Symmetry Transformations in Metazoan Evolution and Development

Abstract
:1. Introduction
2. Phylogenomic Data: The Genetic Background of Axial Patterning
3. Primary Body Axis: The Wnt Signaling Pathway
4. Symmetry in Bilateria
4.1. Axial Hox Code
4.2. Dorsal–Ventral Axis: The BMP Signaling Pathway
4.3. Metamerism in Bilateria: Wnt and Notch Signaling Pathways
4.4. Posterior Growth Mechanism in Bilateria
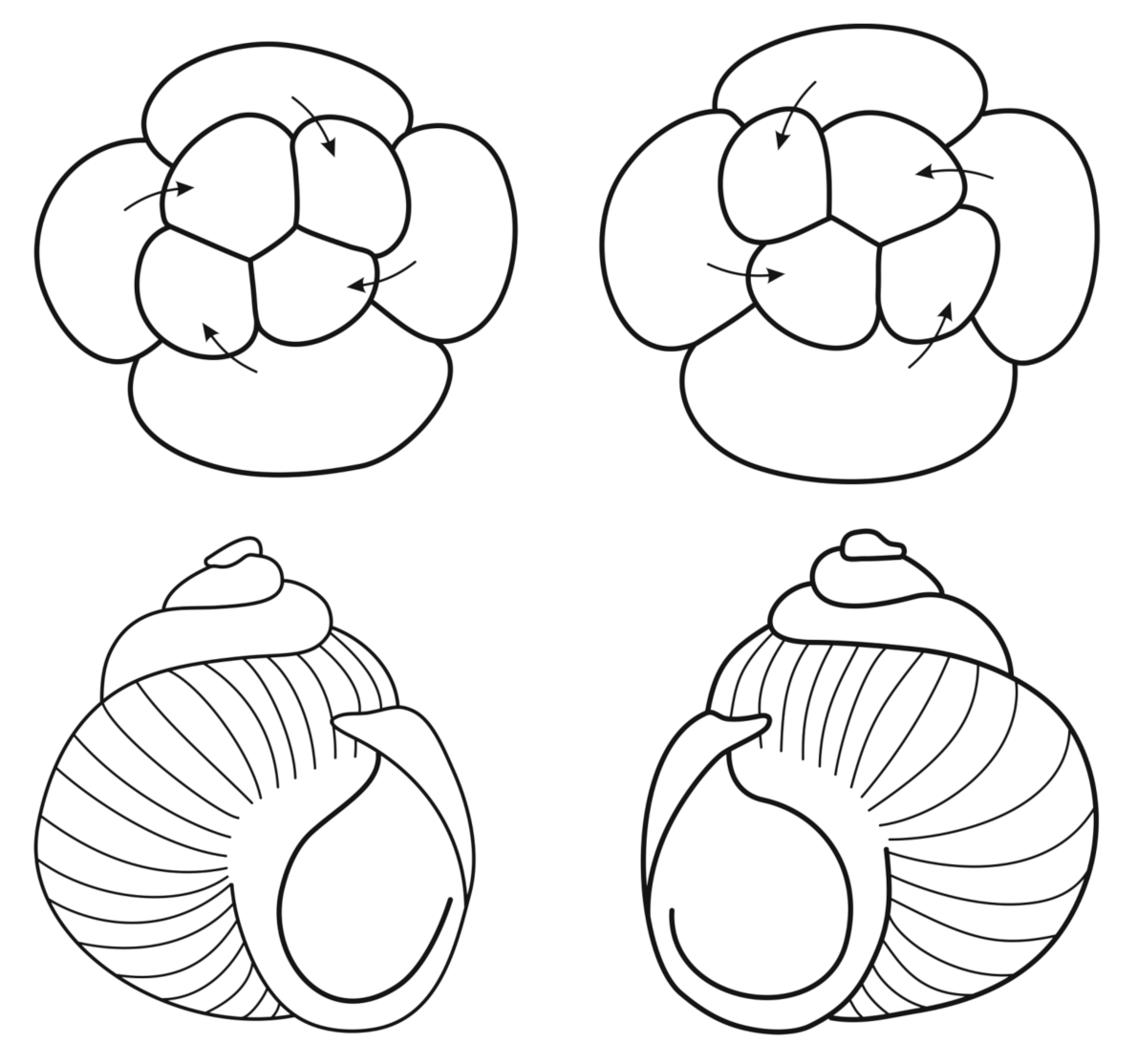
4.5. Left–Right Symmetry/Asymmetry in Bilateria: Nodal and Hedgehog Signaling Pathways
5. Helicoidal Symmetry: Nodal Pathway
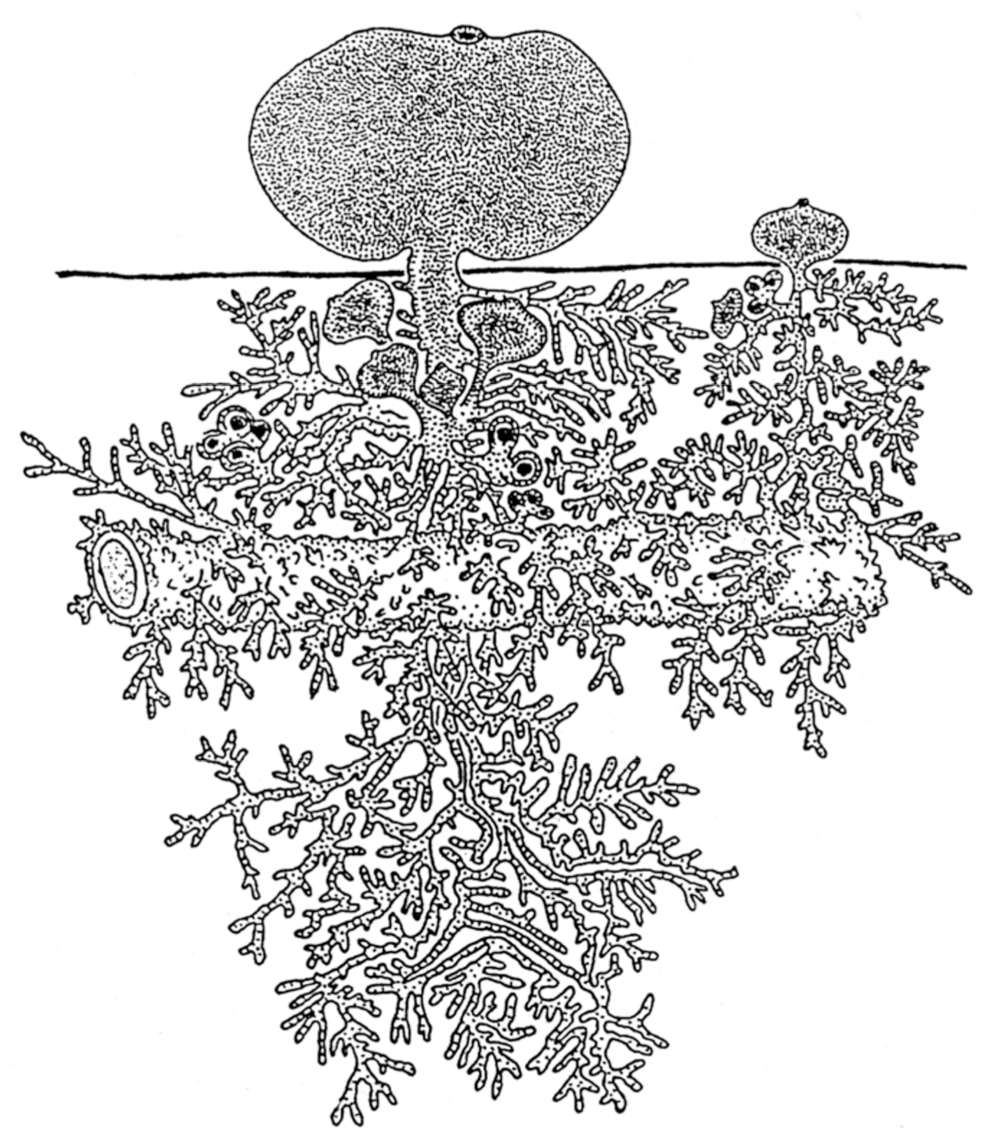
6. Scale Symmetry
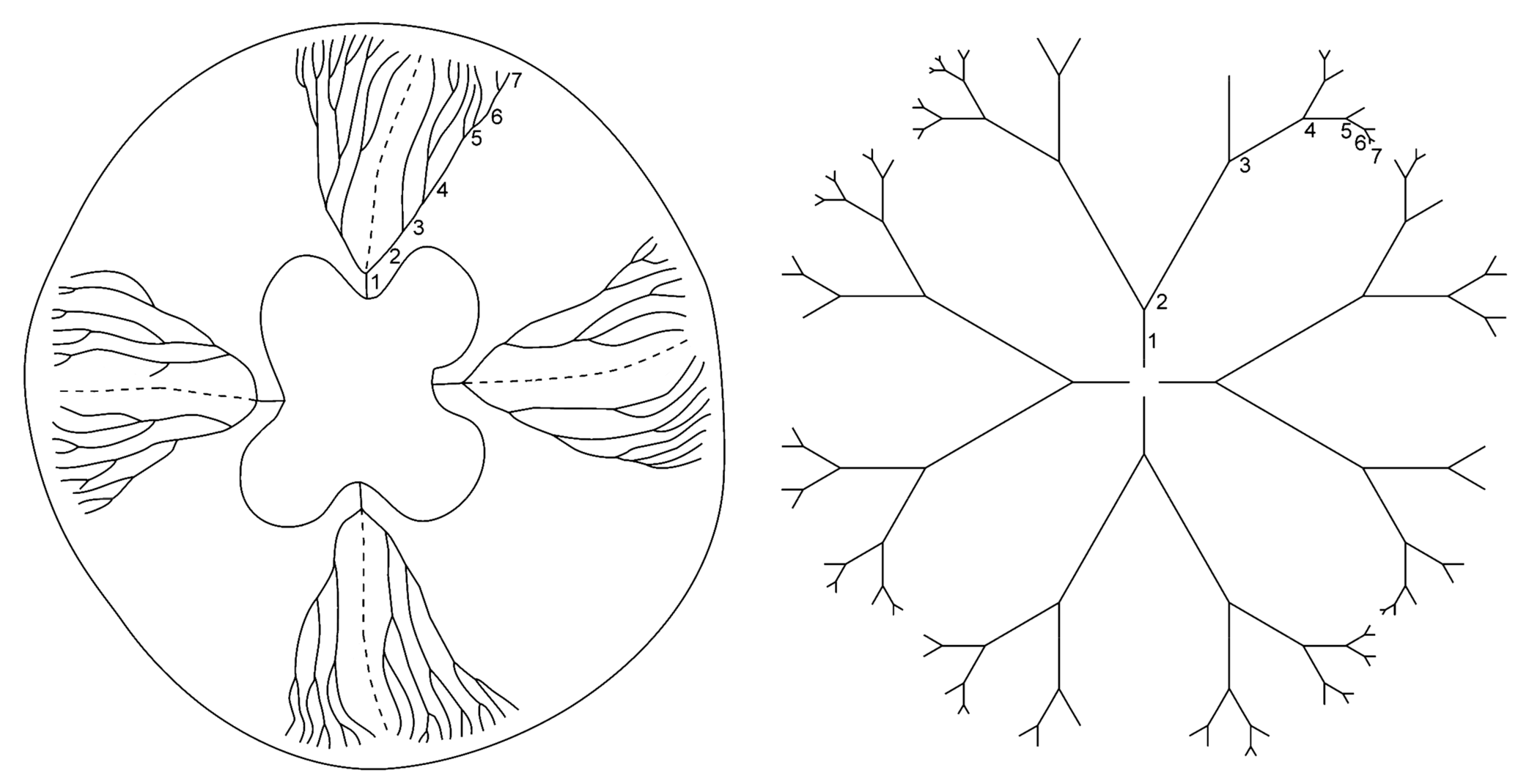
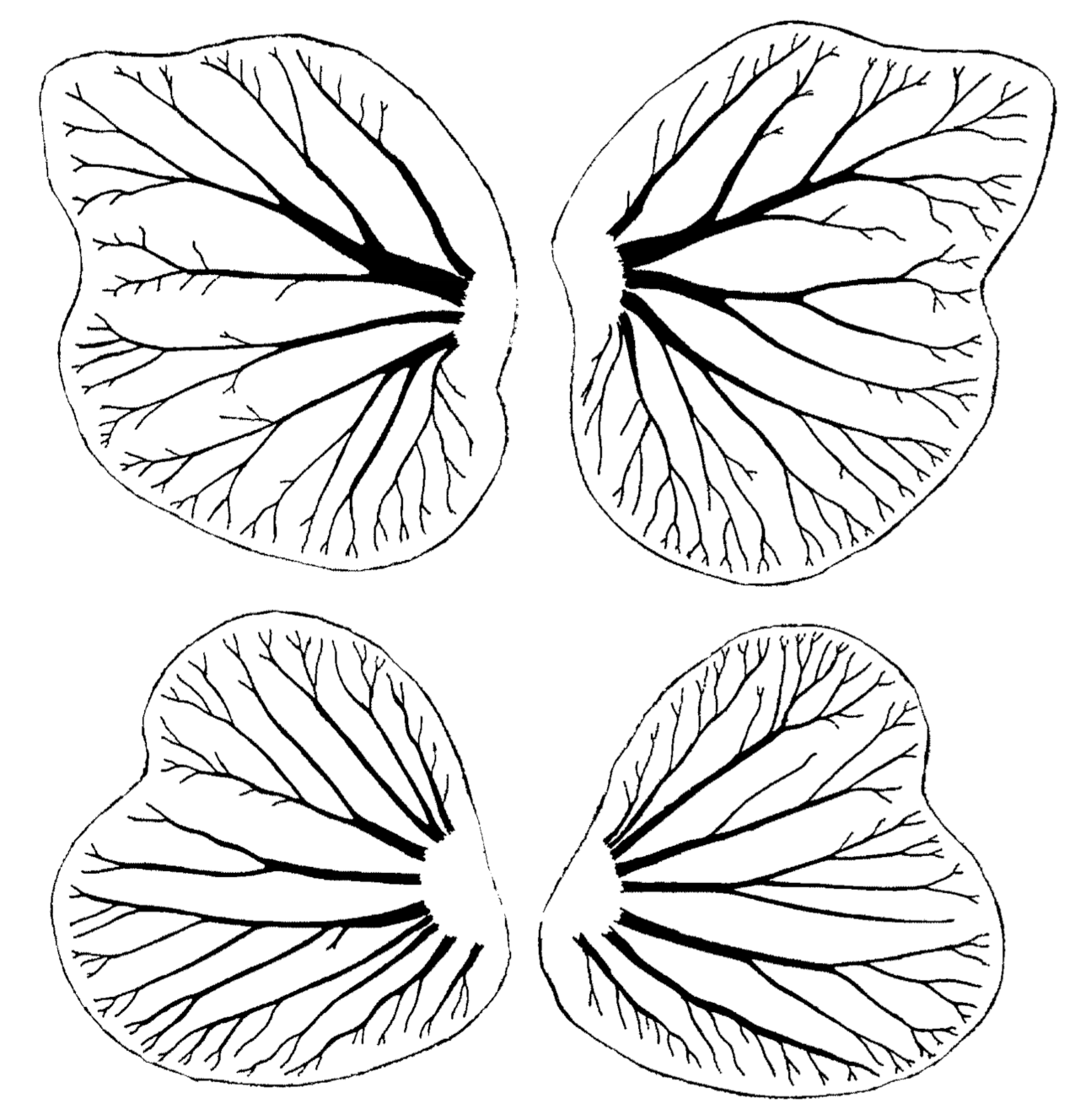
6.1. Fluctuating Asymmetry in Biological Fractal Structures
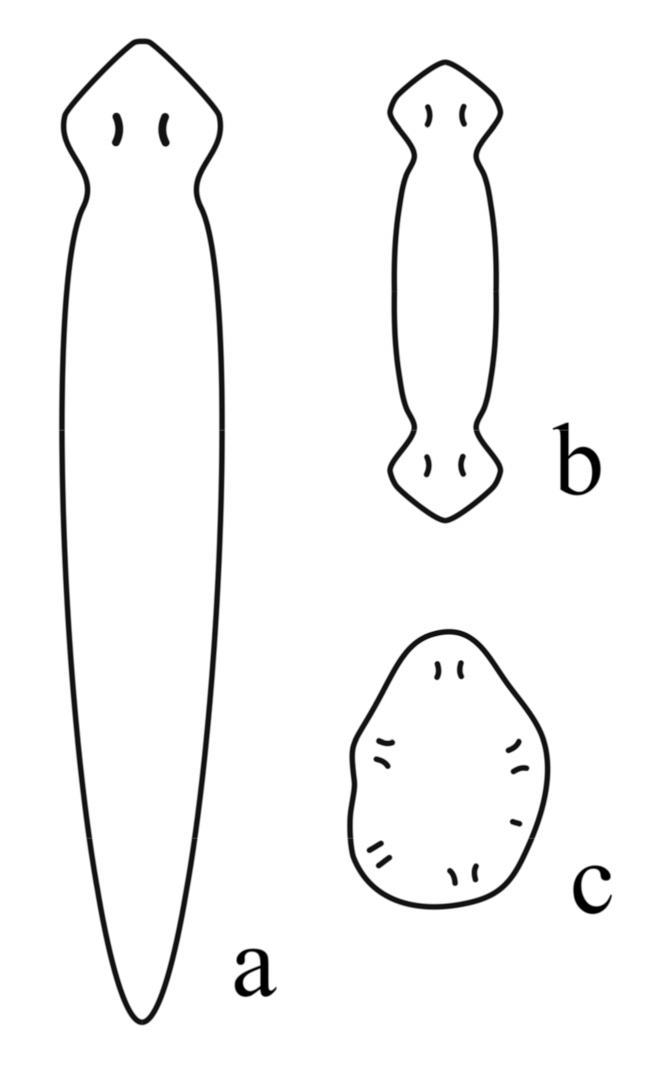
7. Body Plan and Symmetry of Ediacaran (Vendian) Metazoans
7.1. Lost Ediacaran Body Plans
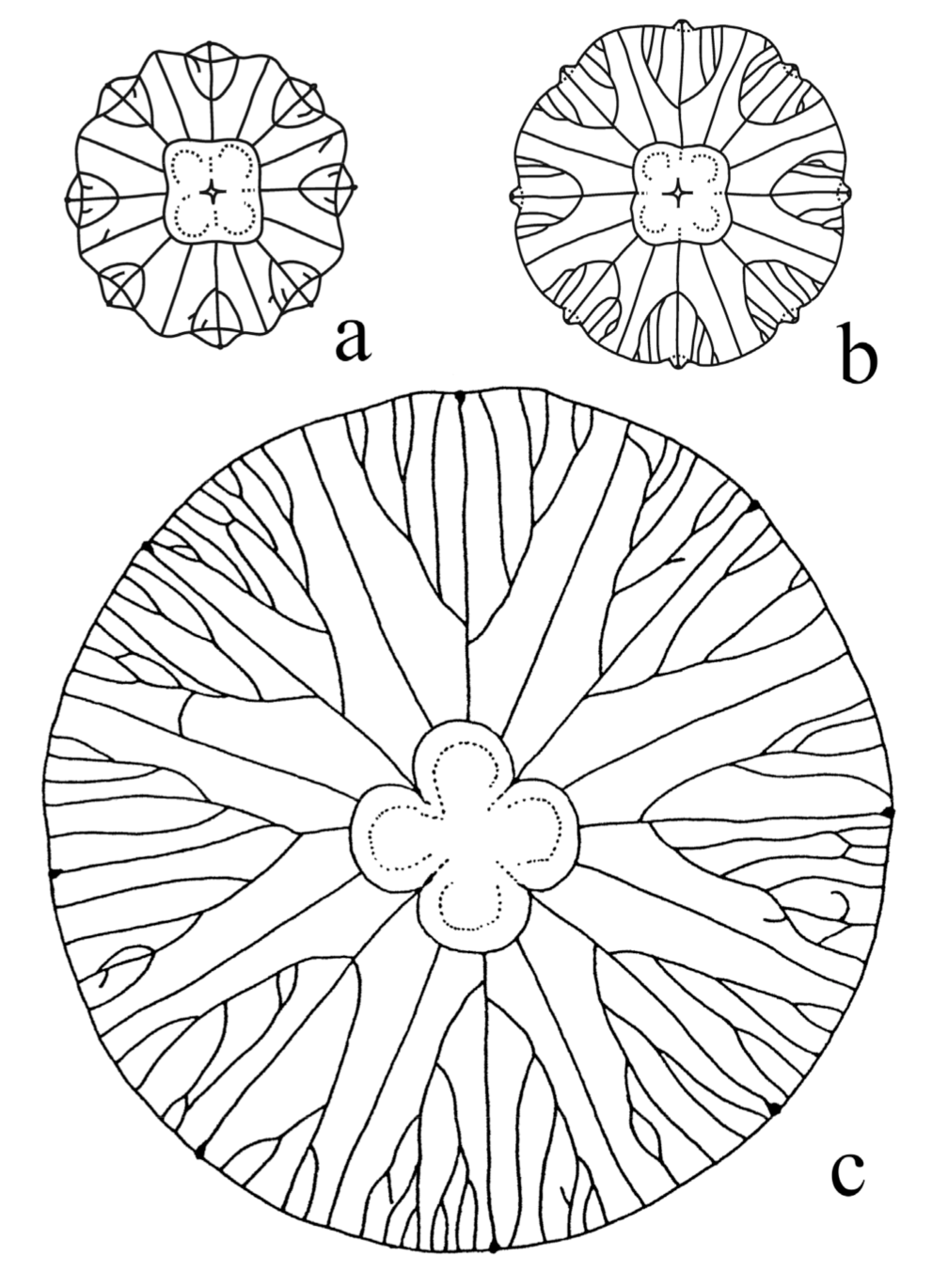
7.2. Early Cambrian Helicoidal-Bodied Echinoderms
7.3. Bilateral-to-Pentameral Transition in Echinodermata
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beklemishev, V.N. Principles of Comparative Anatomy of Invertebrates; Beklemishev, V.N., Ed.; Oliver & Boyd: Edinburgh, UK, 1969; Nauka: Moscow, Russia, 1964. (In Russian) [Google Scholar]
- Gerhart, J.C.; Kirschner, M.W. Cells, Embryos, and Evolution; Blackwell Science: Boston, MA, USA, 1997. [Google Scholar]
- Minelli, A. The Development of Animal Form. Ontogeny, Morphology, and Evolution; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Minelli, A. EvoDevo and its significance for animal evolution and phylogeny. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; Volume 1, pp. 1–24. [Google Scholar]
- Ryan, J.F.; Baxevanis, A.D. Hox, Wnt, and the evolution of the primary body axis: Insights from the early-divergent phyla. Biol. Direct. 2007, 2, 37. [Google Scholar] [CrossRef] [Green Version]
- De Robertis, E.M. Evo-devo: Variations on ancestral themes. Cell 2008, 132, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Meinhardt, H. The primary body axes of vertebrates: The generation of a near-Cartesian coordinate system and the role of Spemann-type organizer. Dev. Dyn. 2006, 235, 2907–2919. [Google Scholar] [CrossRef]
- Manuel, M. Early evolution of symmetry and polarity in metazoan body plans. Comptes Rendus Biol. 2009, 332, 184–209. [Google Scholar]
- Niehrs, C. On growth and form: A Cartesian coordinate system of Wnt and BMP signaling specifies bilaterian body axes. Development 2010, 137, 845–857. [Google Scholar] [CrossRef] [Green Version]
- Holló, G. A new paradigm for animal symmetry. Interface Focus 2015, 5, 20150032. [Google Scholar] [CrossRef] [Green Version]
- Seilacher, A.; Gishlick, A.D.S. Morphodynamics; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2015. [Google Scholar]
- Erwin, D.H. The origin of animal body plans: A view from fossil evidence and the regulatory genome. Development 2020, 147, dev182899. [Google Scholar] [CrossRef] [PubMed]
- Fields, C.; Levin, M. Does regeneration recapitulate phylogeny? Planaria as a model of body-axis specification in ancestral eumetazoa. Comm. Integr. Biol. 2020, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar]
- Mandelbrot, B.B. The Fractal Geometry of Nature; Freeman: New York, NY, USA, 1983. [Google Scholar]
- Stewart, I. Why Beauty Is Truth: The Story of Symmetry; Perseus: New York, NY, USA, 2007. [Google Scholar]
- Isaeva, V.V.; Kasyanov, N.V.; Presnov, E.V. Analysis situs of spatial-emporal architecture in biological morphogenesis. In Progress in Mathematical Biology Research; Kelly, J.T., Ed.; Nova Science Publishers: New York, NY, USA, 2008; pp. 141–189. [Google Scholar]
- Isaeva, V.V.; Kasyanov, N.V.; Presnov, E.V. Topological singularities and symmetry breaking in development. BioSystems 2012, 109, 280–298. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating asymmetry: Methods, theory, and applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef] [Green Version]
- Beloussov, L.V. Self-organization, symmetry and morphomechanics in development of organisms. In Embryology–Updates and Highlights on Classic Topics; Pereira, L.A.V., Ed.; InTech: Rijeka, Croatia, 2012; pp. 189–210. [Google Scholar]
- Isaeva, V.V. Topological and fractal approaches to systematics and biodiversity. In Invertebrates: Classification, Evolution and Biodiversity; Riosmena-Rodrigez, R., Ed.; Nova Science Publishers: New York, NY, USA, 2013; pp. 225–242. [Google Scholar]
- Isaeva, V.V. Symmetry transformations in ontogeny and evolution. Paleontol. J. 2014, 48, 1127–1136. [Google Scholar] [CrossRef]
- Rozhnov, S.V. Symmetry of echinoderms: From initial bilaterally-asymmetric metamerism to pentaradiality. Paleontol. J. 2014, 6, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Stern, C.D.; Charité, J.; Deschamps, J.; Duboule, D.; Durston, A.J.; Kmita, M.; Nicolas, J.-F.; Palmeirim, I.; Smith, J.C.; Wolpert, L. Head-tail patterning of the vertebrate embryo: One, two or many unresolved problems? Int. J. Dev. Biol. 2006, 50, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Corballis, M.C. Bilaterally symmetrical: To be or not to be? Symmetry 2020, 12, 326. [Google Scholar] [CrossRef] [Green Version]
- Hirokawa, N.; Tanaka, Y.; Okada, Y. Left–right determination: Involvement of molecular motor KIF3, cilia, and nodal flow. Cold. Spring Harb. Perspect. Biol. 2009, 1, a000802. [Google Scholar] [CrossRef] [Green Version]
- Bouligand, Y. Morphological singulatities and macroevolution. Memor. Soc. Ital. Sci. Natur. Museo Civico Storia Natur. Milano 1996, 27, 89–94. [Google Scholar]
- Li, R.; Bowerman, B. Symmetry breaking in biology. Cold Spring Harb. Perspect. Biol. 2010, 2, a003475.
- Thom, R. Topological models in biology. Topology 1969, 8, 313–335. [Google Scholar] [CrossRef] [Green Version]
- Thom, R. Structural Stability and Morphogenesis: An Outline of a General Theory of Models; Addison-Wesley Publishing Co: New York, NY, USA, 1997. [Google Scholar]
- Bérczi, S.; Lukács, B. Symmetry versus katachi in animal organisms. Forma 2001, 16, 91–99. [Google Scholar]
- Larroux, C.; Fahey, B.; Degnan, S.M.; Adamski, M.; Rokhsar, D.S.; Degnan, B.M. The NK homeobox gene cluster predates the origin of Hox genes. Curr. Biol. 2007, 17, 706–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.F. Developmental Biology, 10th ed.; Sinauer Ass. Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Holstein, T.W. The evolution of the Wnt pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a007922. [Google Scholar] [CrossRef] [PubMed]
- Simakov, O.; Kawashima, T. Independent evolution of genomic characters during major metazoan transitions. Dev. Biol. 2017, 427, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, M. A Comparative genomics perspective on the origin of multicellularity and early animal evolution. In Evolutionary Transitions to Multicellular Life. Principles and Mechanisms; Ruiz-Trillo, I., Nedelcu, A.M., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2015; pp. 269–300. [Google Scholar]
- Dunn, F.S.; Liu, A.G.; Donoghue, P.C.J. Ediacaran developmental biology. Biol. Rev. 2017, 1. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.-H.; Chen, Y.-C.; Ting, H.-C.; Fan, T.-P.; Lin, C.-Y.; Wang, K.-T.; Yu, J.-K. BMP controls dorsoventral and neural patterning in indirect-developing hemichordates providing insight into a possible origin of chordates. Proc. Natl. Acad. Sci. USA 2019, 116, 12925–12932. [Google Scholar] [CrossRef] [Green Version]
- Danilchik, M.V.; Brown, E.E.; Riegert, K. Intrinsic chiral properties of the Xenopus egg cortex: An early indicator of left–right asymmetry? Development 2006, 133, 4517–4526. [Google Scholar] [CrossRef] [Green Version]
- Davidson, E.H. The Regulatory Genome: Gene Regulatory Networks in Development and Evolution; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- Rogers, K.W.; Schier, A.F. Morphogen gradients: From generation to interpretation. Ann. Rev. Cell Dev. Biol. 2011, 27, 377–407. [Google Scholar] [CrossRef] [Green Version]
- Bajard, L.; Morelli, L.G.; Ares, S.; Pécréaux, J.; Jülicher, F.; Oates, A.C. Wnt-regulated dynamics of positional information in zebrafish somitogenesis. Development 2014, 141, 1381–1391. [Google Scholar] [CrossRef] [Green Version]
- Lyons, D.C.; Martindale, M.Q.; Srivastava, M. The cell’s view of animal body-plan evolution. Integr. Comp. Biol. 2014, 54, 658–666. [Google Scholar] [CrossRef] [Green Version]
- Dunn, C.W.; Ryan, J.F. The evolution of animal genomes. Curr. Opin. Genet. Dev. 2015, 54, 25–32. [Google Scholar] [CrossRef]
- Isaeva, V.V. Genomic-morphogenetic correlations and evolutionary trajectories of Bilateria. Paleontol. J. 2018, 52, 1655–1662. [Google Scholar] [CrossRef]
- Isaeva, V.V.; Rozhnov, S.V. Evolutionary transformations of metazoan body plan: Genomic-morphogenetic correlations. Paleontol. J. 2021, 55, in press. [Google Scholar]
- Erwin, D.H. Early origin of the bilaterian developmental toolkit. Phil. Trans. R. Soc. 2009, 364, 2253–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erwin, D.H.; Valentine, J.W. The Cambrian Explosion: The Construction of Animal Biodiversity; Roberts & Co.: Greenwood, IN, USA, 2013. [Google Scholar]
- Lang, D.; Rensing, S.A. The evolution of transcriptional regulation in the viridiplantae and its correlation with morphological complexity. In Evolutionary Transitions to Multicellular Life. Principles and Mechanisms; Ruiz-Trillo, I., Nedelcu, A.M., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2015; pp. 301–334. [Google Scholar]
- Technau, U.; Genikhovich, G.; Kraus, J.E.M. Cnidaria. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Introduction, Non-Bilateria, Acoelomorpha, Xenoturbellida, Chaetognatha; Springer: Vienna, Austria, 2015; Volume 1, pp. 115–163. [Google Scholar]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Dev. Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuBuc, T.Q.; Bobkov, Y.; Ryan, J.; Martindale, M.Q. The radial expression of dorsal–ventral patterning genes in placozoans, Trichoplax adhaerens, argues for an oral-aboral axis. bioRxiv 2018. [Google Scholar] [CrossRef]
- Lee, P.N.; Pang, K.; Matus, D.Q.; Martindale, M.Q. A Wnt of things to come: Evolution of Wnt signaling and polarity in cnidarians. Sem. Cell Dev. Biol. 2006, 17, 157–167. [Google Scholar] [CrossRef]
- Hogvall, M.; Vellutini, B.C.; Martín-Durán, J.M.; Hejnol, A.; Budd, G.E.; Janssen, R. Embryonic expression of priapulid Wnt genes. Dev. Genes Evol. 2019, 229, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.P.; Reddien, P.W. Wnt signaling and the polarity of the primary body axis. Cell 2009, 139, 1056–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chipman, A.D. Hexapoda: Comparative aspects of early development. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Ecdysozoa III: Hexapoda; Springer: Vienna, Austria, 2015; Volume 5, pp. 92–109. [Google Scholar]
- Constantinou, S.J.; Duan, N.; Nagy, L.M.; Chipman, A.D.; Williams, T.A. Elongation during segmentation shows axial variability, low mitotic rates, and synchronized cell cycle domains in the crustacean, Thamnocephalus platyurus. EvoDevo 2020, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Neijts, R.; Simmini, S.; Giuliani, F.; van Rooijen, C.; Deschamps, J. Region-specific regulation of posterior axial elongation during vertebrate embryogenesis. Dev. Dyn. 2014, 243, 88–98. [Google Scholar] [CrossRef]
- Angerer, L.M.; Yaguchi, S.; Angerer, R.C.; Burke, R.D. The evolution of nervous system patterning: Insights from sea urchin development. Development 2011, 138, 3613–3623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holló, G.; Novák, M. The manoeuvrability hypothesis to explain the maintenance of bilateral symmetry in animal evolution. Biol. Direct. 2012, 7, 22. Available online: http://www.biology-direct.com/content/7/1/22 (accessed on 10 January 2021).
- He, S.; del Viso, F.C.-Y.; Ikmi, A.; Kroesen, A.E.; Gibson, M.C. An axial Hox code controls tissue segmentation and body patterning in Nematostella vectensis. Science 2018, 361, 1377–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolfi, A.; Brown, F.D. Tunicata. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; Volume 6, pp. 135–204. [Google Scholar]
- Peterson, K.J.; Eernisse, D.J. The phylogeny, evolutionary developmental biology, and paleobiology of the Deuterostomia: 25 years of new techniques, new discoveries, and new ideas. Org. Divers. Evol. 2016, 16, 401–418. [Google Scholar] [CrossRef]
- Jacobs, D.K.; Hughes, N.C.; Fitz-Gibbon, S.T.; Winchell, C.J. Terminal addition, the Cambrian radiation and the Phanerozoic evolution of bilaterian form. Evol. Dev. 2005, 7, 498–514. [Google Scholar] [CrossRef]
- Holland, P.W.H. Did homeobox gene duplications contribute to the Cambrian explosion? Zool. Lett. 2015, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Schier, A.F. Nodal morphogens. Cold Spring Harb. Perspect. Biol. 2009, 1, a003459. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Aldea, D.; Oulion, S.; Subirana, L.; de Lera, A.R.; Somorjai, I.; Escriva, H. Evolution of the role of RA and FGF signals in the control of somitogenesis in chordates. PLoS ONE 2015, 10, e0136587. [Google Scholar] [CrossRef] [Green Version]
- Shylo, N.A.; Ramrattan, D.; Weatherbee, S.D. Presence of midline cilia supersedes the expression of Lefty1 in forming the midline barrier during the establishment of left–right asymmetry. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Grande, C.; Patel, N.H. Lophotrochozoa get into the game: The nodal pathway and left/right asymmetry in Bilateria. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soukup, V. Left–right asymmetry specification in amphioxus: Review and prospects. Int. J. Dev. Biol. 2017, 61, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, D.E.K. The origin of the Hox/ParaHox genes, the Ghost Locus hypothesis and the complexity of the first animal. Brief. Funct. Genom. 2016, 15, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Lane, N. The Vital Question: Energy, Evolution, and the Origin of Complex Life; Profile Books: London, UK, 2015. [Google Scholar]
- Koonin, E.V. The Logic of Chance: The Nature and Origin of Biological Evolution; Pearson Education Inc.: Upper Saddle River, NJ, USA, 2012. [Google Scholar]
- Iglesias, M.; Gomez-Skarmeta, J.L.; Saló, E.; Adell, T. Silencing of Smed-beta-catenin1 generates radial-like hypercephalized planarians. Development 2008, 135, 1215–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baguña, J.; Martinez, P.; Paps, J.; Riutort, M. Back in time: A new systematic proposal for the Bilateria. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1481–1491. [Google Scholar] [CrossRef] [Green Version]
- Duboule, D. The rise and fall of Hox gene clusters. Development 2007, 134, 2549–2560. [Google Scholar] [CrossRef] [Green Version]
- Darbelay, F.; Duboule, D. Topological domains, metagenes, and the emergence of pleiotropic regulations at Hox loci. Curr. Top. Dev. Biol. 2016, 116, 299–314. [Google Scholar]
- Halanych, K.M. How our view of animal phylogeny was reshaped by molecular approaches: Lessons learned. Org. Divers. Evol. 2016, 16, 319–328. [Google Scholar] [CrossRef]
- Duboule, D. Temporal colinearity and the phylotypic progression: A basis for the stability of a vertebrate Bauplan and the evolution of morphologies through heterochrony. Development 1994, 1994, 135–142. [Google Scholar]
- Tschopp, P.; Duboule, D. A Genetic approach to the transcriptional regulation of Hox gene clusters. Annu. Rev. Genet. 2011, 45, 145–166. [Google Scholar] [CrossRef]
- Sánchez-Herrero, E. Hox targets and cellular functions. Scientifica 2013, 738257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.-M.; Kanda, M.; Satoh, N.; Furuya, H. The phylogenetic position of dicyemid mesozoans offers insights into spiralian evolution. Zool. Lett. 2017, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Zverkov, O.A.; Mikhailov, K.V.; Isaev, S.V.; Rusin, L.Y.; Popova, O.V.; Logacheva, M.D.; Penin, A.A.; Moroz, L.L.; Panchin, Y.V.; Lyubetsky, V.A.; et al. Dicyemida and Orthonectida: Two stories of body plan simplification. Front. Genet. 2019, 10, 443. [Google Scholar] [CrossRef] [PubMed]
- Høeg, J.T.; Deutsch, J.; Chan, B.K.K.; Le Semmler, H. “Crustacea”: Cirripedia. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Ecdysozoa II: “Crustacea”; Springer: Vienna, Austria, 2015; Volume 4, pp. 153–181. [Google Scholar]
- Shukalyuk, A.I.; Isaeva, V.V. Molecular and subcellular gametogenic machinery of stem and germline cells across Metazoa. In Current Frontiers and Perspectives in Cell Biology; Najman, S., Ed.; InTech: Rieka, Croatia, 2012; pp. 279–314. [Google Scholar]
- Shukalyuk, A.I.; Isaeva, V.V. Molecular signature and sub-cellular machinery of metazoan gametogenic stem cells. In Recent Advances in Germ Cells Research; Perrotte, A., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 1–40. [Google Scholar]
- Mullins, R.D. Cytoskeletal mechanisms for breaking cell symmetry. Cold Spring Harb. Perspect. Biol. 2010, 2, a003392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, F. Evolution and tinkering. Science 1977, 196, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Fedonkin, M.A. The origin of the Metazoa in the light of the Proterozoic fossil records. Palentol. Res. 2003, 7, 9–41. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, A.; Hester, S.D.; Constantinou, S.J.; Blaine, W.G.; Tewksbury, A.B.; Matei, M.T.; Nagy, L.M.; Williams, T.A. Changing cell behaviours during beetle embryogenesis correlates with slowing of segmentation. Nat. Commun. 2015, 6, 6635. [Google Scholar] [CrossRef] [Green Version]
- Maroto, M.; Bone, R.A.; Dale, J.K. Somitogenesis. Development 2012, 139, 2453–2456. [Google Scholar] [CrossRef] [Green Version]
- Oates, A.C.; Morelli, L.G.; Ares, S. Patterning embryos with oscillations: Structure, function and dynamics of the vertebrate segmentation clock. Development 2012, 139, 625–639. [Google Scholar] [CrossRef] [Green Version]
- Aulehla, A.; Pourquié, O. Signaling gradients during paraxial mesoderm development. Cold Spring Harb. Perspect. Biol. 2010, 2, a000869. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Saunders, T.E. Shh induces symmetry breaking in the presomitic mesoderm by inducing tissue shear and orientated cell rearrangements. bioRxiv 2019. [Google Scholar] [CrossRef]
- Pueyo, J.I.; Lanfear, R.; Couso, J.P. Ancestral Notch-mediated segmentation revealed in the cockroach Periplaneta americana. Proc. Natl. Acad. Sci. USA 2008, 105, 16614–16619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrazin, A.F.; Peel, A.D.; Averof, M. A segmentation clock with two-segment periodicity in insects. Science 2012, 336, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Jockusch, E.L.; Smith, F.W. Hexapoda: Comparative aspects of later embryogenesis and metamorphosis. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Ecdysozoa III: Hexapoda; Springer: Vienna, Austria, 2015; Volume 5, pp. 111–208. [Google Scholar]
- Zakany, J.; Kmita, M.; Alarcon, P.; De La Pompa, J.L.; Duboule, D. Localized and transient transcription of Hox genes suggests a link between patterning and the segmentation clock. Cell 2001, 106, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Holland, L.Z. Cephalochordata. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Deuterostomia; Springer: Vienna, Austria, 2015; Volume 6, pp. 91–134. [Google Scholar]
- Martin, B.L.; Kimelman, D. Wnt signaling and the evolution of embryonic posterior development. Curr. Biol. 2009, 19, R215–R219. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.; Martinez, P.; Morris, V. Evolution of a pentameral body plan was not linked to translocation of anterior Hox genes: The echinoderm HOX cluster revisited. Evol. Dev. 2016. [Google Scholar] [CrossRef]
- Cavodeassi, F.; ·Creuzet, S.; Etchevers, H.C. The hedgehog pathway and ocular developmental anomalies. Hum. Genet. 2019, 138, 917–936. [Google Scholar] [CrossRef] [Green Version]
- Abramyan, J. Hedgehog signaling and embryonic craniofacial disorders. J. Dev. Biol. 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Ertzer, R.; Müller, F.; Hadzhiev, Y.; Vinothkumar, S. Cooperation of sonic hedgehog enhancers in midline expression. Dev. Biol. 2006, 301, 578–589. [Google Scholar] [CrossRef]
- Lumsden, A.; Graham, A. A forward role for Hedgehog. Curr. Biol. 1995, 5, 1347–1350. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Kinsman, S.; Makris, N.; Grant, E.; Haselgrove, C.; McInerney, S.; Kennedy, D.N.; Takahashi, T.; Fredrickson, K.; Mori, S.; et al. Semilobar holoprosencephaly with midline ‘seam’: A topologic and morphogenetic model based upon MRI analysis. Cereb. Cortex 2003, 13, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Yadav, J.; Gard, E. Cyclopia syndrome. BMJ Case Rep. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Fernàndez, J.; Benito-Gutiérrez, E. It’s a long way from amphioxus: Descendants of the earliest chordate. BioEssays 2009, 31, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Wanninger, A.; Wollesen, T. Mollusca. In Evolutionary Developmental Biology of Invertebrates; Wanninger, A., Ed.; Springer: Vienna, Austria, 2015; Volume 2, pp. 103–153. [Google Scholar]
- Vopalensky, P.; Tosches, M.A.; Achim, K.; Handberg-Thorsager, M.; Arendt, D. From spiral cleavage to bilateral symmetry: The developmental cell lineage of the annelid brain. BMC Biol. 2019, 17, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, R.; Endo, B.; Masanri, A.; Shimuzu, M. Chiral blastomere arrangement dictates zygotic left–right asymmetry pathway in snails. Nature 2009, 462, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. Experimental Embryology; Columbia Univ. Press: New York, NY, USA, 1927. [Google Scholar]
- Murray, J.; Clarke, B. The inheritance of polymorphic shell characters in Partula (Gastropoda). Genetics 1966, 54, 1261–1277. [Google Scholar] [CrossRef]
- Tee, Y.H.; Shemesh, T.; Thiagarajan, V.; Hariadi, R.F.; Anderson, K.L.; Page, C.; Volkmann, N.; Hanein, D.; Sivaramakrishnan, S.; Kozlov, M.M.; et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat. Cell Biol. 2015, 17, 445–457. [Google Scholar] [CrossRef]
- Mandelbrot, B.B. Fractals: Form, Chance, and Dimension; W. H. Freeman: San Francisco, CA, USA, 1977. [Google Scholar]
- Weibel, E.R. Fractal geometry–a design principle for living organisms. Am. J. Physiol. 1991, 261, 361–369. [Google Scholar] [CrossRef]
- Peitgen, H.-O.; Jürgens, H.; Saupe, D. Fractals for the Classroom; Spriger: Berlin, Germany, 1992. [Google Scholar]
- Davies, J.A. Why a book on branching, and why now? Afterword. In Branching Morphogenesis; Davies, J.A., Ed.; Springer: Boston, MA, USA, 2006. [Google Scholar]
- Høeg, J.T.; Lützen, J. Life cycle and reproduction in the Cirripedia Rhizocephala. Oceanogr. Mar. Biol. Ann. Rev. 1995, 33, 427–485. [Google Scholar]
- Glenner, H.; Lützen, J.; Takahashi, T. Molecular and morphological evidence for a monophyletic clade of asexually reproducing Rhizocephala: Polyascus, new genus (Cirripedia). J. Crustacean Res. 2003, 23, 548–557. [Google Scholar] [CrossRef] [Green Version]
- Isaeva, V.V. Fractal and chaotic patterns in animal morphogenesis. Transact. Zool. Inst. RAN 2009, 1, 199–218. (In Russian) [Google Scholar]
- Isaeva, V.V.; Akhmadieva, A.V.; Shukalyuk, A.I. The hidden coloniality at the parasitic stage in Peltogaster reticulatus (Crustacea: Rhizocephala). J. Mar. Biol. Assoc. UK 2012, 92, 457–462. [Google Scholar] [CrossRef]
- Metzger, R.J.; Klein, O.D.; Martin, G.R.; Krasnow, M.A. The branching programme of mouse lung development. Nature 2008, 453, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannezo, E.; Scheele, C.L.G.J.; Moad, M.; Drogo, N.; Heer, R.; Sampogna, R.V.; van Rheenen, J.; Simons, B.D. A unifying theory of branching morphogenesis. Cell 2017, 171, 242–255. [Google Scholar] [CrossRef] [Green Version]
- Damiani, G. Evolutionary meaning, functions and morphogenesis of branching structures in biology. In Fractals in Biology and Medicine; Nonnenmacher, T.F., Losa, G.A., Weibel, E.R., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1994; pp. 104–115. [Google Scholar]
- Fleury, V.; Watanabe, T.; Nguyen, T.-H.; Unbekandt, M.; Warburton, D.; Dejmek, M.; Nguyen, M.B.; Lindner, A.; Schwartz, L. Physical mechanisms of branching morphogenesis in animals: From viscous fingering to cartilage rings. In Branching Morphogenesis; Davies, J.A., Ed.; Eurekah.com and Springer Science+Business Media: New York, NY, USA, 2006; pp. 202–234. [Google Scholar]
- Warburton, D.; El-Hashash, A.; Carraro, G.; Tiozzo, C.; Sala, F.; Rogers, O.; De Langhe, S.; Kemp, P.J.; Riccardi, D.; Torday, J.; et al. Lung organogenesis. Curr. Top. Dev. Biol. 2010, 90, 73–158. [Google Scholar] [PubMed]
- Chuang, P.; McMahon, A. Branching morphogenesis of the lung: New molecular insights into an old problem. Trends Cell. Biol. 2003, 13, 86–91. [Google Scholar] [CrossRef]
- Menshykau, D.; Kraemer, C.; Iber, D. Branch mode selection during early lung development. PLoS Comput. Biol. 2012, 8, e1002377. [Google Scholar] [CrossRef] [Green Version]
- Chernyshev, A.V.; Isaeva, V.V. Formation of the chaotic patterns of gastro-vascular systems of the jellyfish Aurelia aurita in ontogeny. Russ. J. Mar. Biol. 2002, 28, 377–381. [Google Scholar]
- Isaeva, V.V.; Karetin, Y.A.; Chernyshev, A.V.; Shkuratov, D.Y. Fractals and Chaos in Biological Morphogenesis; Dalnauka: Vladivostok, Russia, 2004. (In Russian) [Google Scholar]
- Isaeva, V.V.; Pushchina, E.V.; Karetin, Y.A. Changes in morphometric parameters and fractal dimensions of spinal marrow neurons in ontogeny of the cherry salmon Oncorhynchus masou. Russ. J. Mar. Biol. 2006, 32, 106–114. [Google Scholar] [CrossRef]
- Savriama, Y.; Vitulo, M.; Gerber, S.; Debat, V.; Fusco, G. Modularity and developmental stability in segmented animals: Variation in translational asymmetry in geophilomorph centipedes. Dev. Genes Evol. 2016. [Google Scholar] [CrossRef]
- Rasskin-Gutman, D.; Izpisua-Belmonte, J.C. Theoretical morphology of developmental asymmetries. BioEssays 2004, 26, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J.; Harper, D.A.T. Introduction to Paleobiology and the Fossil Records; Wiley-Blackwell: Oxford, UK, 2009. [Google Scholar]
- Gould, S.J. Wonderful Life. The Burgess and the Nature of History; Norton Co: New York, NY, USA; London, UK, 1989. [Google Scholar]
- Smith, M. Fossil focus–Hallucigenia and the evolution of animal body plans. Palaeontology 2017, 5, 1–9, Online. [Google Scholar]
- Budd, G.E.; Jackson, I.S.C. Ecological innovations in the Cambrian and the origins of the crown group phyla. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, S.J. The Structure of Evolutionary Theory; Harvard University Press: Cambridge, MA, USA; London, UK, 2002. [Google Scholar]
- Fedonkin, M.A. Metameric features in the Vendian metazoans. Ital. J. Zool. 1998, 65, 11–27. [Google Scholar] [CrossRef]
- Davidson, E.H.; Erwin, D.H. An integrated view of precambrian eumetazoan evolution. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.D.; Hughes, I.V.; Gehling, J.G.; Droser, M.L. Discovery of the oldest bilaterian from the Ediacaran of South Australia. Proc. Natl. Acad. Sci. USA 2020, 117, 7845–7850. [Google Scholar] [CrossRef]
- Giribet, G. New animal phylogeny: Future challenges for animal phylogeny in the age of phylogenomics. Org. Divers. Evol. 2016, 16, 419–426. [Google Scholar] [CrossRef]
- Xiao, S.; Laflamme, M. On the eve of animal radiation: Phylogeny, ecology and evolution of the Ediacara biota. Trends Ecol. Evol. 2008, 24, 31–40. [Google Scholar] [CrossRef]
- Seilacher, A. The nature of vendobionts. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 387–397. [Google Scholar]
- Fedonkin, M.A. Precambrian metazoans: The problems of preservation, systematics and evolution. Philos. Trans. R. Soc. Lond. B 1985, 311, 27–45. [Google Scholar]
- Ivantsov, A.Y. The most probable Eumetazoa among late Precambrian macrofossils. Invertebr. Zool. 2017, 14, 127–133. [Google Scholar] [CrossRef]
- Ivantsov, A.Y.; Zakrevskaya, M.A.; Nagovitsyn, A.L. Morphology of integuments of the Precambrian animals, Proarticulata. Invertebr. Zool. 2019, 16, 19–26. [Google Scholar] [CrossRef]
- Evans, S.D.; Droser, M.L.; Gehling, J.G. Highly regulated growth and development of the Ediacara macrofossil Dickinsonia costata. PLoS ONE 2017, 12, e0176874. [Google Scholar] [CrossRef] [PubMed]
- Muscente, A.D.; Boag, T.H.; Bykova, N.; Schiffbauer, J.D. Environmental disturbance, resource availability, and biologic turnover at the dawn of animal life. Earth-Sci. Rev. 2018, 177, 248–264. [Google Scholar] [CrossRef]
- Antcliffe, J.B.; Brasier, M.D. Towards a morphospace for the Ediacara biota. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 377–386. [Google Scholar]
- Tojo, B.; Saito, R.; Kawakami, S.; Ohno, T. Theoretical morphology of quilt structures in Ediacaran fossils. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 399–404. [Google Scholar]
- Hoyal Cuthill, J.F.; Conway Morris, S. Fractal branching organizations of Ediacaran rangeomorph fronds reveal a lost Proterozoic body plan. Proc. Natl. Acad. Sci. USA 2014, 111, 13122–13126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivantsov, A.Y.; Leonov, M.V. The Imprints of Vendian Animals–Unique Paleontological Objects of the Arkhangelsk Region; Arkhangelsk, Russia. 2008. Available online: https://www.researchgate.net/publication/260191573 (accessed on 10 January 2021). (In Russian).
- Valentine, J.W. Seeing ghosts: Neoproterozoic bilaterian body plans. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarover, P., Eds.; Geological Society: London, UK, 2007; Volume 286, pp. 369–375. [Google Scholar]
- Smith, A.B.; Zamora, S. Cambrian spiral-plated echinoderms from Gondwana reveal the earliest pentaradial body plan. Proc R Soc B 2013, 280, 20131197. [Google Scholar] [CrossRef] [Green Version]
- Topper, T.P.; Guo, J.; Clausen, S.; Skovsted, C.B.; Zhang, Z. A stem group echinoderm from the basal Cambrian of China and the origins of Ambulacraria. Nat. Commun. 2019. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isaeva, V.V.; Kasyanov, N.V. Symmetry Transformations in Metazoan Evolution and Development. Symmetry 2021, 13, 160. https://doi.org/10.3390/sym13020160
Isaeva VV, Kasyanov NV. Symmetry Transformations in Metazoan Evolution and Development. Symmetry. 2021; 13(2):160. https://doi.org/10.3390/sym13020160
Chicago/Turabian StyleIsaeva, Valeria V., and Nickolay V. Kasyanov. 2021. "Symmetry Transformations in Metazoan Evolution and Development" Symmetry 13, no. 2: 160. https://doi.org/10.3390/sym13020160
APA StyleIsaeva, V. V., & Kasyanov, N. V. (2021). Symmetry Transformations in Metazoan Evolution and Development. Symmetry, 13(2), 160. https://doi.org/10.3390/sym13020160