [CrossRef]
- Hamilton, R.H.; Pascual-Leone, A.; Schlaug, G. Absolute pitch in blind musicians. Neuroreport 2004, 15, 803–806. [Google Scholar] [CrossRef]
- Keenan, J.P.; Thangaraj, V.; Halpern, A.R.; Schlaug, G. Absolute pitch and planum temporale. Neuroimage 2001, 14, 1402–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spocter, M.A.; Sherwood, C.C.; Schapiro, S.J.; Hopkins, W.D. Reproducibility of leftward planum temporale asymmetries in two genetically isolated populations of chimpanzees (Pan troglodytes). Proc. Biol. Sci. 2020, 287, 20201320. [Google Scholar] [CrossRef]
- Becker, Y.; Sein, J.; Velly, L.; Giacomino, L.; Renaud, L.; Lacoste, R.; Anton, J.L.; Nazarian, B.; Berne, C.; Meguerditchian, A. Early Left-Planum Temporale Asymmetry in newborn monkeys (Papio anubis): A longitudinal structural MRI study at two stages of development. Neuroimage 2021, 227, 117575. [Google Scholar] [CrossRef]
- Yuan, D.; Luo, D.; Kwok, V.P.Y.; Zhou, Y.; Tian, H.; Yu, Q.; An, J.; Gao, J.H.; Qiu, S.; Tan, L.H. Myeloarchitectonic Asymmetries of Language Regions in the Human Brain. Cereb. Cortex 2021, 31, 4169–4179. [Google Scholar] [CrossRef]
- Altarelli, I.; Leroy, F.; Monzalvo, K.; Fluss, J.; Billard, C.; Dehaene-Lambertz, G.; Galaburda, A.M.; Ramus, F. Planum temporale asymmetry in developmental dyslexia: Revisiting an old question. Hum. Brain Map. 2014, 35, 5717–5735. [Google Scholar] [CrossRef]
- Gough, P.M.; Connally, E.L.; Howell, P.; Ward, D.; Chesters, J.; Watkins, K.E. Planum temporale asymmetry in people who stutter. J. Fluency Dis. 2018, 55, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Vanderauwera, J.; Altarelli, I.; Vandermosten, M.; De Vos, A.; Wouters, J.; Ghesquière, P. Atypical Structural Asymmetry of the Planum Temporale is Related to Family History of Dyslexia. Cereb. Cortex 2018, 28, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Dronkers, N.F.; Wilkins, D.P.; Van Valin, R.D., Jr.; Redfern, B.B.; Jaeger, J.J. Lesion analysis of the brain areas involved in language comprehension. Cognition 2004, 92, 145–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boukrina, O.; Barrett, A.M. Disruption of the ascending arousal system and cortical attention networks in post-stroke delirium and spatial neglect. Neurosci. Biobehav. Rev. 2017, 83, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.M.; Boukrina, O.; Saleh, S. Ventral attention and motor network connectivity is relevant to functional impairment in spatial neglect after right brain stroke. Brain Cogn. 2019, 129, 16–24. [Google Scholar] [CrossRef]
- Fujii, N.; Hihara, S.; Iriki, A. Social cognition in premotor and parietal cortex. Soc. Neurosci. 2008, 3, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Hiser, J.; Koenigs, M. The Multifaceted Role of the Ventromedial Prefrontal Cortex in Emotion, Decision Making, Social Cognition, and Psychopathology. Biol. Psychiatry 2018, 83, 638–647. [Google Scholar] [CrossRef]
- Mukerji, C.E.; Lincoln, S.H.; Dodell-Feder, D.; Nelson, C.A.; Hooker, C.I. Neural correlates of theory-of-mind are associated with variation in children’s everyday social cognition. Soc. Cogn. Affect. Neurosci. 2019, 14, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Anticevic, A.; Repovs, G.; Shulman, G.L.; Barch, D.M. When less is more: TPJ and default network deactivation during encoding predicts working memory performance. Neuroimage 2010, 49, 2638–2648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Mai, X.; Liu, C. The default mode network and social understanding of others: What do brain connectivity studies tell us. Front. Hum. Neurosci. 2014, 8, 74. [Google Scholar] [CrossRef]
- Meyer, M.L.; Davachi, L.; Ochsner, K.N.; Lieberman, M.D. Evidence That Default Network Connectivity During Rest Consolidates Social Information. Cereb. Cortex 2019, 29, 1910–1920. [Google Scholar] [CrossRef] [Green Version]
- Lou, H.C. Self-awareness—An emerging field in neurobiology. Acta Paediatr. 2015, 104, 121–122. [Google Scholar] [CrossRef] [Green Version]
- Feinberg, T.E.; Keenan, J.P. Where in the brain is the self? Conscious. Cogn. 2005, 14, 661–678. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, Y.; Ohnishi, T.; Lane, R.D.; Maeda, M.; Mori, T.; Nemoto, K.; Matsuda, H.; Komaki, G. Impaired self-awareness and theory of mind: An fMRI study of mentalizing in alexithymia. Neuroimage 2006, 32, 1472–1482. [Google Scholar] [CrossRef]
- Chan, Y.C.; Lavallee, J.P. Temporo-parietal and fronto-parietal lobe contributions to theory of mind and executive control: An fMRI study of verbal jokes. Front. Psychol. 2015, 6, 1285. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, S.; van Schie, H.T.; Cross, E.S.; de Lange, F.P.; Wigboldus, D.H. Disentangling neural processes of egocentric and allocentric mental spatial transformations using whole-body photos of self and other. Neuroimage 2015, 116, 30–39. [Google Scholar] [CrossRef]
- Bulgarelli, C.; Blasi, A.; de Klerk, C.; Richards, J.E.; Hamilton, A.; Southgate, V. Fronto-temporoparietal connectivity and self-awareness in 18-month-olds: A resting state fNIRS study. Dev. Cogn. Neurosci. 2019, 38, 100676. [Google Scholar] [CrossRef]
- Salgado-Pineda, P.; Fuentes-Claramonte, P.; Spanlang, B.; Pomes, A.; Landin-Romero, R.; Portillo, F.; Bosque, C.; Franquelo, J.C.; Teixido, C.; Sarró, S.; et al. Neural correlates of disturbance in the sense of agency in schizophrenia: An fMRI study using the ’enfacement’ paradigm. Schizophr. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Salgues, S.; Plancher, G.; Jacquot, L.; Naveteur, J.; Fanuel, L.; Gálvez-García, G.; Michael, G.A. To the self and beyond: Arousal and functional connectivity of the temporo-parietal junction contributes to spontaneous sensations perception. Behav. Brain Res. 2021, 396, 112880. [Google Scholar] [CrossRef] [PubMed]
- Decety, J.; Keenan, J.P. Social Neuroscience: A new journal. Soc. Neurosci. 2006, 1, 1–4. [Google Scholar] [CrossRef]
- Grice-Jackson, T.; Critchley, H.D.; Banissy, M.J.; Ward, J. Common and distinct neural mechanisms associated with the conscious experience of vicarious pain. Cortex 2017, 94, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Saito, D.N.; Ban, M.; Shimada, K.; Okamoto, Y.; Kosaka, H.; Okazawa, H.; Asada, M.; Naito, E. Self-face recognition shares brain regions active during proprioceptive illusion in the right inferior fronto-parietal superior longitudinal fasciculus III network. Neuroscience 2017, 348, 288–301. [Google Scholar] [CrossRef] [PubMed]
- van Veluw, S.J.; Chance, S.A. Differentiating between self and others: An ALE meta-analysis of fMRI studies of self-recognition and theory of mind. Brain Imag. Behav. 2014, 8, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Jardri, R.; Pins, D.; Lafargue, G.; Very, E.; Ameller, A.; Delmaire, C.; Thomas, P. Increased overlap between the brain areas involved in self-other distinction in schizophrenia. PLoS ONE 2011, 6, e17500. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Kaplan, J.T.; Molnar-Szakacs, I.; Zaidel, E.; Iacoboni, M. Self-face recognition activates a frontoparietal "mirror" network in the right hemisphere: An event-related fMRI study. Neuroimage 2005, 25, 926–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, J.P.; Rubio, J.; Racioppi, C.; Johnson, A.; Barnacz, A. The right hemisphere and the dark side of consciousness. Cortex 2005, 41, 695–704. [Google Scholar] [CrossRef]
- Blanke, O.; Arzy, S. The out-of-body experience: Disturbed self-processing at the temporo-parietal junction. Neuroscientist 2005, 11, 16–24. [Google Scholar] [CrossRef]
- Blanke, O.; Landis, T.; Spinelli, L.; Seeck, M. Out-of-body experience and autoscopy of neurological origin. Brain 2004, 127, 243–258. [Google Scholar] [CrossRef] [Green Version]
- Blanke, O.; Mohr, C.; Michel, C.M.; Pascual-Leone, A.; Brugger, P.; Seeck, M.; Landis, T.; Thut, G. Linking out-of-body experience and self processing to mental own-body imagery at the temporoparietal junction. J. Neurosci. 2005, 25, 550–557. [Google Scholar] [CrossRef]
- Arzy, S.; Thut, G.; Mohr, C.; Michel, C.M.; Blanke, O. Neural basis of embodiment: Distinct contributions of temporoparietal junction and extrastriate body area. J. Neurosci. 2006, 26, 8074–8081. [Google Scholar] [CrossRef]
- Lenggenhager, B.; Smith, S.T.; Blanke, O. Functional and neural mechanisms of embodiment: Importance of the vestibular system and the temporal parietal junction. Rev. Neurosci. 2006, 17, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Easton, S.; Blanke, O.; Mohr, C. A putative implication for fronto-parietal connectivity in out-of-body experiences. Cortex 2009, 45, 216–227. [Google Scholar] [CrossRef]
- Ionta, S.; Gassert, R.; Blanke, O. Multi-sensory and sensorimotor foundation of bodily self-consciousness—An interdisciplinary approach. Front. Psychol. 2011, 2, 383. [Google Scholar] [CrossRef] [Green Version]
- Ionta, S.; Heydrich, L.; Lenggenhager, B.; Mouthon, M.; Fornari, E.; Chapuis, D.; Gassert, R.; Blanke, O. Multisensory mechanisms in temporo-parietal cortex support self-location and first-person perspective. Neuron 2011, 70, 363–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ionta, S.; Martuzzi, R.; Salomon, R.; Blanke, O. The brain network reflecting bodily self-consciousness: A functional connectivity study. Soc. Cogn. Affect. Neurosci. 2014, 9, 1904–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, C.; McDonald, J.J.; Theeuwes, J. Electrophysiological evidence of the capture of visual attention. J. Cogn. Neurosci. 2006, 18, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Lamy, D.; Leber, A.; Egeth, H.E. Effects of task relevance and stimulus-driven salience in feature-search mode. J. Exp. Psychol. Hum. Percept. Perform. 2004, 30, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Folk, C.L.; Remington, R.W.; Johnston, J.C. Involuntary covert orienting is contingent on attentional control settings. J. Exp. Psychol. Hum. Percept. Perform. 1992, 18, 1030–1044. [Google Scholar] [CrossRef]
- Krall, S.C.; Volz, L.J.; Oberwelland, E.; Grefkes, C.; Fink, G.R.; Konrad, K. The right temporoparietal junction in attention and social interaction: A transcranial magnetic stimulation study. Hum. Brain Map. 2016, 37, 796–807. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.G.; Xia, G.; Hastings, P.D. Right Temporoparietal Junction Involvement in Autonomic Responses to the Suffering of Others: A Preliminary Transcranial Magnetic Stimulation Study. Front. Hum. Neurosci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Chen, C.; Decety, J. How Situational Context Impacts Empathic Responses and Brain Activation Patterns. Front. Behav. Neurosci. 2017, 11, 165. [Google Scholar] [CrossRef] [Green Version]
- Decety, J.; Grèzes, J. The power of simulation: Imagining one’s own and other’s behavior. Brain Res. 2006, 1079, 4–14. [Google Scholar] [CrossRef]
- Yoder, K.J.; Decety, J. The Good, the bad, and the just: Justice sensitivity predicts neural response during moral evaluation of actions performed by others. J. Neurosci. 2014, 34, 4161–4166. [Google Scholar] [CrossRef] [Green Version]
- Yoder, K.J.; Lahey, B.B.; Decety, J. Callous traits in children with and without conduct problems predict reduced connectivity when viewing harm to others. Sci. Rep. 2016, 6, 20216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, L.; Camprodon, J.A.; Hauser, M.; Pascual-Leone, A.; Saxe, R. Disruption of the right temporoparietal junction with transcranial magnetic stimulation reduces the role of beliefs in moral judgments. Proc. Natl. Acad. Sci. USA 2010, 107, 6753–6758. [Google Scholar] [CrossRef] [Green Version]
- Chou, Y.; Chen, T.Y. Disruption on right temporoparietal junction with transcranial magnetic stimulation affects moral judgment: No difference between first- and third-personal narration with TMS. Neuropsychologia 2021, 157, 107858. [Google Scholar] [CrossRef] [PubMed]
- Zheltyakova, M.; Korotkov, A.; Cherednichenko, D.; Kireev, M. Functional interactions between neural substrates of socio-cognitive mechanisms involved in simple deception and manipulative truth. Brain Connect. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Pereira, A.M.; Gao, X.; Campos, B.M.; Derrington, E.; Corgnet, B.; Zhou, X.; Cendes, F.; Dreher, J.C. Right Temporoparietal Junction Underlies Avoidance of Moral Transgression in Autism Spectrum Disorder. J. Neurosci. 2021, 41, 1699–1715. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Geng, F.; Riggins, T.; Chen, G.; Redcay, E. Neural correlates of developing theory of mind competence in early childhood. Neuroimage 2019, 184, 707–716. [Google Scholar] [CrossRef]
- Bitsch, F.; Berger, P.; Nagels, A.; Falkenberg, I.; Straube, B. Impaired Right Temporoparietal Junction-Hippocampus Connectivity in Schizophrenia and Its Relevance for Generating Representations of Other Minds. Schizophr. Bull. 2019, 45, 934–945. [Google Scholar] [CrossRef]
- Mossad, S.I.; AuCoin-Power, M.; Urbain, C.; Smith, M.L.; Pang, E.W.; Taylor, M.J. Thinking about the thoughts of others; temporal and spatial neural activation during false belief reasoning. Neuroimage 2016, 134, 320–327. [Google Scholar] [CrossRef]
- Lombardo, M.V.; Chakrabarti, B.; Bullmore, E.T.; Baron-Cohen, S. Specialization of right temporo-parietal junction for mentalizing and its relation to social impairments in autism. Neuroimage 2011, 56, 1832–1838. [Google Scholar] [CrossRef]
- Dodell-Feder, D.; Koster-Hale, J.; Bedny, M.; Saxe, R. fMRI item analysis in a theory of mind task. Neuroimage 2011, 55, 705–712. [Google Scholar] [CrossRef]
- Fletcher, P.C.; Happé, F.; Frith, U.; Baker, S.C.; Dolan, R.J.; Frackowiak, R.S.; Frith, C.D. Other minds in the brain: A functional imaging study of “theory of mind” in story comprehension. Cognition 1995, 57, 109–128. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, H.L.; Frith, C.D. Functional imaging of theory of mind. Trends Cogn. Sci. 2003, 7, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Bitsch, F.; Berger, P.; Nagels, A.; Falkenberg, I.; Straube, B. The role of the right temporo-parietal junction in social decision-making. Hum. Brain Map. 2018, 39, 3072–3085. [Google Scholar] [CrossRef] [Green Version]
- Bardi, L.; Desmet, C.; Nijhof, A.; Wiersema, J.R.; Brass, M. Brain activation for spontaneous and explicit false belief tasks overlaps: New fMRI evidence on belief processing and violation of expectation. Soc. Cogn. Affect. Neurosci. 2016, 12, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Hyde, D.C.; Aparicio Betancourt, M.; Simon, C.E. Human temporal-parietal junction spontaneously tracks other’s beliefs: A functional near-infrared spectroscopy study. Hum. Brain Map. 2015, 36, 4831–4846. [Google Scholar] [CrossRef] [PubMed]
- Zeugin, D.; Notter, M.P.; Knebel, J.F.; Ionta, S. Temporo-parietal contribution to the mental representations of self/other face. Brain Cogn. 2020, 143, 105600. [Google Scholar] [CrossRef] [PubMed]
- Duffy, K.A.; Luber, B.; Adcock, R.A.; Chartrand, T.L. Enhancing activation in the right temporoparietal junction using theta-burst stimulation: Disambiguating between two hypotheses of top-down control of behavioral mimicry. PLoS ONE 2019, 14, e0211279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krall, S.C.; Rottschy, C.; Oberwelland, E.; Bzdok, D.; Fox, P.T.; Eickhoff, S.B.; Fink, G.R.; Konrad, K. The role of the right temporoparietal junction in attention and social interaction as revealed by ALE meta-analysis. Brain Struct. Funct. 2015, 220, 587–604. [Google Scholar] [CrossRef]
- Payne, S.; Tsakiris, M. Anodal transcranial direct current stimulation of right temporoparietal area inhibits self-recognition. Cogn. Affect. Behav. Neurosci. 2017, 17, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Santiesteban, I.; Banissy, M.J.; Catmur, C.; Bird, G. Enhancing social ability by stimulating right temporoparietal junction. Curr. Biol. 2012, 22, 2274–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.; Kuang, B.; Hu, P. Right Temporoparietal Junction Modulates In-Group Bias in Facial Emotional Mimicry: A tDCS Study. Front. Behav. Neurosci. 2020, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Chen, S.; Huang, D.; Zheng, H.; Jia, Y.; Luo, J. Modulation of Neural Activity in the Temporoparietal Junction with Transcranial Direct Current Stimulation Changes the Role of Beliefs in Moral Judgment. Front. Human Neurosci. 2015, 9, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogeveen, J.; Obhi, S.S.; Banissy, M.J.; Santiesteban, I.; Press, C.; Catmur, C.; Bird, G. Task-dependent and distinct roles of the temporoparietal junction and inferior frontal cortex in the control of imitation. Soc. Cogn. Affect. Neurosci. 2015, 10, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- Nobusako, S.; Nishi, Y.; Nishi, Y.; Shuto, T.; Asano, D.; Osumi, M.; Morioka, S. Transcranial Direct Current Stimulation of the Temporoparietal Junction and Inferior Frontal Cortex Improves Imitation-Inhibition and Perspective-Taking with no Effect on the Autism-Spectrum Quotient Score. Front. Behav. Neurosci. 2017. [Google Scholar] [CrossRef]
- Wible, C.G. Hippocampal temporal-parietal junction interaction in the production of psychotic symptoms: A framework for understanding the schizophrenic syndrome. Front. Hum. Neurosci. 2012, 6, 180. [Google Scholar] [CrossRef] [Green Version]
- Abu-Akel, A.M.; Apperly, I.A.; Wood, S.J.; Hansen, P.C. Autism and psychosis expressions diametrically modulate the right temporoparietal junction. Soc. Neurosci. 2017, 12, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, D.A.; Xia, P.; McNally Keehn, R.; Dydak, U.; Keehn, B. A Magnetic Resonance Spectroscopy Study of Superior Visual Search Abilities in Children with Autism Spectrum Disorder. Autism Res. 2020, 13, 550–562. [Google Scholar] [CrossRef]
- Kana, R.K.; Libero, L.E.; Hu, C.P.; Deshpande, H.D.; Colburn, J.S. Functional brain networks and white matter underlying theory-of-mind in autism. Soc. Cogn. Affect. Neurosci. 2014, 9, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Nijhof, A.D.; Bardi, L.; Brass, M.; Wiersema, J.R. Brain activity for spontaneous and explicit mentalizing in adults with autism spectrum disorder: An fMRI study. Neuroimag. Clin. 2018, 18, 475–484. [Google Scholar] [CrossRef]
- Nijhof, A.D.; Dhar, M.; Goris, J.; Brass, M.; Wiersema, J.R. Atypical neural responding to hearing one’s own name in adults with ASD. J. Abnorm. Psychol. 2018, 127, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, R.E.; Boutros, N.N.; Berman, R.M.; Roessler, E.; Belger, A.; Krystal, J.H.; Charney, D.S. Transcranial magnetic stimulation of left temporoparietal cortex in three patients reporting hallucinated voices. Biol. Psychiatry 1999, 46, 130–132. [Google Scholar] [CrossRef]
- McIntosh, A.M.; Semple, D.; Tasker, K.; Harrison, L.K.; Owens, D.G.; Johnstone, E.C.; Ebmeier, K.P. Transcranial magnetic stimulation for auditory hallucinations in schizophrenia. Psychiatry Res. 2004, 127, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, W.; Chung, Y.C.; Jung, K.H.; Bahk, W.M.; Jun, T.Y.; Kim, K.S.; George, M.S.; Chae, J.H. A double blind study showing that two weeks of daily repetitive TMS over the left or right temporoparietal cortex reduces symptoms in patients with schizophrenia who are having treatment-refractory auditory hallucinations. Neurosci. Lett. 2005, 376, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Gromann, P.M.; Tracy, D.K.; Giampietro, V.; Brammer, M.J.; Krabbendam, L.; Shergill, S.S. Examining frontotemporal connectivity and rTMS in healthy controls: Implications for auditory hallucinations in schizophrenia. Neuropsychology 2012, 26, 127–132. [Google Scholar] [CrossRef]
- Kindler, J.; Homan, P.; Jann, K.; Federspiel, A.; Flury, R.; Hauf, M.; Strik, W.; Dierks, T.; Hubl, D. Reduced neuronal activity in language-related regions after transcranial magnetic stimulation therapy for auditory verbal hallucinations. Biol. Psychiatry 2013, 73, 518–524. [Google Scholar] [CrossRef]
- Chauhan, P.; Garg, S.; Tikka, S.K.; Khattri, S. Efficacy of Intensive Cerebellar Intermittent Theta Burst Stimulation (iCiTBS) in Treatment-Resistant Schizophrenia: A Randomized Placebo-Controlled Study. Cerebellum 2021, 20, 116–123. [Google Scholar] [CrossRef]
- Wobrock, T.; Guse, B.; Cordes, J.; Wölwer, W.; Winterer, G.; Gaebel, W.; Langguth, B.; Landgrebe, M.; Eichhammer, P.; Frank, E.; et al. Left prefrontal high-frequency repetitive transcranial magnetic stimulation for the treatment of schizophrenia with predominant negative symptoms: A sham-controlled, randomized multicenter trial. Biol. Psychiatry 2015, 77, 979–988. [Google Scholar] [CrossRef]
- Julkunen, P.; Jauhiainen, A.M.; Westerén-Punnonen, S.; Pirinen, E.; Soininen, H.; Könönen, M.; Pääkkönen, A.; Määttä, S.; Karhu, J. Navigated TMS combined with EEG in mild cognitive impairment and Alzheimer’s disease: A pilot study. J. Neurosci. Methods 2008, 172, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Salehinejad, M.A.; Paknia, N.; Hosseinpour, A.H.; Yavari, F.; Vicario, C.M.; Nitsche, M.A.; Nejati, V. Contribution of the right temporoparietal junction and ventromedial prefrontal cortex to theory of mind in autism: A randomized, sham-controlled tDCS study. Autism Res. 2021, 14, 1572–1584. [Google Scholar] [CrossRef]
- Donaldson, P.H.; Kirkovski, M.; Rinehart, N.J.; Enticott, P.G. Autism-relevant traits interact with temporoparietal junction stimulation effects on social cognition: A high-definition transcranial direct current stimulation and electroencephalography study. Eur. J. Neurosci. 2018, 47, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Luckhardt, C.; Schütz, M.; Mühlherr, A.; Mössinger, H.; Boxhoorn, S.; Dempfle, A.; Salvador, R.; Ruffini, G.; Pereira, H.C.; Castelo-Branco, M.; et al. Phase-IIa randomized, double-blind, sham-controlled, parallel group trial on anodal transcranial direct current stimulation (tDCS) over the left and right tempo-parietal junction in autism spectrum disorder-StimAT: Study protocol for a clinical trial. Trials 2021, 22, 248. [Google Scholar] [CrossRef] [PubMed]
- Esse Wilson, J.; Quinn, D.K.; Wilson, J.K.; Garcia, C.M.; Tesche, C.D. Transcranial Direct Current Stimulation to the Right Temporoparietal Junction for Social Functioning in Autism Spectrum Disorder: A Case Report. J. Ect. 2018, 34, e10–e13. [Google Scholar] [CrossRef] [PubMed]
- Esse Wilson, J.; Trumbo, M.C.; Wilson, J.K.; Tesche, C.D. Transcranial direct current stimulation (tDCS) over right temporoparietal junction (rTPJ) for social cognition and social skills in adults with autism spectrum disorder (ASD). J. Neural. Transm. 2018, 125, 1857–1866. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, N.; Zorns, S.; Chavarria, K.; Brenya, J.; Janowska, A.; Keenan, J.P. Are We Right about the Right TPJ? A Review of Brain Stimulation and Social Cognition in the Right Temporal Parietal Junction. Symmetry 2021, 13, 2219. https://doi.org/10.3390/sym13112219
Ahmad N, Zorns S, Chavarria K, Brenya J, Janowska A, Keenan JP. Are We Right about the Right TPJ? A Review of Brain Stimulation and Social Cognition in the Right Temporal Parietal Junction. Symmetry. 2021; 13(11):2219. https://doi.org/10.3390/sym13112219
Chicago/Turabian StyleAhmad, Nathira, Samantha Zorns, Katherine Chavarria, Janet Brenya, Aleksandra Janowska, and Julian Paul Keenan. 2021. "Are We Right about the Right TPJ? A Review of Brain Stimulation and Social Cognition in the Right Temporal Parietal Junction" Symmetry 13, no. 11: 2219. https://doi.org/10.3390/sym13112219
APA StyleAhmad, N., Zorns, S., Chavarria, K., Brenya, J., Janowska, A., & Keenan, J. P. (2021). Are We Right about the Right TPJ? A Review of Brain Stimulation and Social Cognition in the Right Temporal Parietal Junction. Symmetry, 13(11), 2219. https://doi.org/10.3390/sym13112219