
Propagation Dynamics of an Epidemic Model with Heterogeneous Control Strategies on Complex Networks


Abstract
:1. Introduction
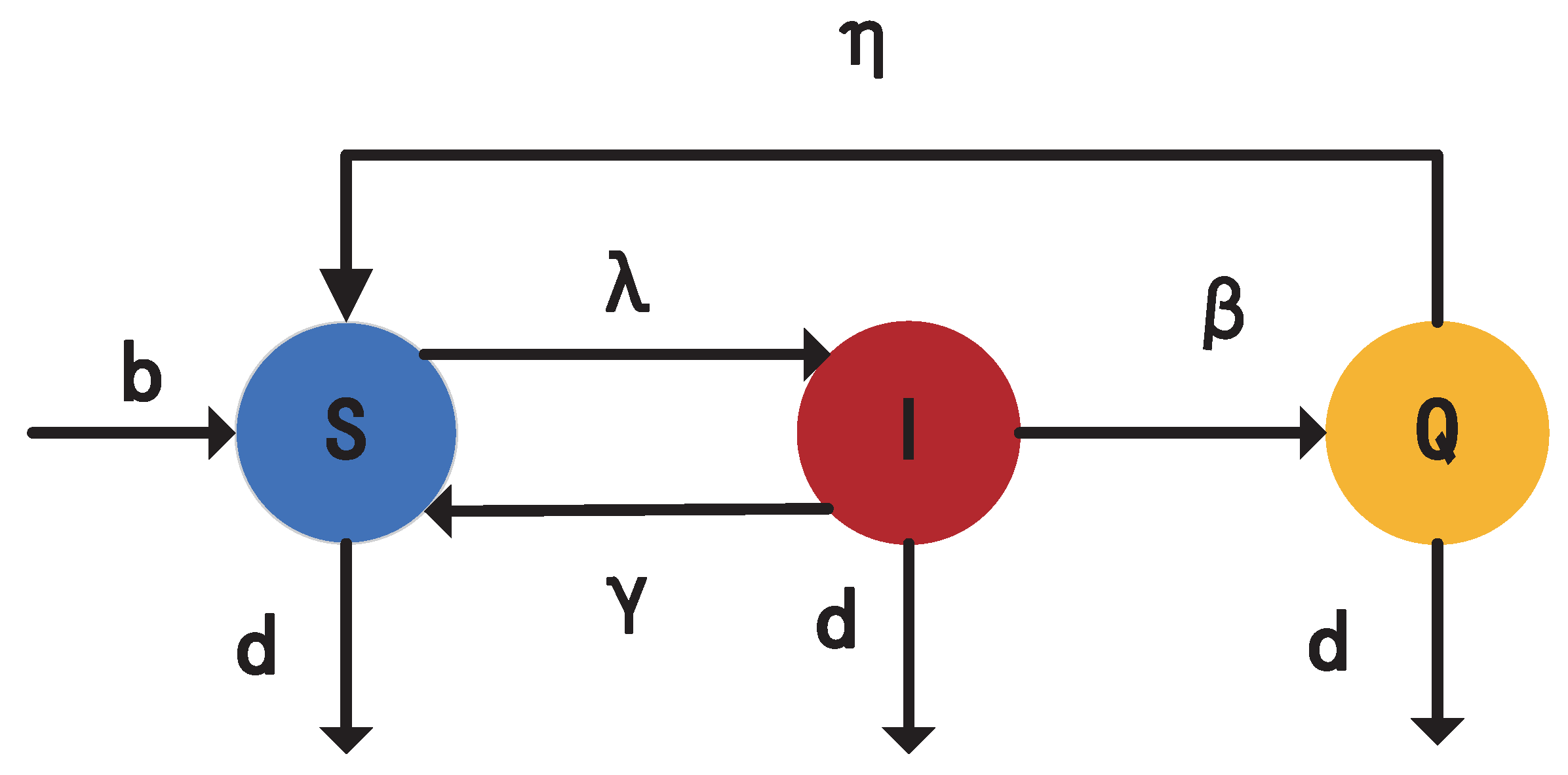
2. Description and Formation of Epidemic Models
- (1)
- Birth and death: Each vacant node i in the network randomly selects a neighbor. If the neighbor is a vacant node, the state of i remains unchanged. If the neighbor is a nonvacant node, the vacant node i will be activated to generate a new susceptible node at the birth rate b. Each nonvacant node becomes a vacant node at a natural death rate d per unit time. We assume that each nonvacant node has the same birth contact ability A (where ) due to physiological constraints.
- (2)
- Immunization and quarantine (): At each time step, susceptible individuals with degree k are immunized at the immune rate . The infected nodes with degree k will be quarantined at rate . The quarantined individuals will recover to a susceptible node at rate . Nodes with the same degree have identical quarantine and immunization strategies, while those with different degrees have different strategies.
- (3)
- SIS epidemic framework: Infection : At the initial moment, some nodes are randomly selected as infected nodes. At each time step, the possibility that each infected node i will connect to its neighboring nodes is , where represents the infectivity of infected nodes with degree k, and [38,39,40], = A [41], = [42], = [43]. If an infected node i interacts with a susceptible node j along a connecting edge, node j has a possibility of being infected by i at a transmission rate . For a node with degree k, the overall transmission rate is .
3. Equilibria and Basic Reproduction Number
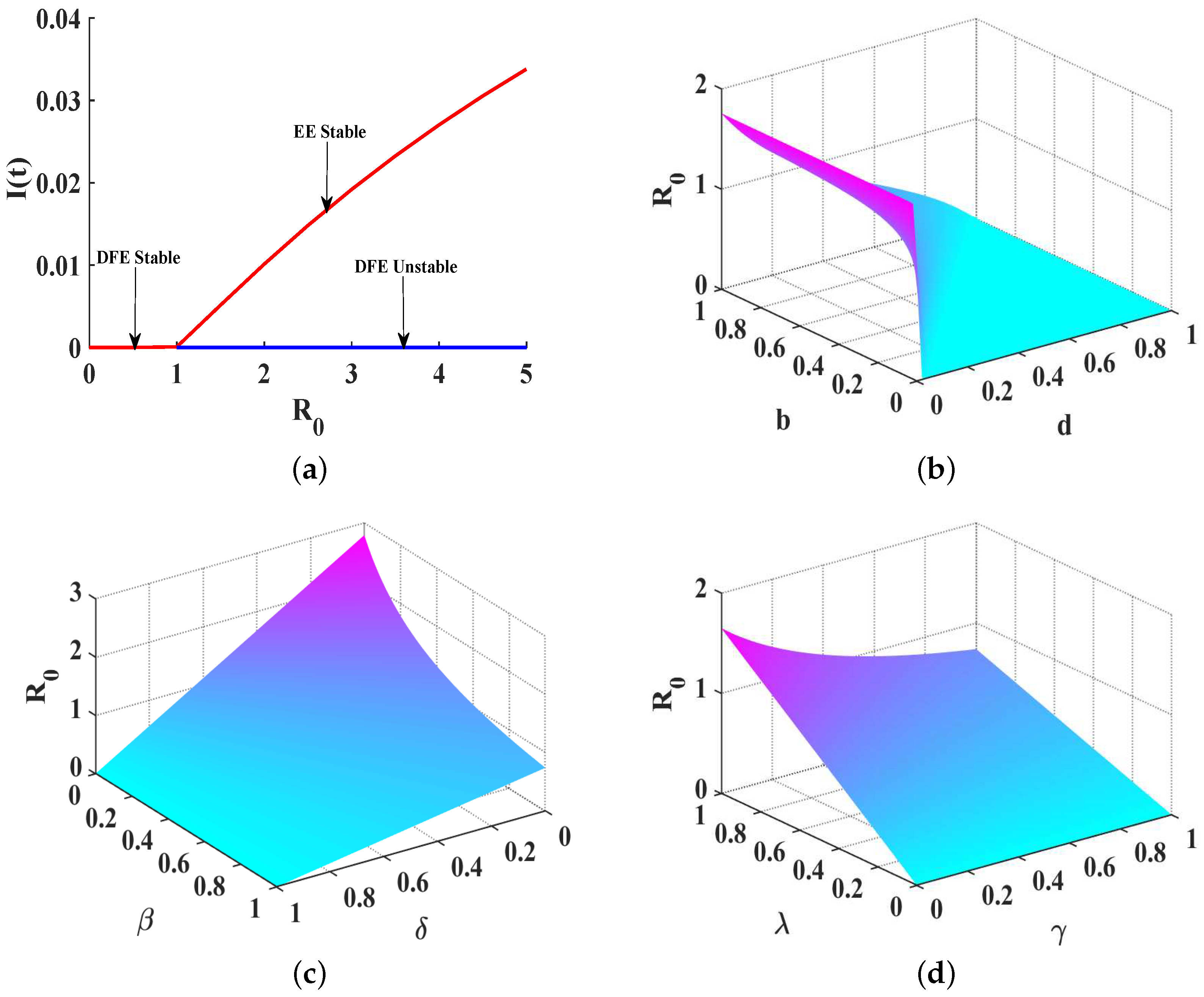
4. Stability Analysis for SIQS Model
4.1. Stability Analysis of Disease-Free Equilibrium
- Case 1
- If then we obtain
- Case 2
- If , since the sum of all eigenvalues is equal to the trace of the matrix, when , , and , .
4.2. Global Stability of Endemic Equilibrium
5. The Optimal Control for the SIQS Model
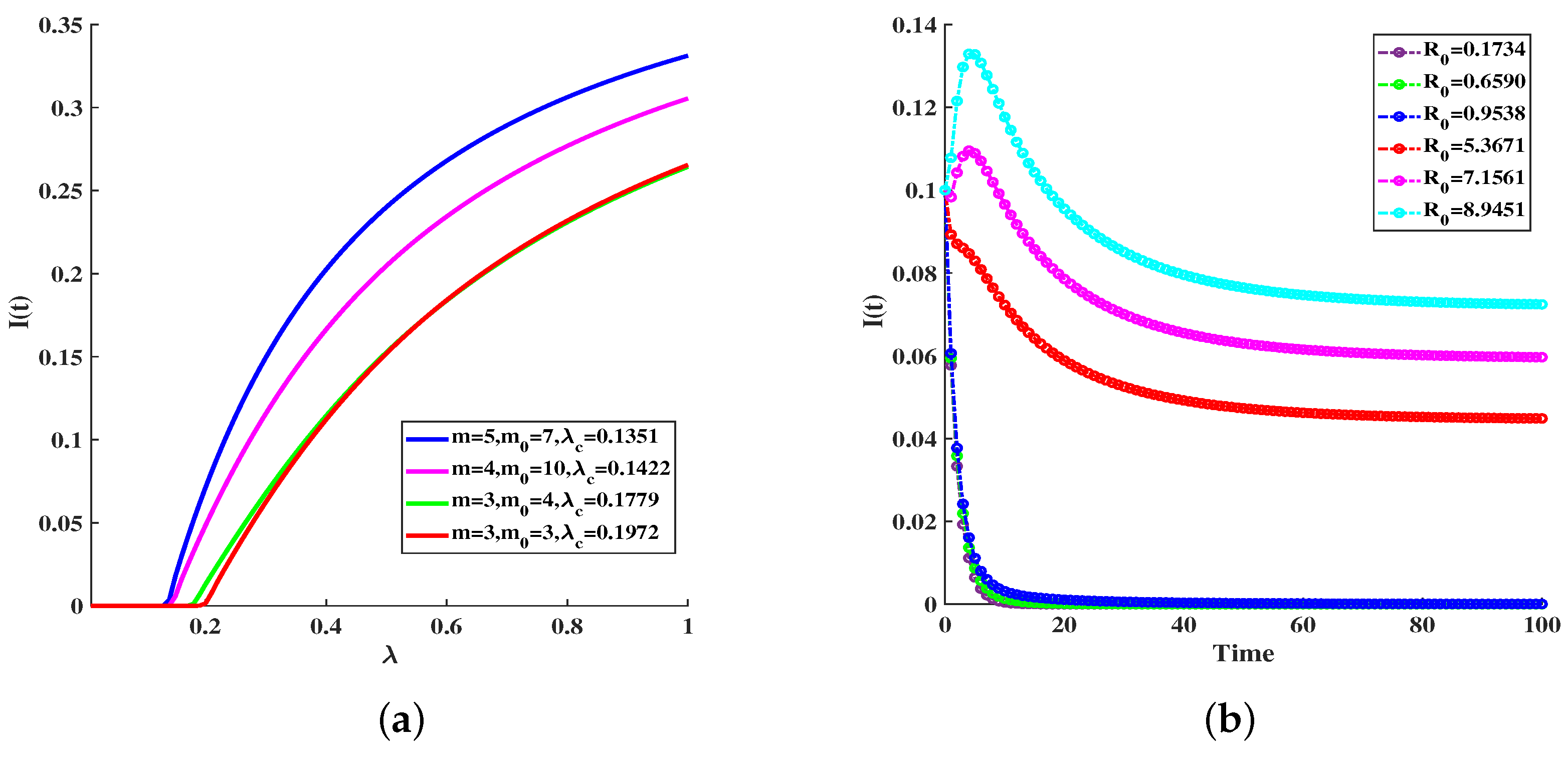
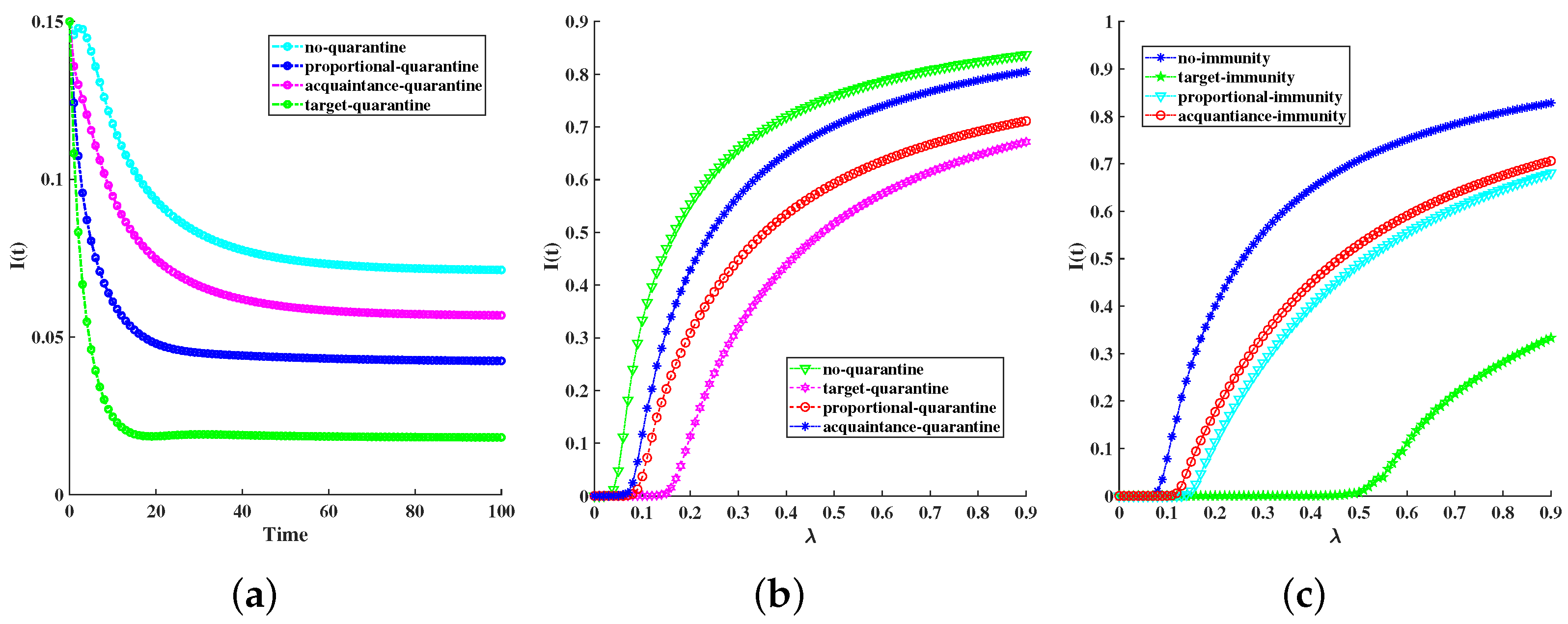
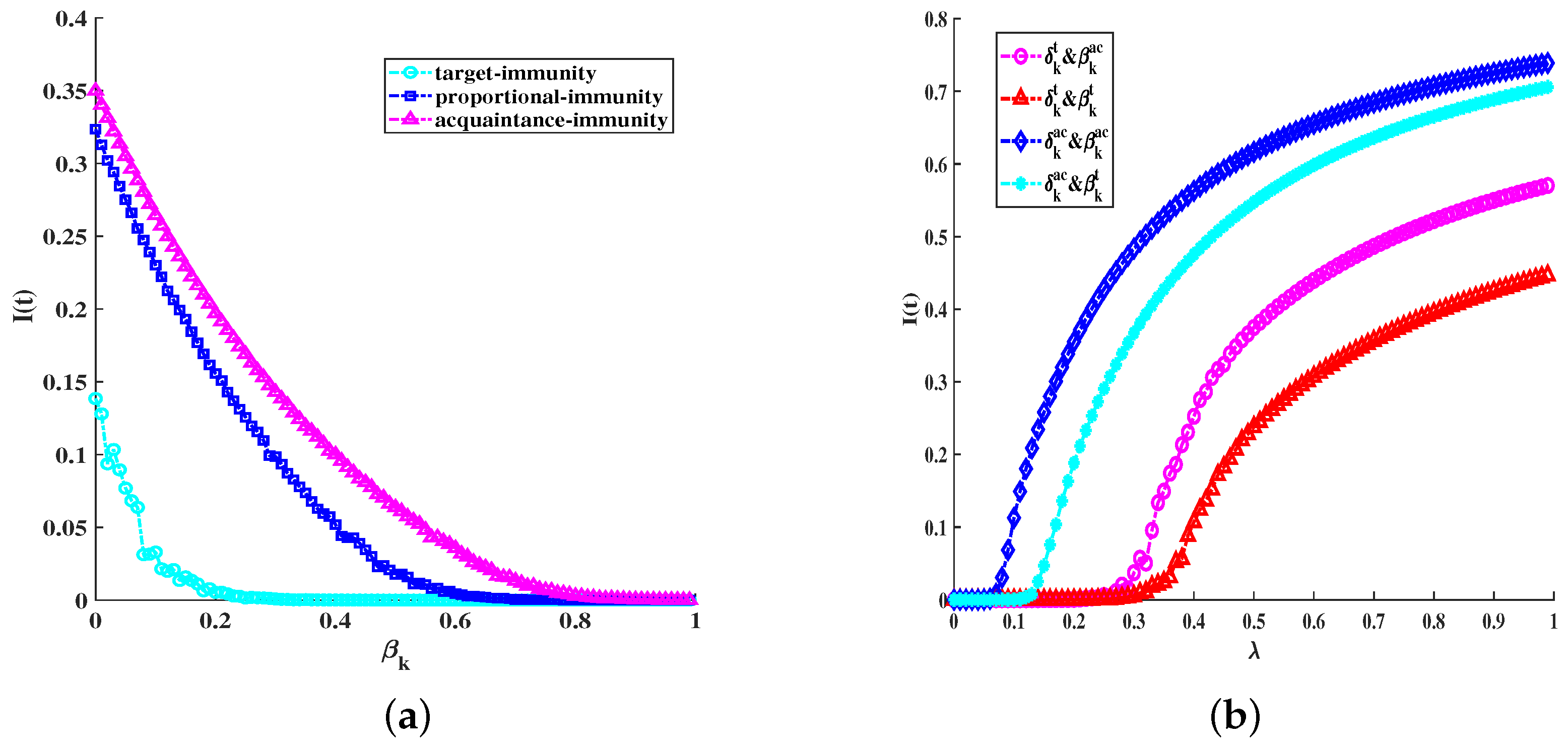
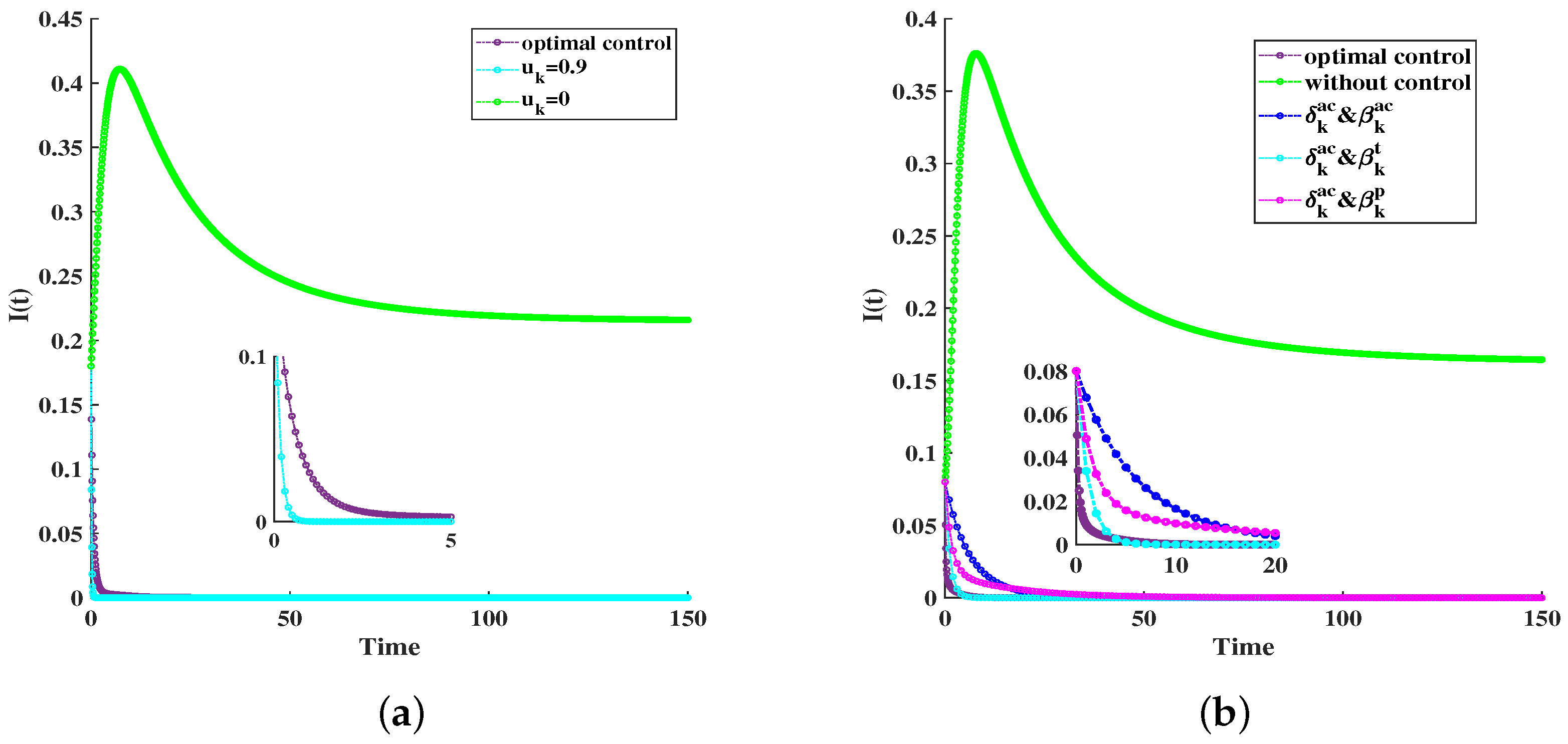
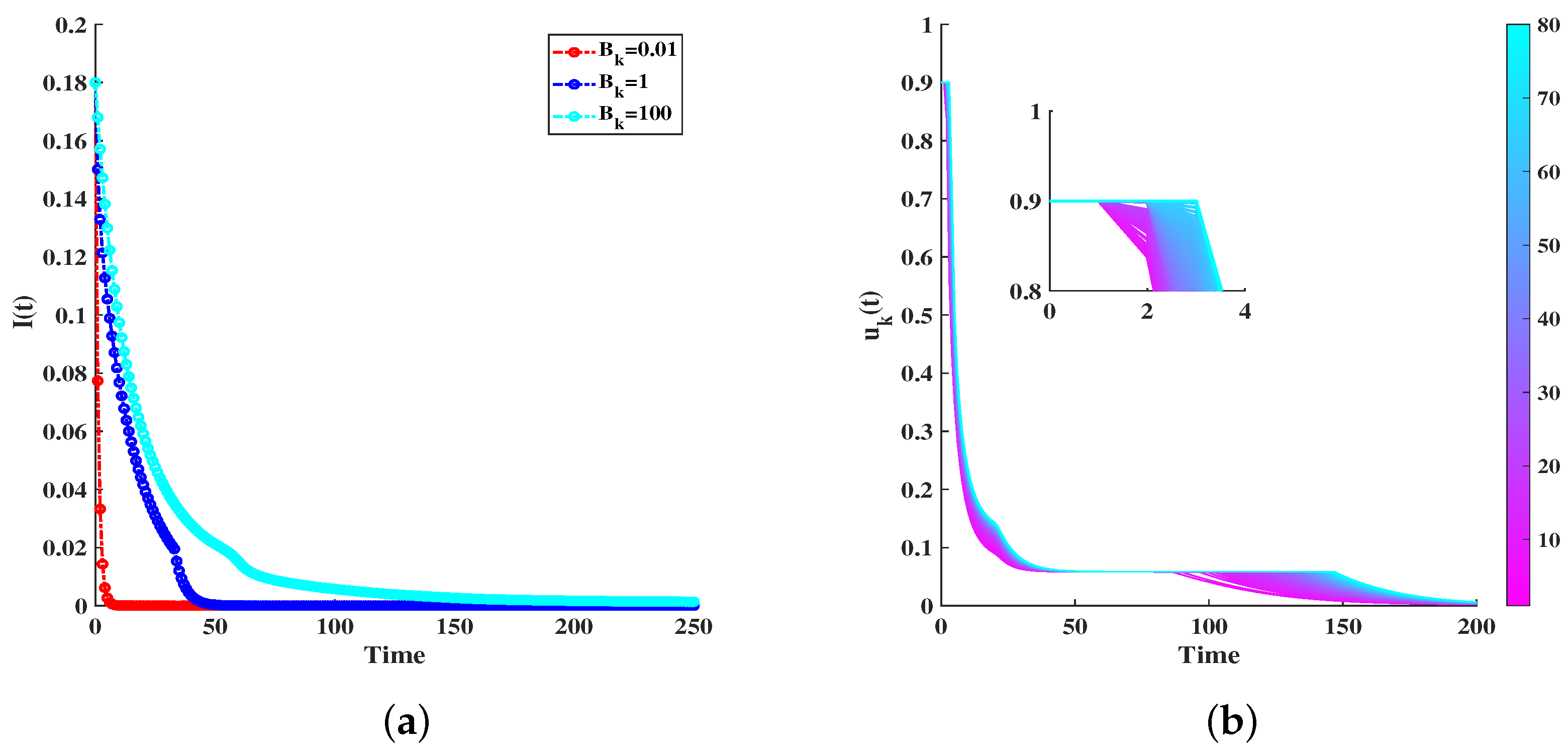
6. Simulations
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, B.; Shang, K.K.; Michael, S.; Chao, N.P. Information overload: How hot topics distract from news—COVID-19 spread in the US. Natl. Sci. Open 2023, 2, 20220051. [Google Scholar] [CrossRef]
- Jose, S.A.; Raja, R.; Omede, B.; Agarwal, R.P.; Alzabut, J.; Cao, J.; Balas, V. Mathematical modeling on co-infection: Transmission dynamics of Zika virus and Dengue fever. Nonlinear Dyn. 2023, 111, 4879–4914. [Google Scholar] [CrossRef]
- Peter, O.J.; Afolabi, O.A.; Victor, A.A.; Akpan, C.E.; Oguntolu, F.A. Mathematical model for the control of measles. J. Appl. Sci. Environ. Manag. 2018, 22, 571–576. [Google Scholar] [CrossRef]
- Fraser, C.; Donnelly, C.A.; Cauchemez, S.; Hanage, W.P.; Van Kerkhove, M.D.; Hollingsworth, T.D.; Griffin, J.; Baggaley, R.F.; Jenkins, H.E.; Lyons, E.J.; et al. Pandemic potential of a strain of influenza A (H1N1): Early findings. Science 2009, 324, 1557–1561. [Google Scholar] [CrossRef]
- Lipsitch, M.; Cohen, T.; Cooper, B.; Robins, J.M.; Ma, S.; James, L.; Gopalakrishna, G.; Chew, S.K.; Tan, C.C.; Samore, M.H.; et al. Transmission dynamics and control of severe acute respiratory syndrome. Science 2003, 300, 1966–1970. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.M.; Li, Z.Q.; Song, X.Y. Global analysis of an epidemic model with vaccination. J. Appl. Math. Comput. 2018, 57, 605–628. [Google Scholar] [CrossRef] [PubMed]
- Kermack, W.O.; McKendrick, A.G. A contribution to the mathematical theory of epidemics. Proc. R. Soc. Lond. Ser. Contain. Pap. Math. Phys. Character 1927, 115, 700–721. [Google Scholar]
- Kermack, W.O.; McKendrick, A.G. Contributions to the mathematical theory of epidemics. II.The problem of endemicity. Proc. R. Soc. Lond. Ser. Contain. Pap. Math. Phys. Character 1932, 138, 55–83. [Google Scholar]
- Kermack, W.O.; McKendrick, A.G. Contributions to the mathematical theory of epidemics. III.Further studies of the problem of endemicity. Proc. R. Soc. Lond. Ser. Contain. Pap. Math. Phys. Character 1933, 141, 94–122. [Google Scholar]
- Chen, S.S.; Small, M.; Fu, X.C. Global stability of epidemic models with imperfect vaccination and quarantine on scale-free networks. IEEE Trans. Netw. Sci. Eng. 2019, 7, 1583–1596. [Google Scholar] [CrossRef]
- Peter, O.J.; Adebisi, A.F.; Ajisope, M.O.; Ajibade, F.O.; Abioye, A.I.; Oguntolu, F.A. Global stability analysis of typhoid fever model. Adv. Syst. Sci. Appl. 2020, 20, 20–31. [Google Scholar]
- Jose, S.A.; Raja, R.; Dianavinnarasi, J.; Baleanu, D.; Jirawattanapanit, A. Mathematical modeling of chickenpox in Phuket: Efficacy of precautionary measures and bifurcation analysis. Biomed. Signal Process. Control 2023, 84, 104714. [Google Scholar] [CrossRef]
- Hethcote, H.; Zhi En, M.; Sheng Bing, L. Effects of quarantine in six endemic models for infectious diseases. Math. Biosci. 2002, 180, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Suo, Y.H. Asymptotical Stability of an SIQS Epidemic Model with Age Dependence and Generally Nonlinear Contact Rate. Appl. Mech. Mater. 2011, 58, 292–297. [Google Scholar] [CrossRef]
- Wei, F.Y.; Chen, F.X. Stochastic permanence of an SIQS epidemic model with saturated incidence and independent random perturbations. Phys. A Stat. Mech. Its Appl. 2016, 453, 99–107. [Google Scholar] [CrossRef]
- Zhang, X.B.; Liu, R.J. The stationary distribution of a stochastic SIQS epidemic model with varying total population size. Appl. Math. Lett. 2021, 116, 106974. [Google Scholar] [CrossRef]
- Barabási, A.L.; Albert, R. Emergence of scaling in random networks. Science 1999, 509–512, 286. [Google Scholar] [CrossRef]
- Pastor-Satorras, R.; Vespignani, A. Epidemic dynamics and endemic states in complex networks. Phys. Rev. E 2001, 63, 066117. [Google Scholar] [CrossRef]
- Cheng, X.X.; Wang, Y.; Huang, G. Global dynamics of a network-based SIQS epidemic model with nonmonotone incidence rate. Chaos Solitons Fractals 2021, 153, 111502. [Google Scholar] [CrossRef]
- Zhao, R.D.; Liu, Q.M.; Sun, M.C. Dynamical behavior of a stochastic SIQS epidemic model on scale-free networks. J. Appl. Math. Comput. 2022, 68, 813–838. [Google Scholar] [CrossRef]
- Li, T.; Wang, Y.M.; Guan, Z.H. Spreading dynamics of a SIQRS epidemic model on scale-free networks. Commun. Nonlinear Sci. Numer. Simul. 2014, 19, 686–692. [Google Scholar] [CrossRef]
- Kang, H.Y.; Liu, K.H.; Fu, X.C. Dynamics of an epidemic model with quarantine on scale-free networks. Phys. Lett. A 2017, 381, 3945–3951. [Google Scholar] [CrossRef]
- Wang, H.Y.; Moore, J.M.; Small, M.; Wang, J.; Yang, H.; Gu, C. Epidemic dynamics on higher-dimensional small world networks. Appl. Math. Comput. 2022, 421, 126911. [Google Scholar] [CrossRef]
- Iacoviello, D.; Stasio, N. Optimal control for SIRC epidemic outbreak. Comput. Methods Programs Biomed. 2013, 110, 333–342. [Google Scholar] [CrossRef]
- Buonomo, B.; Lacitignola, D.; Vargas-De-León, C. Qualitative analysis and optimal control of an epidemic model with vaccination and treatment. Math. Comput. Simul. 2014, 100, 88–102. [Google Scholar] [CrossRef]
- Kandhway, K.; Kuri, J. How to run a campaign: Optimal control of SIS and SIR information epidemics. Appl. Math. Comput. 2014, 231, 79–92. [Google Scholar] [CrossRef]
- Kandhway, K.; Kuri, J. Optimal control of information epidemics modeled as Maki Thompson rumors. Commun. Nonlinear Sci. Numer. Simul. 2014, 19, 4135–4147. [Google Scholar] [CrossRef]
- Jang, J.; Kwon, H.D.; Lee, J. Optimal control problem of an SIR reaction–diffusion model with inequality constraints. Math. Comput. Simul. 2020, 171, 136–151. [Google Scholar] [CrossRef]
- Wang, B.; Tian, X.H.; Xu, R.; Song, C.W. Threshold dynamics and optimal control of a dengue epidemic model with time delay and saturated incidence. J. Appl. Math. Comput. 2023, 69, 871–893. [Google Scholar] [CrossRef]
- Chen, L.J.; Sun, J.T. Global stability and optimal control of an SIRS epidemic model on heterogeneous networks. Phys. A Stat. Mech. Its Appl. 2014, 410, 196–204. [Google Scholar] [CrossRef]
- Chen, L.J.; Sun, J.T. Optimal vaccination and treatment of an epidemic network model. Phys. Lett. A 2014, 378, 3028–3036. [Google Scholar] [CrossRef]
- Xu, D.G.; Xu, X.Y.; Xie, Y.F.; Yang, C.H. Optimal control of an SIVRS epidemic spreading model with virus variation based on complex networks. Commun. Nonlinear Sci. Numer. Simul. 2017, 48, 200–210. [Google Scholar] [CrossRef]
- Jia, N.; Ding, L.; Liu, Y.J.; Hu, P. Global stability and optimal control of epidemic spreading on multiplex networks with nonlinear mutual interaction. Phys. A Stat. Mech. Its Appl. 2018, 502, 93–105. [Google Scholar] [CrossRef]
- Li, K.Z.; Zhu, G.H.; Ma, Z.J.; Chen, L.J. Dynamic stability of an SIQS epidemic network and its optimal control. Commun. Nonlinear Sci. Numer. Simul. 2019, 66, 84–95. [Google Scholar] [CrossRef]
- Wei, X.D.; Xu, G.C.; Zhou, W.S. Global stability of endemic equilibrium for a SIQRS epidemic model on complex networks. Phys. A Stat. Mech. Its Appl. 2018, 512, 203–214. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, M.X.; Xie, B.L. Optimal control of an SIQRS epidemic model with three measures on networks. Nonlinear Dyn. 2021, 103, 2097–2107. [Google Scholar] [CrossRef]
- Yang, P.; Jia, J.B.; Shi, W.; Feng, J.W.; Fu, X.C. Stability analysis and optimal control in an epidemic model on directed complex networks with nonlinear incidence. Commun. Nonlinear Sci. Numer. Simul. 2023, 121, 107206. [Google Scholar] [CrossRef]
- Pastor-Satorras, R.; Vespignani, A. Epidemic dynamics in finite size scale-free networks. Phys. Rev. E 2002, 65, 035108. [Google Scholar] [CrossRef]
- Moreno, Y.; Pastor-Satorras, R.; Vespignani, A. Epidemic outbreaks in complex heterogeneous networks. Eur. Phys. J.-Condens. Matter Complex Syst. 2002, 26, 521–529. [Google Scholar] [CrossRef]
- Wang, L.; Dai, G.Z. Global stability of virus spreading in complex heterogeneous networks. SIAM J. Appl. Math. 2008, 68, 1495–1502. [Google Scholar] [CrossRef]
- Yang, R.; Wang, B.H.; Ren, J.; Bai, W.J.; Shi, Z.W.; Wang, W.X.; Zhou, T. Epidemic spreading on heterogeneous networks with identical infectivity. Phys. Lett. A 2007, 364, 189–193. [Google Scholar] [CrossRef]
- Chu, X.W.; Zhang, Z.Z.; Guan, J.H.; Zhou, S.G. Epidemic spreading with nonlinear infectivity in weighted scale-free networks. Phys. A Stat. Mech. Its Appl. 2011, 390, 471–481. [Google Scholar] [CrossRef]
- Zhang, H.F.; Fu, X.C. Spreading of epidemics on scale-free networks with nonlinear infectivity. Nonlinear Anal. Theory Methods Appl. 2009, 70, 3273–3278. [Google Scholar] [CrossRef]
- Zhu, G.H.; Chen, G.R.; Xu, X.J.; Fu, X.C. Epidemic spreading on contact networks with adaptive weights. J. Theor. Biol. 2013, 317, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.N.; Takeuchi, Y.; Iwami, S. SVIR epidemic models with vaccination strategies. J. Theor. Biol. 2008, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lajmanovich, A.; Yorke, J.A. A deterministic model for gonorrhea in a nonhomogeneous population. Math. Biosci. 1976, 28, 221–236. [Google Scholar] [CrossRef]
- Culshaw, R.V.; Ruan, S.; Spiteri, R.J. Optimal HIV treatment by maximising immune response. J. Math. Biol. 2004, 48, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Y.; Cheng, S.M.; Chen, K.C. Optimal control of epidemic information dissemination over networks. IEEE Trans. Cybern. 2014, 44, 2316–2328. [Google Scholar] [CrossRef]
- Abboubakar, H.; Guidzavai, A.K.; Yangla, J.; Damakoa, I.; Mouangue, R. Mathematical modeling and projections of a vector-borne disease with optimal control strategies: A case study of the Chikungunya in Chad. Chaos Solitons Fractals 2021, 150, 111197. [Google Scholar] [CrossRef]
- Fleming, W.; Rishel, R. Deterministic and Stochastic Optimal Control; Springer: Berlin/Heidelberg, Germany, 1975; p. 222. [Google Scholar]
- Fu, X.C.; Small, M.; Walker, D.M.; Zhang, H.F. Epidemic dynamics on scale-free networks with piecewise linear infectivity and immunization. Phys. Rev. E 2008, 77, 036113. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbols | Description |
|---|---|
| Proportion of nodes with degree k. | |
| Average degree . | |
| n | Maximum degree. |
| b | Birth rate. |
| d | Natural death rate. |
| Fertile contact probability between a node with degree k and its neighbors. | |
| Transmission rate of infected nodes with degree k. | |
| Vaccination rate of susceptible nodes with degree k. | |
| Quarantine rate of infected nodes with degree k. | |
| Recovery rate of infected nodes. | |
| Recovery rate of quarantined nodes. |
| Basic reproduction number | 0.1734 | 0.6590 | 0.9538 | 5.3671 | 7.1561 | 8.9451 |
| Final infection scale | 0 | 0 | 0 | 0.0449 | 0.0597 | 0.0724 |
| Quarantining Control Strategies | Final Infection Scale | Epidemic Threshold |
|---|---|---|
| No quarantine | 0.0713 | 0.0468 |
| Acquaintance quarantine | 0.0568 | 0.0724 |
| Proportional quarantine | 0.0424 | 0.0798 |
| Target quarantine | 0.0182 | 0.1483 |
| Optimal Control | Max Control | No Control |
|---|---|---|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Chen, S.; Yu, D.; Liu, L.; Shang, K.-K. Propagation Dynamics of an Epidemic Model with Heterogeneous Control Strategies on Complex Networks. Symmetry 2024, 16, 166. https://doi.org/10.3390/sym16020166
Wang Y, Chen S, Yu D, Liu L, Shang K-K. Propagation Dynamics of an Epidemic Model with Heterogeneous Control Strategies on Complex Networks. Symmetry. 2024; 16(2):166. https://doi.org/10.3390/sym16020166
Chicago/Turabian StyleWang, Yan, Shanshan Chen, Dingguo Yu, Lixiang Liu, and Ke-Ke Shang. 2024. "Propagation Dynamics of an Epidemic Model with Heterogeneous Control Strategies on Complex Networks" Symmetry 16, no. 2: 166. https://doi.org/10.3390/sym16020166
APA StyleWang, Y., Chen, S., Yu, D., Liu, L., & Shang, K. -K. (2024). Propagation Dynamics of an Epidemic Model with Heterogeneous Control Strategies on Complex Networks. Symmetry, 16(2), 166. https://doi.org/10.3390/sym16020166