
Spatial Integration of Cellular Shapes in Green Microalgae with Complex Morphology, the Genus Micrasterias (Desmidiales, Zygnematophyceae)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation and Data Acquisition
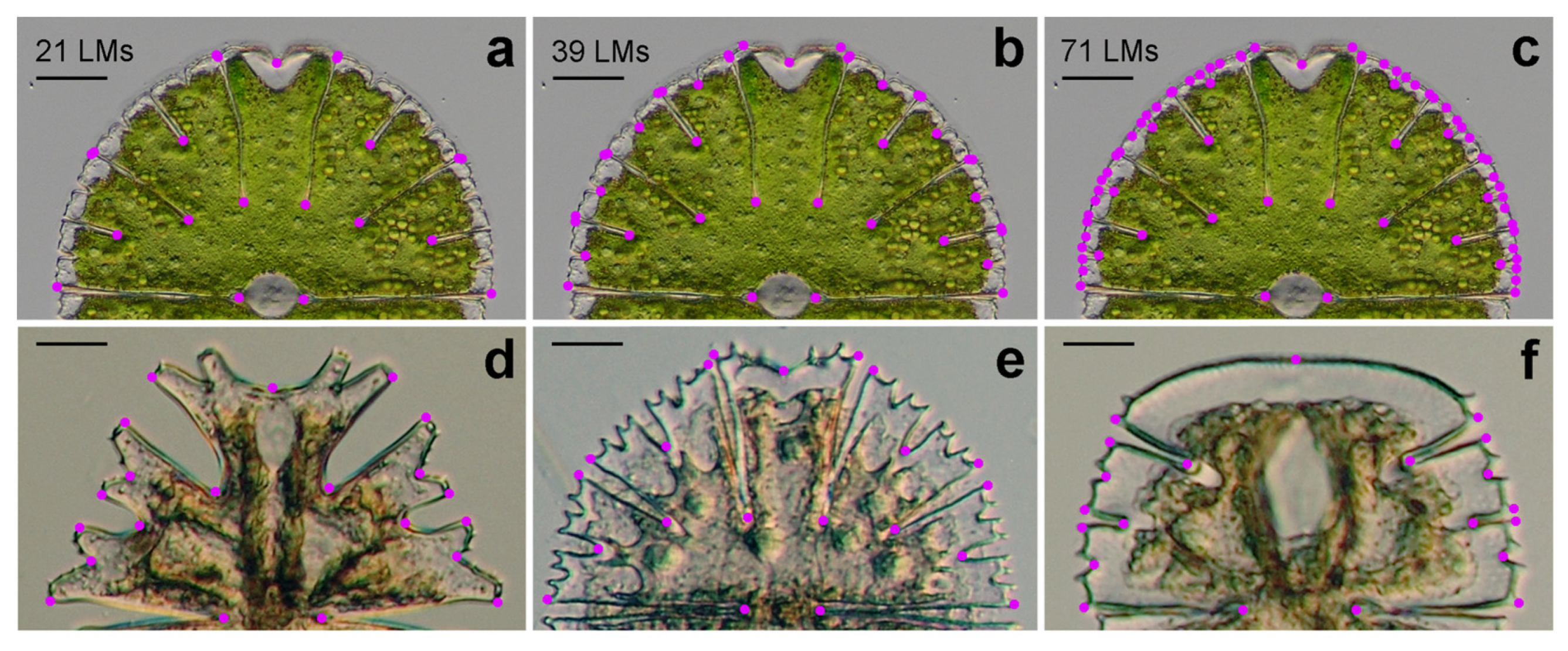
2.2. Geometric Morphometrics
3. Results
3.1. Patterns of PWs—An Example of M. thomasiana Reared at 25 °C
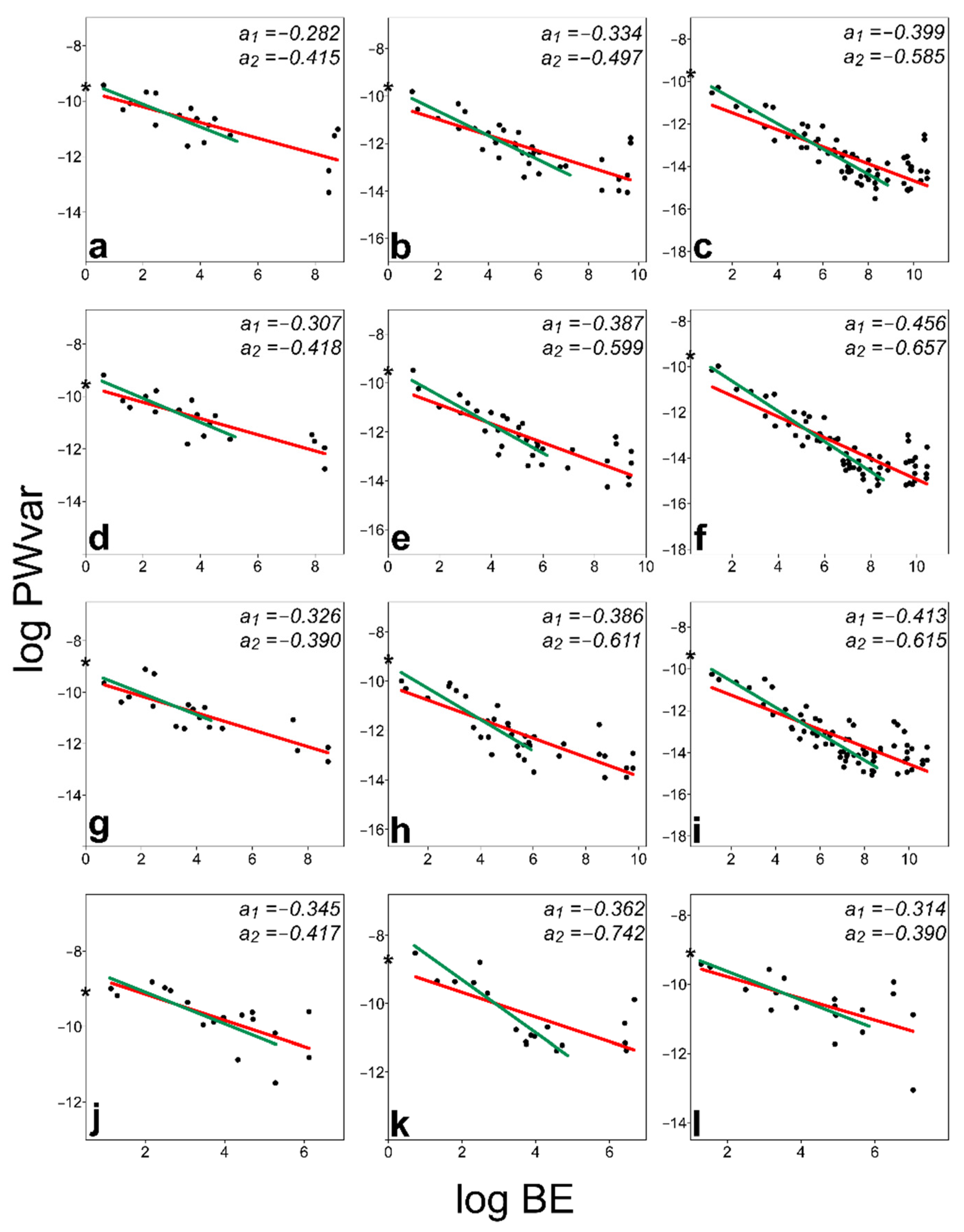
3.2. BE–PWv Plots
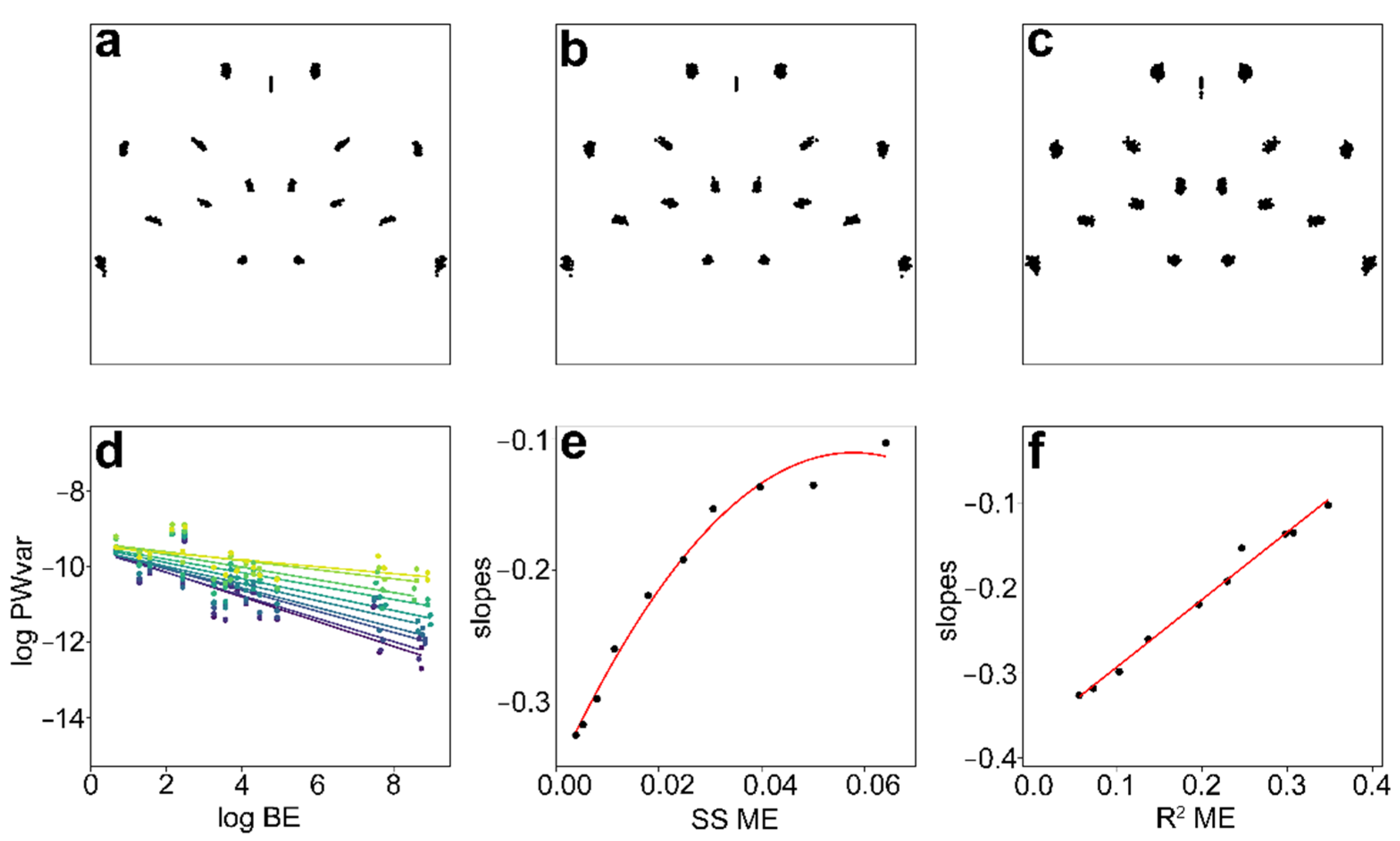
3.3. ME Assessment
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olson, E.C.; Miller, R.L. Morphological Integration; University of Chicago Press: Chicago, IL, USA, 1958; p. 376. [Google Scholar]
- Parés-Casanova, P.M.; Rissech, C.; Davis, S.; Lloveras, L. Morphological integration on the calcaneum of domestic sheep (Ovis aries Linnaeus, 1758)–a geometric morphometric study. Appl. Sci. 2023, 13, 4177. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Studying morphological integration and modularity at multiple levels: Concepts and analysis. Phil. Trans. R. Soc. B 2014, 369, 20130249. [Google Scholar] [CrossRef] [PubMed]
- Conaway, M.A.; Adams, D.C. An effect size for comparing the strength of morphological integration across studies. Evolution 2022, 76, 2244–2259. [Google Scholar] [CrossRef] [PubMed]
- Benítez, H.A.; Püschel, T.A.; Suazo, M.J. Drosophila wing integration and modularity: A multi-level approach to understand the history of morphological structures. Biology 2022, 11, 567. [Google Scholar] [CrossRef]
- Harold, F.M. To shape a cell: An inquiry into the causes of morphogenesis of microorganisms. Microb. Rev. 1990, 54, 381–431. [Google Scholar] [CrossRef]
- Lütz-Meindl, U. Micrasterias as a model system in plant cell biology. Front. Plant Sci. 2016, 7, 999. [Google Scholar] [CrossRef]
- Weiss, D.; Lütz, C.; Lütz-Meindl, U. Photosynthesis and heat response of the green alga Micrasterias denticulata (Desmidiaceae). Zeitsch. Naturf. 1999, 54, 508–516. [Google Scholar] [CrossRef]
- Song, X.; Kong, F.; Liu, B.F.; Song, Q.; Ren, N.Q.; Ren, H.Y. Thallium-mediated NO signalling induced lipid accumulation in microalgae and its role in heavy metal bioremediation. Water Res. 2023, 239, 120027. [Google Scholar] [CrossRef]
- Brook, A.J. The Biology of Desmids; Blackwell Science: Oxford, UK, 1981; p. 276. [Google Scholar]
- Felhofer, M.; Mayr, K.; Lütz-Meindl, U.; Gierlinger, N. Raman imaging of Micrasterias: New insights into shape formation. Protoplasma 2021, 258, 1323–1334. [Google Scholar] [CrossRef]
- Neustupa, J. Asymmetry and integration of cellular morphology in Micrasterias compereana. BMC Evol. Biol. 2017, 17, 1. [Google Scholar] [CrossRef]
- Harrison, L.G. The Shaping of Life: The Generation of Biological Pattern; Cambridge University Press: Cambridge, UK, 2011; p. 247. [Google Scholar]
- Neustupa, J. Static allometry of unicellular green algae: Scaling of cellular surface area and volume in the genus Micrasterias (Desmidiales). J. Evol. Biol. 2016, 29, 292–305. [Google Scholar] [CrossRef]
- Mandelbrot, B.B. The Fractal Geometry of Nature; W.H. Freeman and Company: New York, NY, USA, 1982; p. 497. [Google Scholar]
- Bookstein, F.L. Integration, disintegration, and self-similarity: Characterizing the scales of shape variation in landmark data. Evol. Biol. 2015, 42, 395–426. [Google Scholar] [CrossRef]
- Bookstein, F.L. A Course in Morphometrics for Biologists; Cambridge University Press: Cambridge, UK, 2018; p. 527. [Google Scholar]
- Windhager, S.; Bookstein, F.L.; Millesi, E.; Wallner, B.; Schaefer, K. Patterns of correlation of facial shape with physiological measurements are more integrated than patterns of correlation with ratings. Sci. Rep. 2017, 7, 45340. [Google Scholar] [CrossRef]
- Mitteroecker, P.; Bartsch, S.; Erkinger, C.; Grunstra, N.D.S.; Le Maître, A.; Bookstein, F.L. Morphometric variation at different spatial scales: Coordination and compensation in the emergence of organismal form. Syst. Biol. 2020, 69, 913–926. [Google Scholar] [CrossRef]
- Cardini, A. Integration and modularity in Procrustes shape data: Is there a risk of spurious results? Evol. Biol. 2019, 46, 90–105. [Google Scholar] [CrossRef]
- McNally, J.G. Scale-invariant pattern in the alga Micrasterias. J Biol. Phys. 1990, 17, 235–243. [Google Scholar] [CrossRef]
- Škaloud, P.; Nemjová, K.; Veselá, J.; Černá, K.; Neustupa, J. A multilocus phylogeny of the desmid genus Micrasterias (Streptophyta): Evidence for the accelerated rate of morphological evolution in protists. Mol. Phyl. Evol. 2011, 61, 933–943. [Google Scholar] [CrossRef]
- Poulíčková, A.; Mazalová, P.; Vašut, R.J.; Šarhanová, P.; Neustupa, J. DNA content variation and its significance in the evolution of the genus Micrasterias (Desmidiales, Streptophyta). PLoS ONE 2014, 9, e86247. [Google Scholar] [CrossRef]
- Araujo, C.B.D.; Bicudo, C.E.D.M.; Silva, T.G.D.; Trumhová, K.; Škaloud, P. Hidden generic diversity in desmids: Description of Pseudomicrasterias gen. nov. (Desmidiaceae, Zygnematophyceae). Phycologia 2022, 61, 227–240. [Google Scholar] [CrossRef]
- Andersen, R.A. Algal Culturing Techniques; Academic Press: Amsterdam, The Netherlands, 2005; p. 578. [Google Scholar]
- Rohlf, F.J. The Tps series of software. Hystrix 2015, 26, 9–12. [Google Scholar]
- Klingenberg, C.P. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Bookstein, F.L. Reflections on a biometrics of organismal form. Biol. Theory 2019, 14, 177–211. [Google Scholar] [CrossRef]
- Muggeo, V.M. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. A Language and Environment for Statistical Computing. Available online: https://www.r-project.org (accessed on 12 August 2023).
- Adams, D.C.; Otárola-Castillo, E. Geomorph: An R package for the collection and analysis of geometric morphometric shape data. Meth. Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- Dryden, I.L. Shapes Packzage. Available online: https://cran.r-project.org/web/packages/shapes/index.html (accessed on 12 August 2023).
- Muggeo, V.M. Segmented: An R Package to Fit Regression Models with Broken-Line Relationships. Available online: https://cran.r-project.org/doc/Rnews/ (accessed on 12 August 2023).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeont. Electr. 2001, 4, 1–9. [Google Scholar]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D. Geometric Morphometrics for Biologists. A Primer; Elsevier: Amsterdam, The Netherlands, 2012; p. 443. [Google Scholar]
- Piras, P.; Torromeo, C.; Evangelista, A.; Gabriele, S.; Esposito, G.; Nardinocchi, P.; Teresi, L.; Madeo, A.; Schiariti, M.; Varano, V.; et al. Homeostatic left heart integration and disintegration links atrio-ventricular covariation’s dyshomeostasis in hypertrophic cardiomyopathy. Sci. Rep. 2017, 7, 6257. [Google Scholar] [CrossRef]
- Grunstra, N.D.S.; Bartsch, S.J.; Le Maître, A.; Mitteroecker, P. Detecting phylogenetic signal and adaptation in papionin cranial shape by decomposing variation at different spatial scales. Syst. Biol. 2021, 70, 694–706. [Google Scholar] [CrossRef]
- Sansalone, G.; Paolo, C.; Riccardo, C.; Stephen, W.; Silvia, C.; Pasquale, R. Trapped in the morphospace: The relationship between morphological integration and functional performance. Evolution 2022, 76, 2020–2031. [Google Scholar] [CrossRef]
- Waris, H.; Kallio, P. Morphogenesis in Micrasterias. Adv. Morphog. 1964, 4, 45–80. [Google Scholar]
- Kallio, P.; Lehtonen, J. Nuclear control of morphogenesis in Micrasterias. In Cytomorphogenesis in Plants; Kiermayer, O., Ed.; Springer: Vienna, Austria, 1981; pp. 191–213. [Google Scholar]
- Lacalli, T.C. Morphogenesis in Micrasterias. II. Patterns of morphogenesis. J. Embryol. Exp. Morph. 1975, 33, 117–126. [Google Scholar] [CrossRef]
- Lacalli, T.C. Morphogenesis in Micrasterias. III. The morphogenetic template. Protoplasma 1976, 88, 133–146. [Google Scholar] [CrossRef]
- Lacalli, T.C.; Harrison, L.G. Turing’s model and branching tip growth: Relation of time and spatial scales in morphogenesis, with application to Micrasterias. Can. J. Bot. 1987, 65, 1308–1319. [Google Scholar] [CrossRef]
- Holloway, D.M.; Harrison, L.G. Algal morphogenesis: Modelling interspecific variation in Micrasterias with reaction–diffusion patterned catalysis of cell surface growth. Phil. Trans. R. Soc. B 1999, 354, 417–433. [Google Scholar] [CrossRef]
- Holloway, D.M.; Harrison, L.G. Pattern selection in plants: Coupling chemical dynamics to surface growth in three dimensions. Ann. Bot 2008, 101, 361–374. [Google Scholar] [CrossRef]
- Holloway, D.M. The chemical kinetics of shape determination in plants. In Chemical Kinetics; Patel, V., Ed.; InTech Press: Rijeka, Croatia, 2012; pp. 203–226. [Google Scholar]
- Lenarczyk, J. Evolution of morphological variability and modularity in single cells of algal colonies: A case study Pseudopediastrum (Hydrodictyaceae, Sphaeropleales, Chlorophyceae). Phycologia 2019, 58, 180–191. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Slope a1 | SE | 95% CI for a1 | Slope a1 (ME Adjusted) | Slope a2 | SE | 95% CI for a2 | Slope a2 (ME Adjusted) |
|---|---|---|---|---|---|---|---|---|
| MiTh, 17 °C 71 LMs | −0.399 | 0.038 | [−0.497, −0.320] | −0.457 | −0.585 | 0.036 | [−0.648, −0.515] | −0.671 |
| MiTh, 17 °C 39 LMs | −0.334 | 0.046 | [−0.454, −0.221] | −0.422 | −0.497 | 0.057 | [−0.590, −0.392] | −0.614 |
| MiTh, 17 °C 21 LMs | −0.282 | 0.059 | [−0.419, −0.115] | −0.358 | −0.415 | 0.056 | [−0.547, −0.326] | −0.490 |
| MiTh, 21 °C 71 LMs | −0.456 | 0.036 | [−0.544, −0.378] | −0.485 | −0.657 | 0.036 | [−0.722, −0.593] | −0.681 |
| MiTh, 21 °C 39 LMs | −0.387 | 0.046 | [−0.494, −0.284] | −0.434 | −0.599 | 0.068 | [−0.716, −0.489] | −0.673 |
| MiTh, 21 °C 21 LMs | −0.307 | 0.050 | [−0.397, −0.202] | −0.371 | −0.418 | 0.104 | [−0.604, −0.256] | −0.486 |
| MiTh, 25 °C 71 LMs | −0.413 | 0.039 | [−0.502, −0.338] | −0.437 | −0.615 | 0.042 | [−0.688, −0.539] | −0.636 |
| MiTh, 25 °C 39 LMs | −0.386 | 0.047 | [−0.472, −0.298] | −0.416 | −0.611 | 0.089 | [−0.726, −0.401] | −0.672 |
| MiTh, 25 °C 21 LMs | −0.326 | 0.055 | [−0.428, −0.219] | −0.369 | −0.390 | 0.140 | [−0.576, −0.061] | −0.462 |
| MiAm 21 LMs | −0.345 | 0.085 | [−0.496, −0.137] | −0.360 | −0.417 | 0.094 | [−0.589, −0.163] | −0.435 |
| MiPa 21 LMs | −0.362 | 0.095 | [−0.567, −0.117] | −0.373 | −0.742 | 0.092 | [−0.867, −0.518] | −0.760 |
| MiTr 21 LMs | −0.314 | 0.096 | [−0.527, −0.092] | −0.322 | −0.390 | 0.084 | [−0.532, −0.251] | −0.406 |
| Dataset | SS (Individual) | R2 (Individual) | SS (Measurement Error) | R2 (Measurement Error) | SS (Total) |
|---|---|---|---|---|---|
| original data | 0.0653 | 0.9448 | 0.0038 | 0.0552 | 0.0691 |
| random ME_+1 | 0.0674 | 0.9280 | 0.0052 | 0.0720 | 0.0727 |
| random ME_+2 | 0.0694 | 0.8977 | 0.0079 | 0.1023 | 0.0773 |
| random ME_+3 | 0.0719 | 0.8636 | 0.0114 | 0.1364 | 0.0833 |
| random ME_+4 | 0.0735 | 0.8041 | 0.0179 | 0.1959 | 0.0914 |
| random ME_+5 | 0.0832 | 0.7709 | 0.0247 | 0.2291 | 0.1079 |
| random ME_+6 | 0.0936 | 0.7539 | 0.0306 | 0.2462 | 0.1242 |
| random ME_+7 | 0.0937 | 0.7025 | 0.0397 | 0.2975 | 0.1334 |
| random ME_+8 | 0.1132 | 0.6933 | 0.0501 | 0.3067 | 0.1633 |
| random ME_+9 | 0.1204 | 0.6522 | 0.0642 | 0.3478 | 0.1845 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neustupa, J.; Woodard, K. Spatial Integration of Cellular Shapes in Green Microalgae with Complex Morphology, the Genus Micrasterias (Desmidiales, Zygnematophyceae). Symmetry 2023, 15, 1700. https://doi.org/10.3390/sym15091700
Neustupa J, Woodard K. Spatial Integration of Cellular Shapes in Green Microalgae with Complex Morphology, the Genus Micrasterias (Desmidiales, Zygnematophyceae). Symmetry. 2023; 15(9):1700. https://doi.org/10.3390/sym15091700
Chicago/Turabian StyleNeustupa, Jiri, and Katerina Woodard. 2023. "Spatial Integration of Cellular Shapes in Green Microalgae with Complex Morphology, the Genus Micrasterias (Desmidiales, Zygnematophyceae)" Symmetry 15, no. 9: 1700. https://doi.org/10.3390/sym15091700
APA StyleNeustupa, J., & Woodard, K. (2023). Spatial Integration of Cellular Shapes in Green Microalgae with Complex Morphology, the Genus Micrasterias (Desmidiales, Zygnematophyceae). Symmetry, 15(9), 1700. https://doi.org/10.3390/sym15091700