
Individual Laterality in Ghost Crabs (Ocypode saratan) Influences Burrowing Behavior




Abstract
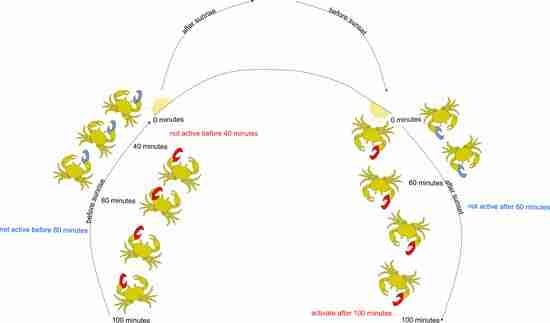
:
{kind=link}
{kind=link}
{kind=link}
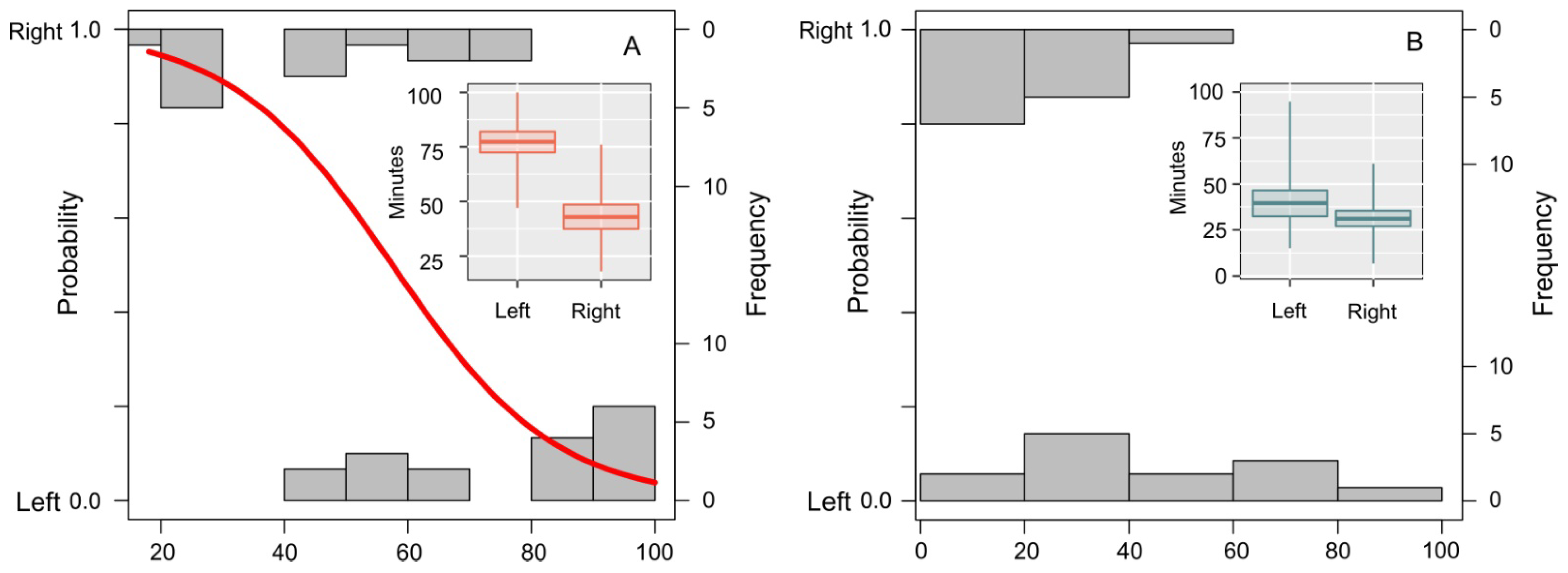{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Study Area
2.2. Field Study
2.3. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Lussanet, M.H.E. Opposite asymmetries of face and trunk and of kissing and hugging, as predicted by the axial twist hypothesis. PeerJ 2019, 7, e7096. [Google Scholar] [CrossRef] [Green Version]
- Anfora, G.; Frasnelli, E.; Maccagnani, B.; Rogers, L.J.; Vallortigara, G. Behavioural and electrophysiological lateralization in a social Apis mellifera but not in a non-social Osmia cornuta species of bee. Behav. Brain Res. 2010, 206, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left-right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D. Looking for the right mate: What do we really know on the courtship and mating of Luciliasericata (Meigen)? Acta Tropica 2019, 189, 145–153. [Google Scholar] [CrossRef]
- Bisazza, A.; Cantalupo, C.; Robins, A.; Rogers, L.J.; Vallortigara, G. Right-pawedness in toads. Nature 1996, 379, 408. [Google Scholar] [CrossRef]
- Bisazza, A.; Facchin, L.; Vallortigara, G. Heritability of lateralization in fish: Concordance of right–left asymmetry between parents and offspring. Neuropsychologia 2000, 38, 907–912. [Google Scholar] [CrossRef]
- Stancher, G.; Sovrano, V.A.; Vallortigara, G. Motor asymmetries in fishes, amphibians, and reptiles. Prog. Brain Res. 2018, 238, 33–56. [Google Scholar]
- Torres-Dowdall, J.; Rometsch, S.J.; Gastón, A.; Goyenola, G.; Meyer, A. Asymmetry in genitalia is in sync with lateralized mating behavior but not with the lateralization of other behaviors. Curr. Zool. 2020, 66, 71–81. [Google Scholar] [CrossRef]
- Hews, D.K.; Castellano, M.; Hara, E. Agression in females is also lateralized:left-eye bias during aggressive courtship rejection in lizards. Anim. Behav. 2004, 68, 1201–1207. [Google Scholar] [CrossRef]
- Yosef, R.; Gindi, C.; Sukenik, N. Footedness in Steppe Buzzard (Buteo vulpinus). Behav. Process. 2019, 158, 113–116. [Google Scholar] [CrossRef]
- Hugo, C.; Henry, L.; George, I.; Schedir, M.; Martine, H. Lateralization of social signal brain processing correlates with the degree of social integration in a songbird. Sci. Rep. 2020, 10, 14093. [Google Scholar]
- Galuret, S.; Lumineau, S.; Pouzol, D.; George, I. Mothering influences domestic chick’s laterality. Anim. Behav. 2020, 159, 69–79. [Google Scholar] [CrossRef]
- Zaynagutdinova, E.; Karenina, K.; Giljov, A. Lateralization in monogamous pairs: Wild geese prefer to keep their partner in the left hemifield except when disturbed. Curr. Zool. 2021, 67, 419–429. [Google Scholar] [CrossRef]
- Sinisalchi, M.; Padalino, B.; Lusito, R.; Quaranta, A. Is the left forelimb preference indicative of a stressful situation in horses? Behav. Proc. 2014, 107, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Karenina, K.; Giljov, A.; Ingram, J.; Rowntree, V.J.; Malashichev, Y. Lateralization of mother—Infant interactions in a diverse range of mammal species. Nat. Ecol. Evol. 2017, 1, 1–4. [Google Scholar] [CrossRef]
- Hauser, M.D.; Akre, K. Asymmetries in the timing of facial and vocal expressions by rhesus monkeys: Implications for hemispheric specialisation. Anim. Behav. 2001, 61, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Leliveld, L.M.C.; Scheumann, M.; Zimmermann, E. Effects of caller characteristics on auditory laterality in an early primate Microcebus murinus. PLoS ONE 2010, 5, e9031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govind, C.K. Asymmetry in lobster claws. Am. Sci. 1989, 77, 468–474. [Google Scholar]
- Hamilton, P.V.; Nishimoto, R.T.; Halusky, J.G. Cheliped laterality in Callinectes sapidus (Crustacea: Portunidae). Biol. Bull. Woods Hole 1976, 150, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Vermeij, G.J. Patterns in claw size: The geography of crushing. Systematic. Zool. 1977, 26, 138–151. [Google Scholar] [CrossRef]
- Rosenberg, M.S. The systematics and taxonomy of fiddler crabs: A phylogeny of the genus Uca. J. Crustacean Biol. 2001, 21, 839–869. [Google Scholar] [CrossRef] [Green Version]
- Crane, J. Fiddler Crabs of the World; (Ocypodidae: Genus Uca); Princeton University Press: Princetown, NJ, USA, 1975. [Google Scholar]
- Vannini, M. Researches on the coast of Somalia. The shore and dune of Sar Uanle. 27. Burrows and digging behavior in Ocypode and other crabs (Crustacean Brachyura). Monite. Zool. Ital. N. S. Ecol. Monog. 1980, 13, 11–44. [Google Scholar]
- Trott, T.J. The Prevalence of Left-handedness in the Painted Ghost Crab Ocypode gaudichaudii H. Milne Edwards & Lucas (Decapoda: Brachyura: Ocypodidae). Crustaceana 1987, 52, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Ng, P.K.L.; Tan, L.W.H. ‘Right handedness’ in the heterochelous calappoid and xanthoid crabs—Suggestion for a functional advantage. Crustaceana 1985, 49, 98–100. [Google Scholar]
- Morrell, L.J.; Backwell, P.R.Y.; Metcalfe, N.B. Fighting in fiddler crabs Uca mjoebergi: What determines duration? Anim. Behav. 2005, 70, 653–662. [Google Scholar] [CrossRef]
- Chan, B.K.K.; Chan, K.T.Y.; Leung, P.C.M. Burrow architecture of the Ghost Crab Ocypode ceratophtalma on a sandy shore in Hong Kong. Hydrobiologia 2006, 560, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Perez, D.M.; Rosenberg, M.S.; Pie, M.R. The evolution of waving displays in fiddler crabs (Uca spp., Crustacea: Ocypodidae). Biol. J. Linn. Soc. 2012, 106, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Perez, D.M.; Heatwole, S.J.; Morell, L.J.; Blackwell, P.R.Y. Handedness in fiddler crab fights. Anim. Behav. 2015, 110, 99–104. [Google Scholar] [CrossRef]
- Backwell, P.R.Y.; Matsumasa, M.; Double, M.; Roberts, A.; Murai, M.; Keogh, J.S.; Jennions, M.D. What are the consequences of being left-clawed in a predominantly right-clawed fiddler crab? Proc. R. Soc. B 2007, 274, 2723–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tina, F.W.; Jaroensutasinee, M.; Jaroensutasinee, K. Body size, resident status, handedness and claw originality in Uca rosea (Tweedie, 1937) male fights. Crustaceana 2016, 89, 1687–1700. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Marino, S.; Bellomo, S.; Vanadia, A.; Borzì, L.; Tibullo, D.; Di Stefano, A.; Lombardo, B.M. Abundance, distribution and ecology of the tufted ghost crab Ocypode cursor (Linnaeus, 1758) (Crustacea: Ocypodidae) from a recently colonized urban sandy beach, and new records from Sicily (central Mediterranean Sea). J. Sea Res. 2020, 156, 101832. [Google Scholar] [CrossRef]
- Von Linsenmair, K.E. Konstruktion und Signalfunktion de Sandpyramide der Reiterkrabbe Ocypode saratan Forsk. (Decapoda Brachyura Ocypodidae). Ph.D. Thesis, Aus dem Zoologischen Institut der Universität Frankfurt, Frankfurt, Germany, 1967; p. 54. [Google Scholar]
- Eshky, A.A. Aspects of the Ecology, Behaviour and Physiology of the Ghostcrab (Ocypode Saratan). Ph.D. Thesis, University of Glasgow, Glasgow, Scotland, UK, 1985; p. 451. [Google Scholar]
- Fishelson, L. Ecology and distribution of the benthic fauna in the shallow waters of the Red Sea. Mar. Biol. 1971, 10, 113–133. [Google Scholar] [CrossRef]
- Fishelson, L. Population ecology and biology of Dotilla sulcata (Crustacea, Ocypodidae) typical for sandy beaches of the Red Sea. In Sandy Beaches as Ecosystems; McLachlan, A., Erasmus, T., Eds.; Dr W. Junk Publishers: The Hague, The Netherlands, 1983; pp. 643–654. [Google Scholar]
- Karleskint, G.; Turner, R.K.; Small, J. Intertidal communities. In Introduction to Marine Biology, 3rd ed.; Cengage Learning: Boston, MA, USA, 2009. [Google Scholar]
- Wolcott, T.G. Ecological role of ghost crabs, Ocypode quadrata (Fabricius) on an ocean beach: Scavengers or predators? J. Exp. Mar. Biol. Ecol. 1978, 31, 67–82. [Google Scholar] [CrossRef]
- Trott, T.J. Gustatory responses of ghost crab Ocypode quadrata to seawater extracts and chemical fractions of natural stimuli. J. Chem. Ecol. 1999, 25, 375–388. [Google Scholar] [CrossRef]
- Marco, A.; Da Graça, J.; García-Cerdá, R.; Abella, E.; Freitas, R. Patterns and intensity of ghost crab predation on the nests of an important endangered loggerhead turtle population. J. Exp. Mar. Biol. Ecol. 2015, 468, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Deidun, A.; Crocetta, F.; Sciberras, A.; Sciberras, J.; Insacco, G.; Zava, B. The protected taxon Ocypode cursor (Linnaeus, 1758) (Crustacea: Decapoda: Ocypodidae)—Documenting its well-established presence in the central Mediterranean. Eur. Zool. J. 2017, 84, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Nye, P.A. Burrowing and burying by the crab Macropthalmus hirtipes. N. Z. J. Mar. Freshw. Res. 1974, 8, 243–254. [Google Scholar] [CrossRef]
- Seike, K.; Nara, M. Occurrence of bioglyphs on Ocypode crab burrows in a modern sandy beach and its palaeoenvironmental implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 252, 458–463. [Google Scholar] [CrossRef]
- Haque, H.; Choudhury, A. Ecology and behavior of the ghost crab Ocypode macrocerca Edwards 1834 occuring in the sandy beaches of Sagar Island, Sunderbans. Int. J. Eng. Sci. Invent. 2014, 3, 38–43. [Google Scholar]
- Pratt, A.E.; McLain, D.K.; Lathrop, G.R. The assessment game in sand fiddler crab contests for breeding burrows. Anim. Behav. 2003, 65, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; W. H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- Jennions, M.D.; Backwell, P.R.Y. Residency and size affect fight duration and outcome in the fiddler crab Uca annulipes. Biol. J. Linn. Soc. 1996, 57, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Seike, K.; Nara, M. Burrow morphologies of the ghost crabs Ocypode ceratophtalma and O. sinensis in foreshore, backshore, and dune environments of a sandy beach in Japan. J. Geol. Soc. Jpn. 2008, 114, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.L.; Soares-Gomes, A.; Zalmon, I.R. Burrow occupation rates and spatial distribution within habitat of the ghost crab Ocypode quadrata (Fabricius, 1787): Implications for impact assessments. Reg. Stud. Mar. Sci. 2021, 44, 101699. [Google Scholar] [CrossRef]
- De Souza, G.N.; Oliviera, C.A.G.; Tardem, A.S.; Soares-Gomes, A. Counting and measuring ghost crab burrows as a way to assess the environmental quality of beaches. Ocean Coast. Manag. 2017, 140, 1–10. [Google Scholar] [CrossRef]
- Schober, U.M.; Christy, J.H. Sand disposal of the painted ghost crab Ocypode gaudichaudii (Decapoda: Ocypodidae): A possible role in courtship. Mar. Biol. 1993, 116, 53–60. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yosef, R.; Daraby, M.; Semionovikh, A.; Kosicki, J.Z. Individual Laterality in Ghost Crabs (Ocypode saratan) Influences Burrowing Behavior. Symmetry 2021, 13, 1512. https://doi.org/10.3390/sym13081512
Yosef R, Daraby M, Semionovikh A, Kosicki JZ. Individual Laterality in Ghost Crabs (Ocypode saratan) Influences Burrowing Behavior. Symmetry. 2021; 13(8):1512. https://doi.org/10.3390/sym13081512
Chicago/Turabian StyleYosef, Reuven, Michal Daraby, Alexei Semionovikh, and Jakub Z. Kosicki. 2021. "Individual Laterality in Ghost Crabs (Ocypode saratan) Influences Burrowing Behavior" Symmetry 13, no. 8: 1512. https://doi.org/10.3390/sym13081512
APA StyleYosef, R., Daraby, M., Semionovikh, A., & Kosicki, J. Z. (2021). Individual Laterality in Ghost Crabs (Ocypode saratan) Influences Burrowing Behavior. Symmetry, 13(8), 1512. https://doi.org/10.3390/sym13081512