
Brain Size Associated with Foot Preferences in Australian Parrots



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Source of Data
2.3. Statistical Tests
3. Results
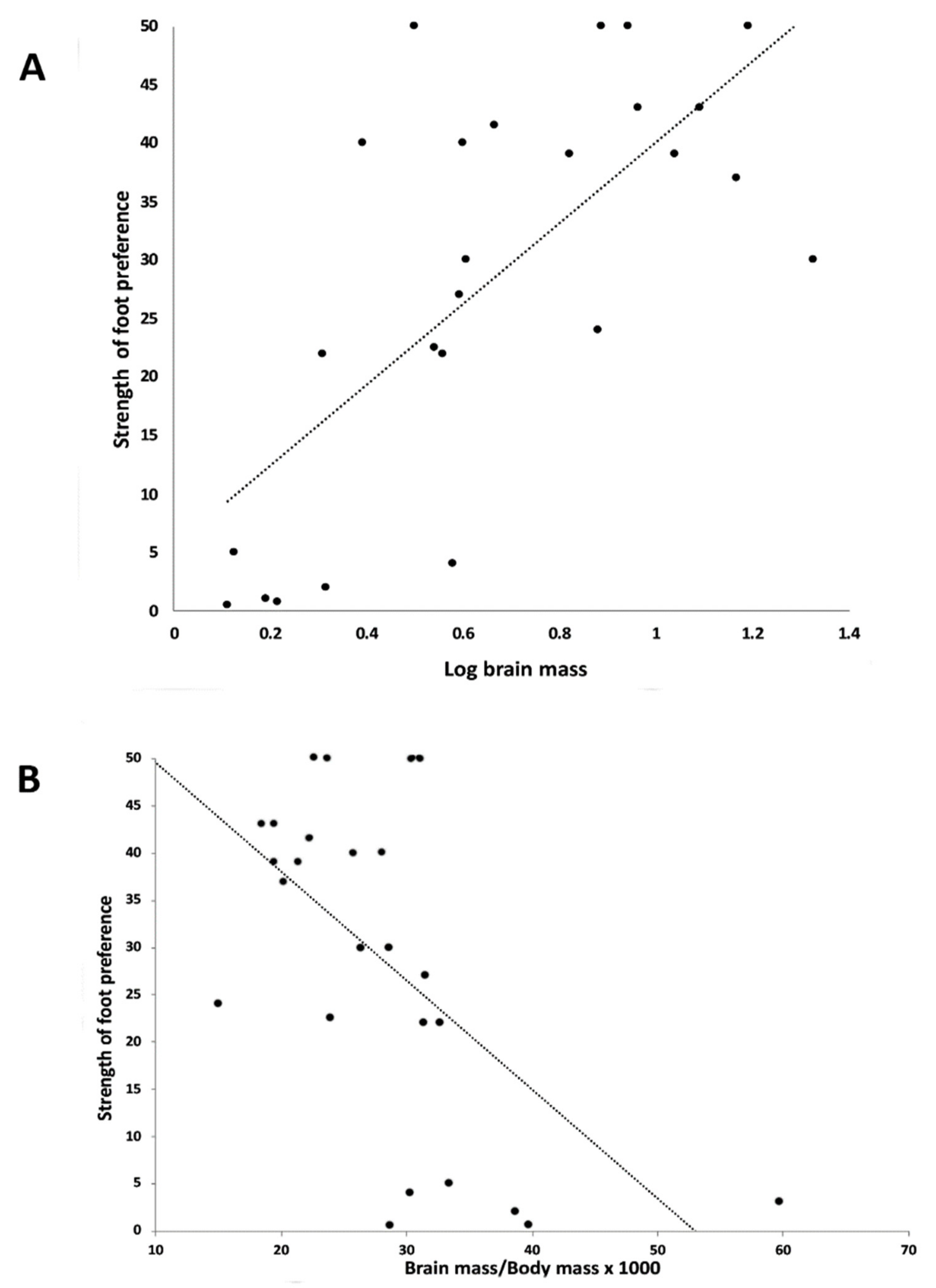
3.1. Association between Foot Preference and Body Mass
3.2. Association between Foot Preference and Brain Mass
3.3. Association between Foot Preference and Brain Mass Relative to Body Mass
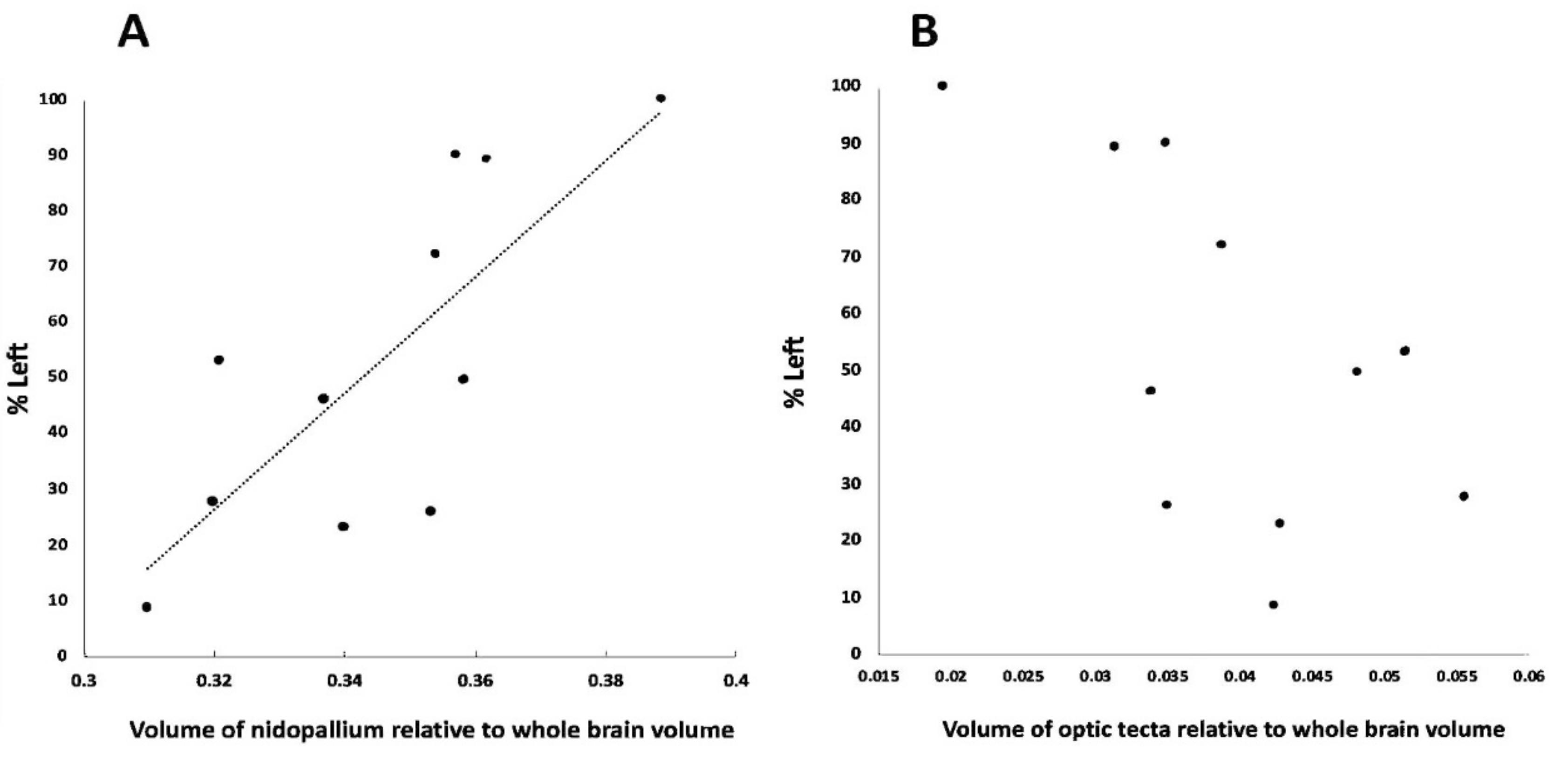
3.4. Association between Foot Preference and Brain Mass in the Smaller Sample
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ocklenburg, S.; Metzen, D.; Schlüter, C.; Fraenz, C.; Arning, L.; Streit, F.; Güntürkün, O.; Kumsta, R.; Genc, E. Polygenic scores for handedness and their association with asymmetries in brain structure. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Dragovic, M.; Hammond, G. Handedness in schizophrenia: A quantitative review of evidence. Acta Psychiatr. Scand. 2005, 111, 410–419. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Güntürkün, O.; Hugdahl, K.; Hirnstein, M. Laterality and mental disorders in the postgenomic age—A closer look at schizophrenia and language lateralization. Neurosci. Biobehav. Rev. 2015, 59, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Packheiser, J.; Schmitz, J.; Stein, C.C.; Pfeifer, L.S.; Berretz, G.; Papadatou-Pastou, M.; Peterburs, J.; Ocklenburg, S. Handedness and depression: A meta-analysis across 87 studies. PsyArXiv 2021. [Google Scholar] [CrossRef]
- Sommer, I.E.C.; Aleman, A.; Ramsey, N.; Bouma, A.; Kahn, R. Handedness, language lateralisation and anatomical asymmetry in schizophrenia—Meta-analysis. Br. J. Psychiatr. 2001, 178, 344–351. [Google Scholar] [CrossRef]
- Chance, S.A.; Crow, T.J. Distinctively human: Cerebral lateralisation and language in Homo sapiens. J. Anthropol. Sci. 2007, 85, 83–100. [Google Scholar]
- McGrew, W.C.; Marchant, L.F. On the other hand: Current issues in and meta-analysis of the behavioral laterality of hand function in nonhuman primates. Yearb. Phys. Anthropol. 1997, 40, 201–232. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Cantalupo, C. Handedness in chimpanzees (Pan troglodytes) is associated with asymmetries of the primary motor cortex but not with homologous language areas. Behav. Neurosci. 2004, 118, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Hook, M.A.; Rogers, L.J. Visuospatial reaching preferences of common marmosets: An assessment of individual biases across a variety of tasks. J. Comp. Psychol. 2008, 122, 41–51. [Google Scholar] [CrossRef]
- Hook-Costigan, M.A.; Rogers, L.J. Eye preferences in common marmosets (Callithrix jacchus): Influence of age, stimulus and hand preference. Laterality 1998, 3, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.J.; Rogers, L.J. Cognitive bias, hand preference and welfare in common marmosets. Behav. Brain Res. 2015, 287, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Ströckens, F.; Güntürkün, O.; Ocklenburg, S. Limb preferences in non-human vertebrates. Laterality 2013, 18, 536–575. [Google Scholar] [CrossRef]
- Crow, T.J. Why cerebral asymmetry is the key to the origin of Homo sapiens: How to find the gene of eliminate the theory. Curr. Psychol. Cogn. 1998, 17, 1237–1277. [Google Scholar]
- Corballis, M.C. Cerebral asymmetry and human uniqueness. In The Evolution of Hemispheric Specialization in Primates; Special Topics in Primatology; Hopkins, W.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 5, pp. 1–21. [Google Scholar]
- Corballis, M.C. Bilaterally symmetrical: To be or not to be? Symmetry 2020, 12, 326. [Google Scholar] [CrossRef] [Green Version]
- Corballis, M.C. Humanity and the left hemisphere: The story of half a brain. Laterality 2020, 26. [Google Scholar] [CrossRef]
- Malashichev, Y.B.; Wassersug, R.J. Left and right in the amphibian world: Which way to develop and where to turn? BioEssays 2004, 26, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Stancher, G.; Sovrano, V.A.; Vallortigara, G. Motor asymmetries in fishes, amphibians, and reptiles. Proc. Brain Res. 2018, 238, 33–56. [Google Scholar]
- Randler, C. Foot preferences during resting in wildfowl and waders. Laterality 2007, 12, 191–197. [Google Scholar] [CrossRef]
- Yu, G.; Guo, J.; Xie, W.; Wang, J.; Wu, Y.; Zhang, J.; Xu, J.; Li, J. Footedness predicts escape performance in a passerine bird. Ecol. Evol. 2020, 10, 4251–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, L.J. Lateralisation in the avian brain. Bird Behav. 1980, 2, 1–12. [Google Scholar] [CrossRef]
- Brown, C.; Magat, M. The evolution of lateralized foot use in parrots: A phylogenetic approach. Behav. Ecol. 2011, 22, 1201–1208. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.J. Laterality in animals. Int. J. Comp. Psychol. 1989, 3, 5–25. [Google Scholar]
- Magrath, D.I. Footedness in the glossy black-cockatoo: Some observations and a review of the literature with a note on the husking of Allocasuarina cones by this species. Corella 1994, 18, 21–24. [Google Scholar]
- Woodall, P.F.; Woodall, L.B. Little and long-billed Corellas feeding on hoop pine seeds, and their ‘footedness’. Sunbird J. Qld. Ornithol. Soc. 2001, 31, 30–32. [Google Scholar]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Vallortigara, G.; Rogers, L.J. A function for the bicameral mind. Cortex 2020, 124, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Strökens, F.; Ocklenburg, S. Brain lateralization: A comparative perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef] [PubMed]
- Ocklenburg, S.; Güntürkün, O. The Lateralized Brain: The Neuroscience and Evolution of Hemispheric Asymmetries; Academic Press: London, UK, 2018. [Google Scholar]
- Magat, M.; Brown, C. Laterality enhances cognition in Australian parrots. Proc. R. Soc. B 2009, 276, 4155–4162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cussen, V.A.; Mench, J.A. Performance on the Hamilton search task, and the influence of lateralization, in captive orange-winged Amazon parrots (Amazona amazonica). Anim. Cogn. 2014, 17, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Joseph, L.; Toon, A.; Schirtzinger, R.E.; Wright, T.F.; Schodde, R. A revised nomenclature and classification for family-group taxa of parrots (Psittaciformes). Zootaxa 2012, 40, 26–40. [Google Scholar] [CrossRef]
- Barker, F.K.; Cibois, A.; Schikler, P.A.; Feinstein, J.; Cracraft, J. Phylogeny and diversification of the largest avian radiation. Proc. Nat. Acad. Sci. USA 2004, 101, 11040–11045. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.V.; Bowles, W.E. Out of Gondwana: The origin of passerine birds. Trends Ecol. Evol. 2002, 17, 347–349. [Google Scholar] [CrossRef]
- Wright, T.F.; Schirtzinger, E.E.; Matsumoto, T.; Eberhard, J.R.; Graves, G.R.; Sanchez, J.J.; Capelli, S.; Müller, H.; Scharpegge, J.; Chambers, G.K.; et al. A multilocus molecular phylogeny of the parrots (Psittaciformes): Support for a Gondwanan origin during the Cretaceous. Mol. Biol. Evol. 2008, 25, 2141–2156. [Google Scholar] [CrossRef] [PubMed]
- Olkowicz, S.; Kocourek, M.; Lučan, R.K.; Porteš, M.; Fitch, W.T.; Herculano-Houzel, S.; Němec, P. Birds have primate-like numbers of neurons in the forebrain. Proc. Natl. Acad. Sci. USA 2016, 113, 7255–7260. [Google Scholar] [CrossRef] [Green Version]
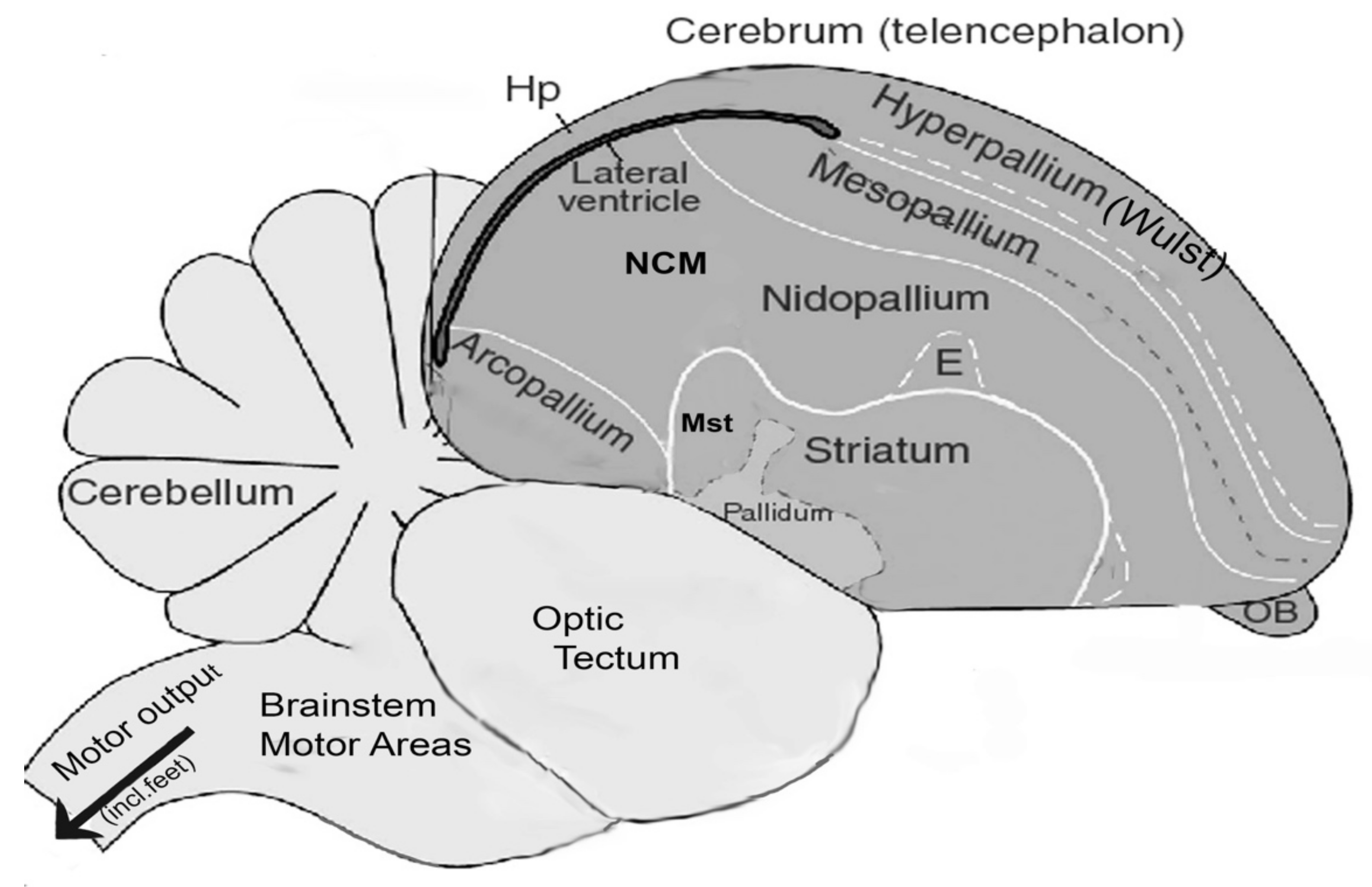
- Kuenzel, W.J. The Avian subpallium and autonomic nervous system. In Sturkie’s Avian Physiology, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 135–163. [Google Scholar]
- Chakraborty, M.; Jarvis, E.D. Brain evolution by brain pathway duplication. Philos. Trans. R. Soc. Lond. B 2015, 370, 20150056. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, M.; Walløe, S.; Nedergaard, S.; Fridel, E.E.; Dabelsteen, T.; Pakkenberg, B.; Bertelsen, M.F.; Dorrestein, G.M.; Brauth, S.E.; Durand, S.; et al. Core and shell song systems unique to the parrot brain. PLoS ONE 2015, 10, e0118496. [Google Scholar] [CrossRef]
- Wild, J.M.; Williams, M.N. Rostral Wulst in passerine birds. 1. Origin, course, and terminations of an avian pyramidal tract. J. Comp. Neurol. 2000, 416, 429–450. [Google Scholar] [CrossRef]
- Wild, J.M. Neural pathways for the control of birdsong production. J. Neurobiol. 1997, 33, 653–670. [Google Scholar] [CrossRef]
- Güntürkün, O. The avian ’prefrontal cortex’ and cognition. Curr. Opin. Neurobiol. 2005, 15, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O. The convergent evolution of neural substrates for cognition. Psychol. Res. 2012, 76, 212–219. [Google Scholar] [CrossRef]
- Herold, C.; Palomero-Gallagher, N.; Hellmann, B.; Kröner, S.; Theiss, C.; Güntürkün, O.; Zilles, K. The receptor architecture of the pigeons’ nidopallium caudolaterale: An avian analogue to the mammalian prefrontal cortex. Brain Struct. Funct. 2011, 216, 239–254. [Google Scholar] [CrossRef]
- Von Eugen, K.; Tabrik, S.; Güntürkün, O.; Strökens, F. A comparative analaysis of the dopaminergic innervation of the executive caudal nidopallium in pigeon, chicken, zebra finch, and carrion crow. J. Comp. Neurol. 2020, 528, 2929–2955. [Google Scholar] [CrossRef]
- Kröner, S.; Güntürkün, O. Afferent and efferent connections of the caudolateral neostriatum in the pigeon (Columba livia): A retro-and antero-grade pathway tracing study. J. Comp. Neurol. 1999, 407, 228–260. [Google Scholar] [CrossRef]
- Dykes, M.; Klarer, A.; Porter, B.; Rose, J.; Colombo, M. Neurons in the pigeon nidopallium caudolaterale display value-related activity. Sci. Rep. 2018, 8, 5377. [Google Scholar] [CrossRef] [PubMed]
- Kalenscher, T.; Windmann, S.; Diekamp, B.; Rose, J.; Güntürkün, O.; Colombo, M. Single units in the pigeon brain integrate reward amount and time-to-reward in an impulsive choice task. Curr. Biol. 2005, 15, 594–602. [Google Scholar] [CrossRef]
- Kuenzel, W.J.; Medina, L.; Csillag, A.; Perkel, D.J.; Reiner, A. The avian subpallium: New insights into structural and functional subdivisions occupying the lateral subpallial wall and their embryological origins. Brain Res. 2011, 1424, 67–101. [Google Scholar] [CrossRef] [Green Version]
- Reiner, A.; Perkel, D.J.; Bruce, L.L.; Butler, A.B.; Csillag, A.; Kuenzel, W.; Medina, L.; Paxinos, G.; Shimizu, T.; Striedter, G.; et al. Revised nomenclature for avian telencephalon and some related brainstem nuclei. J Comp. Neurol. 2004, 473, 377–414. [Google Scholar] [CrossRef]
- Niederleitner, B.; Gutierrez-Ibanez, C.; Krabichler, Q.; Weigel, S.; Luksch, H. A novel relay nucleus between the inferior colliculus and the optic tectum in the chicken (Gallus gallus). J. Comp. Neurol. 2017, 525, 513–534. [Google Scholar] [CrossRef]
- Brown, C.; Magat, M. Cerebral lateralization determines hand preferences in Australian parrots. Biol. Letts 2011, 7, 496–498. [Google Scholar] [CrossRef] [PubMed]
- Franklin, D.C.; Garnett, S.T.; Luck, G.W.; Gutierrez-Ibanez, C.; Iwaniuk, A.N. Relative brain size in Australian birds. Emu 2014, 114. [Google Scholar] [CrossRef]
- Iwaniuk, A.N.; Dean, K.M.; Nelson, J.E. Interspecific allometry of the brain and brain regions in parrots (Psittaciformes): Comparisons with other birds and primates. Brain Behav. Evol. 2005, 65, 40–59. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Workman, L. Footedness in birds. Anim. Behav. 1993, 45, 409–411. [Google Scholar] [CrossRef] [Green Version]
- Iwaniuk, A.N.; Hurd, P.L. The evolution of cerebrotypes in birds. Brain Behav. Evol. 2005, 65, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Lateralization in its many forms, and its evolution and development. In The Evolution of Hemispheric Specialization in Primates; Special Topics in Primatology; Hopkins, W.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 5, pp. 23–56. [Google Scholar]
- Andrews, C.B.; Gregory, T.R. Genome size is inversely correlated with relative brain size in parrots and cockatoos. Genome 2009, 52, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Healy, S.D.; Rowe, C. A critique of comparative brain studies of brain size. Proc. R. Soc. B 2007, 274, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Willemet, R. Reconsidering the evolution of brain, cognition, and behavior in birds and mammals. Front. Psychol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herculano-Houzel, S. Numbers of neurons as biological correlates of cognitive capability. Curr. Opin. Behav. Sci. 2017, 16, 1–7. [Google Scholar] [CrossRef]
- Güntürkün, O.; Hellmann, B.; Melsbach, G.; Prior, H. Asymmetries of representation in the visual system of pigeons. Neuroreport 1998, 9, 4127–4130. [Google Scholar] [CrossRef] [Green Version]
- Güntürkün, O.; Stüttgen, M.C.; Manns, M. Pigeons as a model species for cognitive neuroscience. Neuroforum 2014, 5, 86–92. [Google Scholar] [CrossRef]
- Morandi-Raikova, A.; Danieli, K.; Lorenzi, E.; Rosa-Salva, O.; Mayer, U. Anatomical asymmetries in the tectofugal pathway of dark-incubated domestic chicks: Rightwards lateralization of parvalbumin neurons in the entopallium. Laterality 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Güntürkün, O. The commissura anterior compensates asymmetries of visual representation in pigeons. Laterality 2021, 26. [Google Scholar] [CrossRef]
- Striedter, G.F.; Charvet, C.J. Developmental origins of species differences in telencephalon and tectum size: Morphometric comparisons between a par-akeet (Melopsittacus undulatus) and a quail (Colinus virgianus). J. Comp. Neurol. 2008, 507, 1663–1675. [Google Scholar] [CrossRef]
- Lee, H.J.; Schneider, R.F.; Manousaki, T.; Kang, J.H.; Lein, E.; Franchini, P.; Meyer, A. Lateralized feeding behavior is associated with asymmetrical neuroanatomy and lateralized gene expressions in the brain in scale-eating cichlid fish. Genome Biol. Evol. 2017, 9, 3122–3136. [Google Scholar] [CrossRef] [Green Version]
- MacNeilage, P.; Rogers, L.J.; Vallortigara, G. Origins of the left and right brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [CrossRef]
- Kaplan, G. Play behaviour, not tool using, relates to brain mass in a sample of birds. Sci. Rep. 2020, 10, 20437. [Google Scholar] [CrossRef]
- Zucca, P.; Sovrano, V.A. Animal lateralization and social recognition: Quails use their left visual hemifield when approaching a companion and their right visual hemifield when approaching a stranger. Cortex 2008, 44, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Salva, O.R.; Regolin, L.; Mascalzoni, E.; Vallortigara, G. Cerebral and behavioural asymmetries in animal social recognition. Comp. Cogn. Behav. Rev. 2012, 7, 110–138. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Aust, U.; Huber, L.; Hausmann, M.; Güntürkün, O. Lateralized cognition: Asymmetrical and complementary strategies of pigeons during discrimination of the “human concept”. Cognition 2007, 104, 315–344. [Google Scholar] [CrossRef]
- Kaplan, G. Audition and hemispheric specialization in songbirds and new evidence from Australian magpies. Symmetry 2017, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Costalunga, G.; Kobylkov, D.; Rosa-Salva, O.; Vallortigara, G.; Mayer, U. Light-Incubation effects on lateralisation of single unit responses in the visual Wulst of domestic chicks. Brain Struct. Funct. 2021. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Latin Species Name | No | % Left | Strength | Brain Mass /Body Mass × 1000 |
|---|---|---|---|---|---|
| Galah | Eolophus roseicapella | 58 | 89 | 39 | 21.3509 |
| Sulphur-crested cockatoo | Cacatua galerita | 98 | 87 | 37 | 20.2646 |
| Little corella | Cacatua sanguinea | 14 | 93 | 43 | 19.5567 |
| Long-billed corella | Cacatua tenuirostris | 17 | 89 | 39 | 19.5098 |
| Pink cockatoo | Cacatua leadbeateri | 24 | 100 | 50 | 22.6742 |
| Yellow-tailed black cockatoo | Calyptorhynchus funereus | 7 | 100 | 50 | 23.6710 |
| Gang-gang cockatoo | Callocephalon fimbriatum | 38 | 100 | 50 | 30.9971 |
| Yellow rosella | Platycercus f. flaveolus | 6 | 100 | 50 | 30.6296 |
| Crimson rosella | Platycercus elegans | 17 | 23 | 27 | 31.5813 |
| Budgerigar | Melopsittacus undulatus | 9 | 51 | 1 | 59.7692 |
| Cockatiel | Nymphicus hollandicus | 20 | 90 | 40 | 28.0889 |
| Red-tailed black cockatoo | Calyptorhynchus banksii | 20 | 93 | 43 | 18.4779 |
| King parrot | Alisterus scapularis | 20 | 8.5 | 41.5 | 22.4216 |
| Palm cockatoo | Probosciger aterrimus | 5 | 80 | 30 | 26.3499 |
| Eclectus parrot | Eclectus roratus | 20 | 26 | 24 | 15.0394 |
| Turquoise parrot | Neophema pulchella | 10 | 45 | 5 | 33.4110 |
| Red-winged parrot | Aprosmictus erythropterus | 10 | 10 | 40 | 25.7329 |
| Australian ringneck | Barnardius zonarius | 5 | 20 | 30 | 28.6071 |
| Red-capped parrot | Purpureicephalus spurius | 5 | 72 | 22 | 31.4403 |
| Superb parrot | Plytelis swainsonii | 20 | 27.5 | 22.5 | 23.9352 |
| Red-rumped parrot | Psephotus haematonotus | 20 | 72 | 22 | 32.6547 |
| Little lorikeet | Glossopsitta pusilla | 15 | 49.3 | 0.7 | 39.6925 |
| Varied lorikeet | Psitteuleles versicolor | 5 | 48 | 2 | 38.6567 |
| Rainbow lorikeet | Trichoglossus moluccanus | 20 | 46 | 4 | 30.3341 |
| Bourke’s parrot | Neopsephotus bourkii | 20 | 49.5 | 0.5 | 28.7778 |
| Common Name | Scientific Name | % Left | Nidopallium | Optic Tectum |
|---|---|---|---|---|
| Galah | Eolophus roseicapella | 89 | 0.3618 | 0.0314 |
| Yellow-tailed black cockatoo | Calyptorhynchus funereus | 100 | 0.3887 | 0.0196 |
| Cockatiel | Nymphicus hollandicus | 90 | 0.3571 | 0.0350 |
| Crimson rosella | Platycerus elegans | 23 | 0.3401 | 0.0429 |
| Budgerigar | Melopsittacus undulatus | 53 | 0.3210 | 0.0514 |
| Superb parrot | Plytelis swainsonii | 27.5 | 0.3200 | 0.0556 |
| Red-rumped parrot | Psephotus haematonotus | 72 | 0.3540 | 0.0387 |
| Rainbow lorikeet | Trichoglossus haematodus | 46 | 0.3370 | 0.0339 |
| Bourke’s parrot | Neopsephotus bourkii | 49.5 | 0.3586 | 0.0482 |
| Australian king parrot | Alisterus scapularis | 8.5 | 0.3098 | 0.0424 |
| Eclectus parrot | Eclectus roratus | 26 | 0.3534 | 0.0351 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaplan, G.; Rogers, L.J. Brain Size Associated with Foot Preferences in Australian Parrots. Symmetry 2021, 13, 867. https://doi.org/10.3390/sym13050867
Kaplan G, Rogers LJ. Brain Size Associated with Foot Preferences in Australian Parrots. Symmetry. 2021; 13(5):867. https://doi.org/10.3390/sym13050867
Chicago/Turabian StyleKaplan, Gisela, and Lesley J. Rogers. 2021. "Brain Size Associated with Foot Preferences in Australian Parrots" Symmetry 13, no. 5: 867. https://doi.org/10.3390/sym13050867
APA StyleKaplan, G., & Rogers, L. J. (2021). Brain Size Associated with Foot Preferences in Australian Parrots. Symmetry, 13(5), 867. https://doi.org/10.3390/sym13050867