
Lateralization of the Avian Magnetic Compass: Analysis of Its Early Plasticity

Abstract
:1. Introduction
2. Results
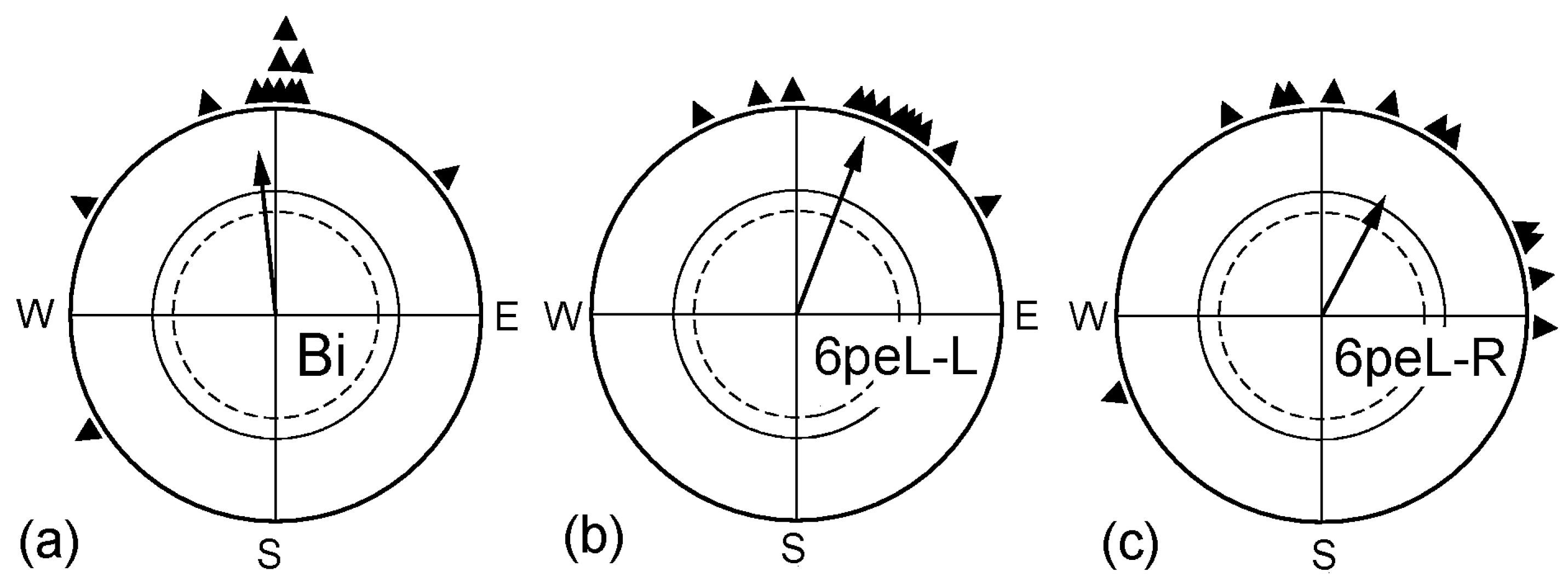
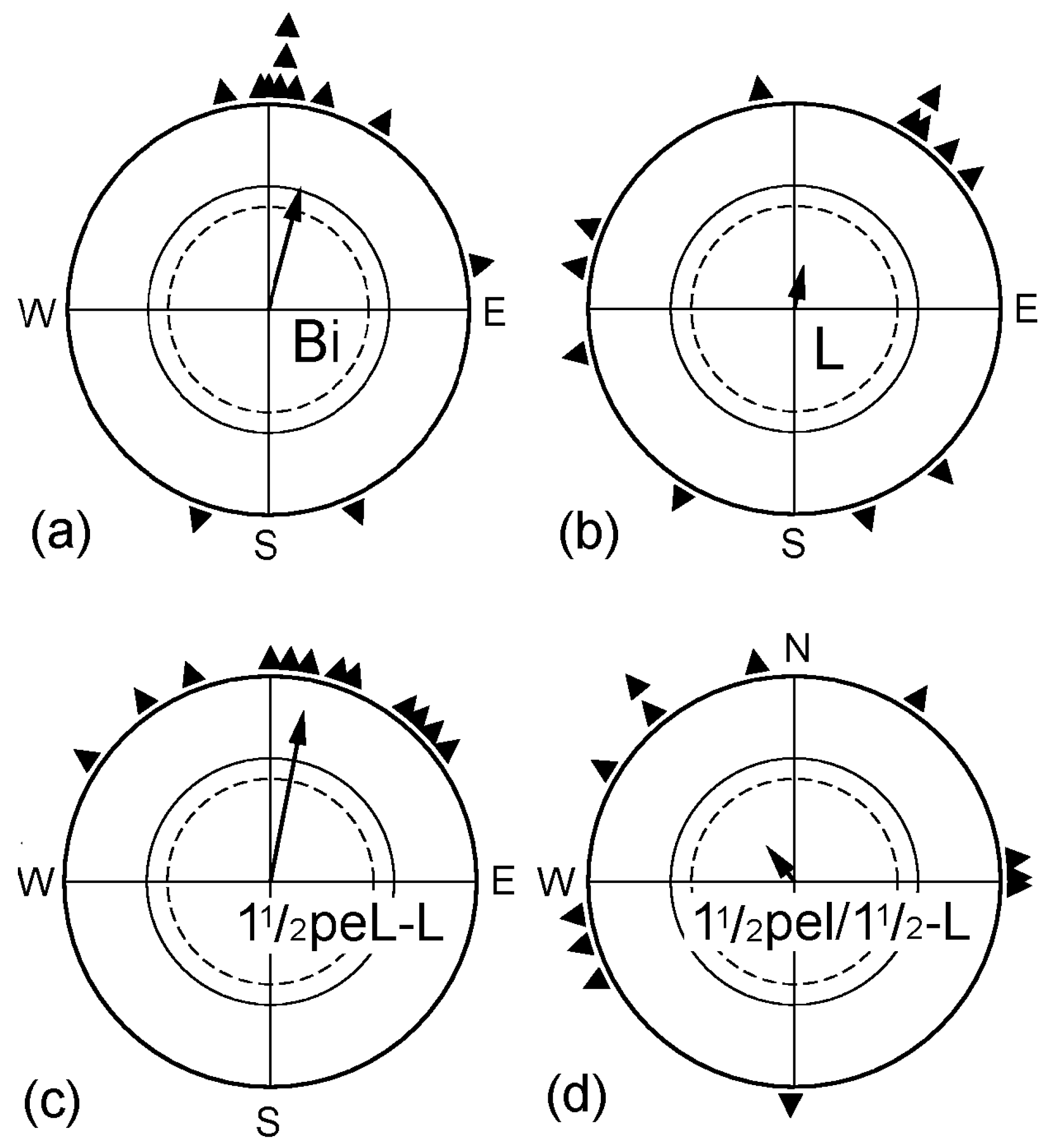
2.1. The Effect of Monocular Pre-Exposure
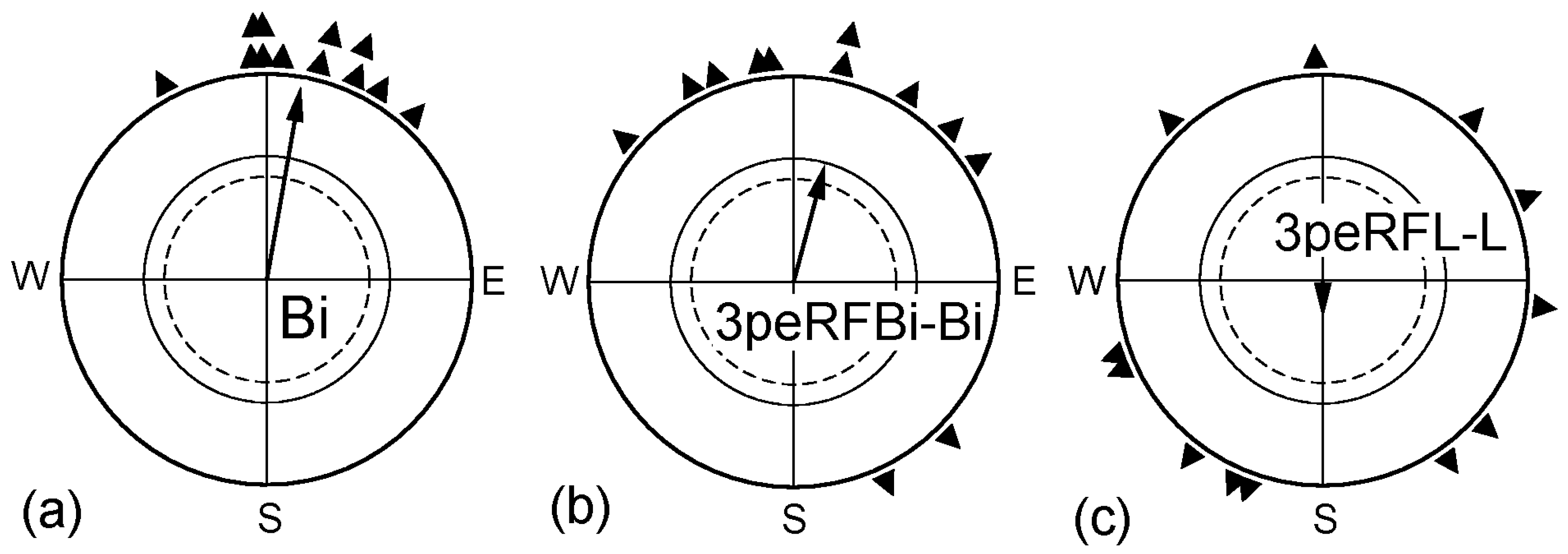
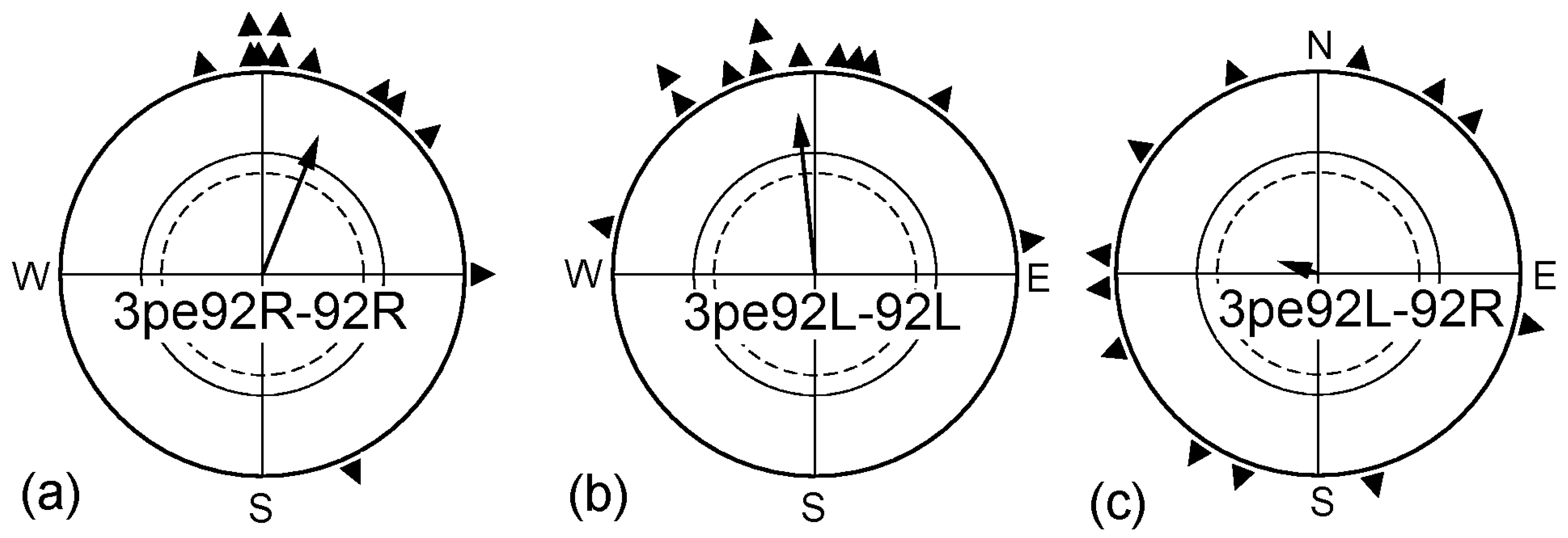
2.2. Pre-Exposure in Altered Magnetic Conditions
3. Discussion
3.1. Fast Re-Activation of the Left Eye/Right Hemisphere System for Sensing Magnetic Directions
3.2. Conditions Required for the Re-Activation of the Left Eye/Right Hemisphere System
3.3. Lasting Flexibility in the Avian Magnetic Compass?
4. Material and Methods
4.1. Experimental Birds
4.2. Covering One Eye
4.3. Test Performance
4.4. Data Analysis and Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Ethical Statement
References
- Ocklenburg, S.; Ströckens, F.; Güntürkün, O. Lateralisation of conspecific vocalisation in non-human vertebrates. Laterality 2013, 18, 1–31. [Google Scholar] [CrossRef]
- Ströckens, F.; Güntürkün, O.; Ocklenburg, S. Limb preferences in non-human vertebrates. Laterality 2013, 18, 536–575. [Google Scholar] [CrossRef]
- Güntürkün, O.; Diekamp, B.; Manns, M.; Nottelmann, F.; Prior, H.; Schwarz, A.; Skiba, M. Asymmetry pays: Visual lateralization improves discrimination success in pigeons. Curr. Biol. 2000, 10, 1079–1081. [Google Scholar] [CrossRef]
- Dadda, M.; Bisazza, A. Does brain asymmetry allow efficient performance of simultaneous tasks? Anim. Behav. 2006, 72, 523–529. [Google Scholar] [CrossRef]
- Hirnstein, M.; Hugdahl, K.; Hausmann, M. How brain asymmetry relates to performance—A large-scale dichotic listening study. Front. Psychol. 2014, 4, 997. [Google Scholar] [CrossRef]
- Ventolini, N.; Ferrero, E.A.; Sponza, S.; Della Chiesa, A.; Zucca, P.; Vallortigara, G. Laterality in the wild: Preferential hemifield use during predatory and sexual behaviour in the black-winged stilt. Anim. Behav. 2005, 69, 1077–1084. [Google Scholar] [CrossRef]
- Gülbetekin, E.; Güntürkün, O.; Dural, S.; Cetinkaya, H. Visual asymmetries in Japanese quail (Coturnix japonica) retain a lifelong potential for plasticity. Behav. Neurosci. 2009, 123, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Chiandetti, C.; Sovrano, V.A. Brain asymmetry (animal). WIREs Cogn. Sci. 2011, 2, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Zucca, P.; Vallortigara, G. Advantages of having a lateralized brain. Proc. R. Soc. B 2004, 271 (Suppl. S6), S420–S422. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G. The evolutionary psychology of left and right: Costs and benefits of lateralization. Dev. Psychobiol. 2006, 48, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, W.; Traudt, J.; Güntürkün, O.; Prior, H.; Wiltschko, R. Lateralisation of magnetic compass orientation in a migratory bird. Nature 2002, 419, 467–470. [Google Scholar] [CrossRef]
- Wiltschko, W.; Munro, U.; Ford, H.; Wiltschko, R. Lateralisation of magnetic compass orientation in Silvereyes, Zosterops lateralis. Aust. J. Zool. 2003, 51, 597–602. [Google Scholar] [CrossRef]
- Rogers, L.; Munro, U.; Freire, R.; Wiltschko, R.; Wiltschko, W. Lateralized response of chicks to magnetic cues. Behav. Brain Res. 2008, 186, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Stapput, K.; Güntürkün, O.; Hoffmann, K.P.; Wiltschko, R.; Wiltschko, W. Magnetoreception of directional information requires non-degraded vision. Curr. Biol. 2010, 20, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Wilzeck, C.; Wiltschko, W.; Güntürkün, O.; Wiltschko, R.; Prior, H. Lateralization of magnetic compass orientation in pigeons. J. R. Soc. Interface 2010, 7, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Ritz, T.; Adem, S.; Schulten, K. A model for photoreceptor-based magnetoreception in birds. Biophys. J. 2000, 78, 707–718. [Google Scholar] [CrossRef]
- Henbest, K.B.; Kukura, P.; Rodgers, C.T.; Hore, P.J.; Timmel, C.R. Radio frequency magnetic field effects on a radical recombination reaction: A diagnostic test for the radical pair mechanism. J. Am. Chem. Soc. 2004, 126, 8102–8103. [Google Scholar] [CrossRef] [PubMed]
- Ritz, T.; Thalau, P.; Philllips, J.B.; Wiltschko, R.; Wiltschko, W. Resonance effects indicate a radical-pair mechanism for avian magnetic compass. Nature 2004, 429, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Thalau, P.; Ritz, T.; Stapput, K.; Wiltschko, R.; Wiltschko, W. Magnetic compass orientation of migratory birds in the presence of a 1.315 MHz oscillating field. Naturwissenschaften 2005, 92, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, W.; Freire, R.; Munro, U.; Ritz, T.; Rogers, L.; Thalau, P.; Wiltschko, R. The magnetic compass of domestic chickens, Gallus gallus. J. Exp. Biol. 2007, 210, 2300–2310. [Google Scholar] [CrossRef] [PubMed]
- Keary, N.; Roploh, T.; Voss, J.; Thalau, P.; Wiltschko, R.; Wiltschko, W.; Bischof, H.J. Oscillating magnetic field disrupts magnetic orientation in Zebra finches, Taeniopygia guttata. Front. Zool. 2009, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Kavokin, K.; Chernetsov, N.; Pakomov, A.; Bojarinova, J.; Kobylkov, D.; Namozov, B. Magnetic orientation of garden warblers (Sylvia borin) under 1.4 MHz radio frequency field. J. R. Soc. Interface 2014, 11, 20140451. [Google Scholar] [CrossRef] [PubMed]
- Nießner, C.; Denzau, S.; Gross, J.C.; Peichl, L.; Bischof, H.J.; Fleissner, G.; Wiltschko, W.; Wiltschko, R. Avian ultraviolet/violet cones identified as probable magnetoreceptors. PLoS ONE 2011, 6, e20091. [Google Scholar] [CrossRef] [PubMed]
- Nießner, C.; Denzau, S.; Stapput, K.; Ahmad, M.; Peichl, L.; Wiltschko, W.; Wiltschko, R. Magnetoreception: Activated cryptochrome 1a concurs with magnetic orientation in birds. J. R. Soc. Interface 2013, 10, 20130618. [Google Scholar]
- Hein, C.M.; Zapka, M.; Heyers, D.; Kurzschbauch, S.; Schneider, N.-L.; Mouritsen, H. Night-migratory Garden Warblers can orient with their magnetic compass using the left, the right or both eyes. J. R. Soc. Interface 2010, 7, S227–S233. [Google Scholar] [CrossRef] [PubMed]
- Hein, C.M.; Engels, S.; Kishkinev, D.; Mouritsen, H. Robins have a magnetic compass in both eye. Nature 2011, 271, E11–E12. [Google Scholar] [CrossRef] [PubMed]
- Engels, S.; Hein, C.; Lefeldt, N.; Prior, H.; Mouritsen, H. Night-migratory songbirds possess a magnetic compass in both eye. PLoS ONE 2012, 7, e43271. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, W.; Traudt, J.; Güntürkün, O.; Prior, H.; Wiltschko, R. Reply to Hein et al. Nature 2011, 471, E12–E13. [Google Scholar]
- Gehring, D.; Wiltschko, W.; Güntürkün, O.; Denzau, S.; Wiltschko, R. Development of lateralization of the magnetic compass in a migratory bird. Proc. R. Soc. B 2012, 279, 4230–4235. [Google Scholar] [CrossRef] [PubMed]
- Batschelet, E. Circular Statistics in Biology; Academic Press: London, UK, 1981. [Google Scholar]
- Wiltschko, W.; Stapput, K.; Thalau, P.; Wiltschko, R. Avian magnetic compass: Fast adjustment to intensities outside the normal functional window. Naturwissenschaften 2006, 93, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, R.; Wiltschko, W. Sensing magnetic directions in birds: Radical pair processes involving cryptochrome. Biosensors 2014, 4, 221–242. [Google Scholar] [CrossRef] [PubMed]
- Semm, P.; Demaine, C. Neurophysiological properties of magnetic cells in the pigeon’s visual system. J. Comp. Physiol. A 1986, 159, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Heyers, D.; Manns, M.; Luksch, H.; Güntürkün, O.; Mouritsen, H. A visual pathway links brain structures active during magnetic compass orientation in migratory birds. PLoS ONE 2007, 9, e937. [Google Scholar] [CrossRef] [PubMed]
- Zapka, M.; Heyers, D.; Hein, C.M.; Engels, S.; Schneider, N.L.; Hans, J.; Weiler, S.; Dreyer, D.; Kishkinev, D.; Wild, J.M.; et al. Visual but not trigeminal mediation of magnetic compass information in a migratory bird. Nature 2009, 461, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, H.; Heyers, D.; Güntürkün, O. The neural basis of long-distance navigation in birds. Annu. Rev. Physiol. 2016, 78, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O. Morphological asymmetries of the tectum opticum in the pigeon. Exp. Brain Res. 1997, 116, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Hellmann, B.; Melsbach, G.; Prior, H. Asymmetries of representation in the visual system of pigeons. Neuroreport 1998, 9, 4127–4130. [Google Scholar] [CrossRef] [PubMed]
- Ströckens, F.; Freund, N.; Manns, M.; Ocklenburg, S.; Güntürkün, O. Visual asymmetries and the ascending thalamofugal pathway in pigeons. Brain Struct. Funct. 2013, 218, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Güntürkün, O. “Natural” and artificial monocular deprivation effects on thalamic soma sizes in pigeons. Neuroreport 1999, 10, 3223–3228. [Google Scholar] [CrossRef] [PubMed]
- Folta, K.; Diekamp, B.; Güntürkün, O. Asymmetrical modes of visual bottom-up and top-down integration in the thalamic nucleus rotundus of pigeons. J. Neurosci. 2004, 24, 9475–9485. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Hahmann, U. Visual acuity and hemispheric asymmetries in pigeons. Behav. Brain Res. 1994, 60, 171–175. [Google Scholar] [CrossRef]
- Valencia-Alfonso, C.-E.; Verhaal, J.; Güntürkün, O. Ascending and descending mechanisms of visual lateralization in pigeons. Philos. Trans. Roy. Soc. B 2009, 364, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Freund, N.; Valencia-Alfonso, C.E.; Kirsch, J.; Brodmann, K.; Manns, M.; Güntürkün, O. Asymmetric top-down modulation of ascending visual pathways in pigeons. Neuropsychologia 2016, 83, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Aust, U.; Huber, L.; Hausmann, M.; Güntürkün, O. Lateralized cognition: Asymmetrical and complementary strategies of pigeons during discrimination of the “human concept”. Cognition 2007, 104, 315–344. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Asymmetry of brain and behavior in animals: Its development, function, and human relevance. Genesis 2014, 52, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Keysers, C.; Diekamp, B.; Güntürkün, O. Evidence for physiological asymmetries in the intertectal connections of the pigeon (Columba livia) and their potential role in brain lateralisation. Brain Res. 2000, 852, 406–413. [Google Scholar] [CrossRef]
- Genç, E.; Ocklenburg, S.; Singer, W.; Güntürkün, O. Abnormal interhemispheric motor interactions in patients with callosal agenesis. Behav. Brain Res. 2015, 293, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.H.; Kandel, E.R.; Harris, K.M. Structural components of synaptic plasticity and memory consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Liedvogel, M.; Feender, G.; Wada, K.; Troje, N.F.; Jarvis, E.D.; Mouritsen, H. Lateralized activation of cluster N in the brain of migratory songbird. Eur. J. Neurosci. 2007, 25, 116–1173. [Google Scholar] [CrossRef] [PubMed]
- Nottelmann, F.; Wohlschläger, A.; Güntürkün, O. Unihemispheric memory in pigeons—Knowledge, the left hemisphere is reluctant to share. Behav. Brain Res. 2002, 133, 309–315. [Google Scholar] [CrossRef]
- Xiao, Q.; Güntürkünn, O. Natural split brains? Lateralized memory for task contingencies in pigeons. Neurosci. Lett. 2009, 458, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Zapka, M.; Heyers, D.; Liedvogel, M.; Jarvis, E.D.; Mouritsen, H. Night-time neuronal activation of cluster N in a day- and night-migration songbird. Eur. J. Neurosci. 2010, 32, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Nießner, C.; Ernst Strüngmann Institut, Frankfurt am Main, Germany. Personal communication, 2013.
- Rogers, L.J. Light input and the reversal of functional lateralization in the chicken brain. Behav. Brain Res. 1990, 38, 211–221. [Google Scholar] [CrossRef]
- Manns, M.; Güntürkün, O. Monocular deprivation alters the direction of functional and morphological asymmetries in the pigeon’s visual system. Behav. Neurosci. 1999, 113, 1–10. [Google Scholar] [CrossRef]
- Skiba, M.; Diekamp, B.; Güntürkün, O. Embryonic light stimulation induces different asymmetries in visuoperceptual and visuomotor pathways of pigeons. Behav. Brain Res. 2002, 134, 149–156. [Google Scholar] [CrossRef]
- Hausmann, M. Why sex hormones matter for neuroscience: A very short review on sex, sex hormones, and functional brain asymmetries. J. Neurosci. Res. 2017, 95, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, H.; Feender, G.; Hegemann, A.; Liedvogel, M. Thermopaer can replace typewriter correction paper in Emlen funnels. J. Ornithol. 2009, 150, 713–715. [Google Scholar] [CrossRef]
- Wiltschko, R.; Thalau, P.; Gehring, D.; Nießner, C.; Ritz, T.; Wiltschko, W. Magnetoreception in birds: The effect of radio frequency fields. J. R. Soc. Interface 2015, 12, 20141103. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Test Condition |
|---|---|
| Bi | binocularly tested, control |
| L | monocularly left-eyed |
| 6peL-L | 6 h pre-exposed monocularly left-eyed, tested left-eyed |
| 6peL-R | 6 h pre-exposed monocularly left-eyed, tested right-eyed |
| 1½peL-L | 1½ h pre-exposed monocularly left-eyed, tested left-eyed |
| 1½peL/1½-L | 1½ h pre-exposed monocularly left-eyed, then 1½ h without eye cover, tested left-eyed |
| 3peRFBi-Bi | 3 h binocularly pre-exposed in an RF field (1.314 MHz, 480 nT), tested binocularly |
| 3peRFL-L | 3 h monocularly left-eyed pre-exposed in a RF field(1.314 MHz, 480 nT), tested left-eyed |
| 3pe92R-92R | 3 h right-eyed pre-exposed in a 92 µT field, tested right-eyed in the 92 µT field |
| 3pe92L-92L | 3 h left-eyed pre-exposed in a 92 µT field, tested left-eyed in the 92 µT field |
| 3pe92L-92R | 3 h left-eyed pre-exposed in a 92 µT field, tested right-eyed in the 92 µT field |
| Year | Condition | Test Magnetic Field | n | Med. rb | αN | rN | ΔBi | ΔX |
|---|---|---|---|---|---|---|---|---|
| 2011 | Bi | geomagnetic field | 4 | 0.79 | 354° | 0.80 *** | ||
| “ | 6peL-L | “ | 3 | 0.81 | 21° | 0.92 *** | +27° *d | X1 |
| “ | 6peL-R | “ | 3 | 0.91 | 28° | 0.66 ** | +32° n.s | +7 *s |
| 2012 | Bi | “ | 3 | 0.45 | 15° | 0.62 ** | ||
| “ | L | “ | 3 | 0.48 | (12°) | 0.22 n.s. | (−3°) *s | X2 |
| “ | 1½peL-L | “ | 3 | 0.77 | 11° | 0.85 *** | −4°n.s. | (−11° *s) |
| “ | 1½peL/1½-L | “ | 3 | 0.60 | (322°) | 0.22 n.s. | (−53°) *d | (−50° n.s.) |
| 2013 I | Bi | geomagnetic field | 3 | 0.92 | 351° | 0.65 ** | ||
| “ | 3pe92R-92R | 92 nT | 3 | 0.90 | 22° | 0.74 *** | +31°n.s. | |
| 2013 II | Bi | geomagnetic field | 3 | 0.82 | 10° | 0.95 *** | ||
| “ | 3peRFBi-Bi | “ | 3 | 0.44 | 15° | 0.59 * | +5° *s | X3 |
| “ | 3peRFL-L | “ | 3 | 0.41 | (180°) | 0.18 n.s. | (+170°) ***s | (−165° *s) |
| “ | 3pe92L-92L | 92 nT | 4 | 0.50 | 354° | 0.80 *** | −16° n.s. | X4 |
| “ | 3pe92L-92R | 92 nT | 3 | 0.83 | (287°) | 0.21 n.s. | (−103°) *** | (−67° **s) |
| Test Round | Monocularly Right-Eyed Birds | Monocularly Left-Eyed Birds | ||||||
|---|---|---|---|---|---|---|---|---|
| N | α | r | Δ Round 1 | N | α | r | Δ Round 1 | |
| 1 | 12 | 8° | 0.87 *** | 11 | (304°) | 0.38 n.s. | ||
| 2 | 11 | (20°) | 0.46 n.s | +12° n.s. | 12 | (10°) | 0.36 n.s. | +66° n.s. |
| 3 | 10 | 22° | 0.64 ** | +14° n.s. | 11 | 3° | 0.54 * | +59° n.s. |
| 4 | 12 | 13° | 0.74 *** | +69° * | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gehring, D.; Güntürkün, O.; Wiltschko, W.; Wiltschko, R. Lateralization of the Avian Magnetic Compass: Analysis of Its Early Plasticity. Symmetry 2017, 9, 77. https://doi.org/10.3390/sym9050077
Gehring D, Güntürkün O, Wiltschko W, Wiltschko R. Lateralization of the Avian Magnetic Compass: Analysis of Its Early Plasticity. Symmetry. 2017; 9(5):77. https://doi.org/10.3390/sym9050077
Chicago/Turabian StyleGehring, Dennis, Onur Güntürkün, Wolfgang Wiltschko, and Roswitha Wiltschko. 2017. "Lateralization of the Avian Magnetic Compass: Analysis of Its Early Plasticity" Symmetry 9, no. 5: 77. https://doi.org/10.3390/sym9050077
APA StyleGehring, D., Güntürkün, O., Wiltschko, W., & Wiltschko, R. (2017). Lateralization of the Avian Magnetic Compass: Analysis of Its Early Plasticity. Symmetry, 9(5), 77. https://doi.org/10.3390/sym9050077