
Bonding Behavior and Mechanism of U(VI) by Chemically Modified Deinococcus radiodurans


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material and Bacteria
2.2. Chemical Modifications of Biomass
2.3. Batch Experiment
2.4. Characterization of U-Loaded and Unloaded Samples
2.5. Biosorption Kinetics
3. Results
3.1. SEM Observations of the Raw and Chemically Modified Biomass
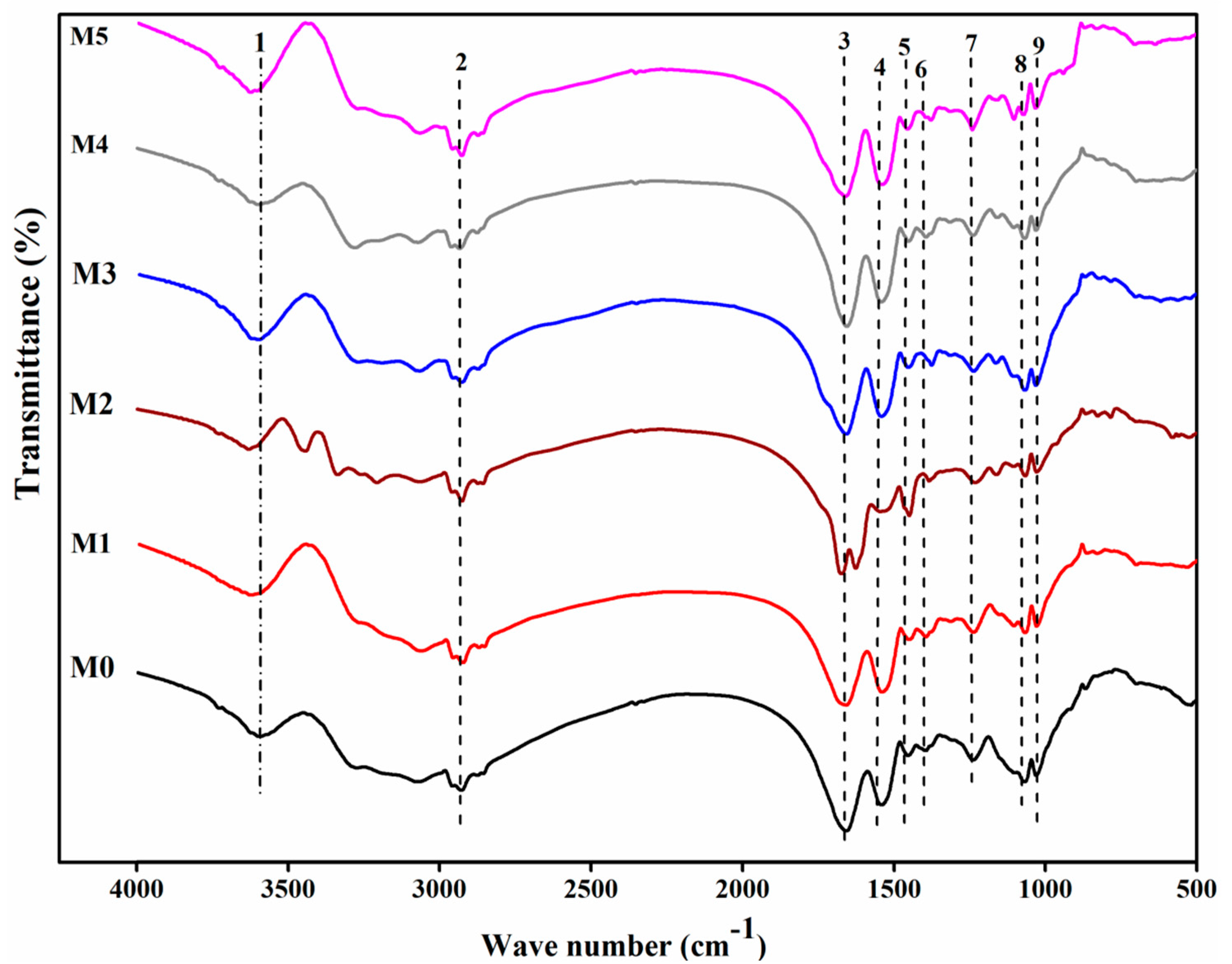
3.2. Changes in the Functional Groups of Chemically Modified D. radiodurans
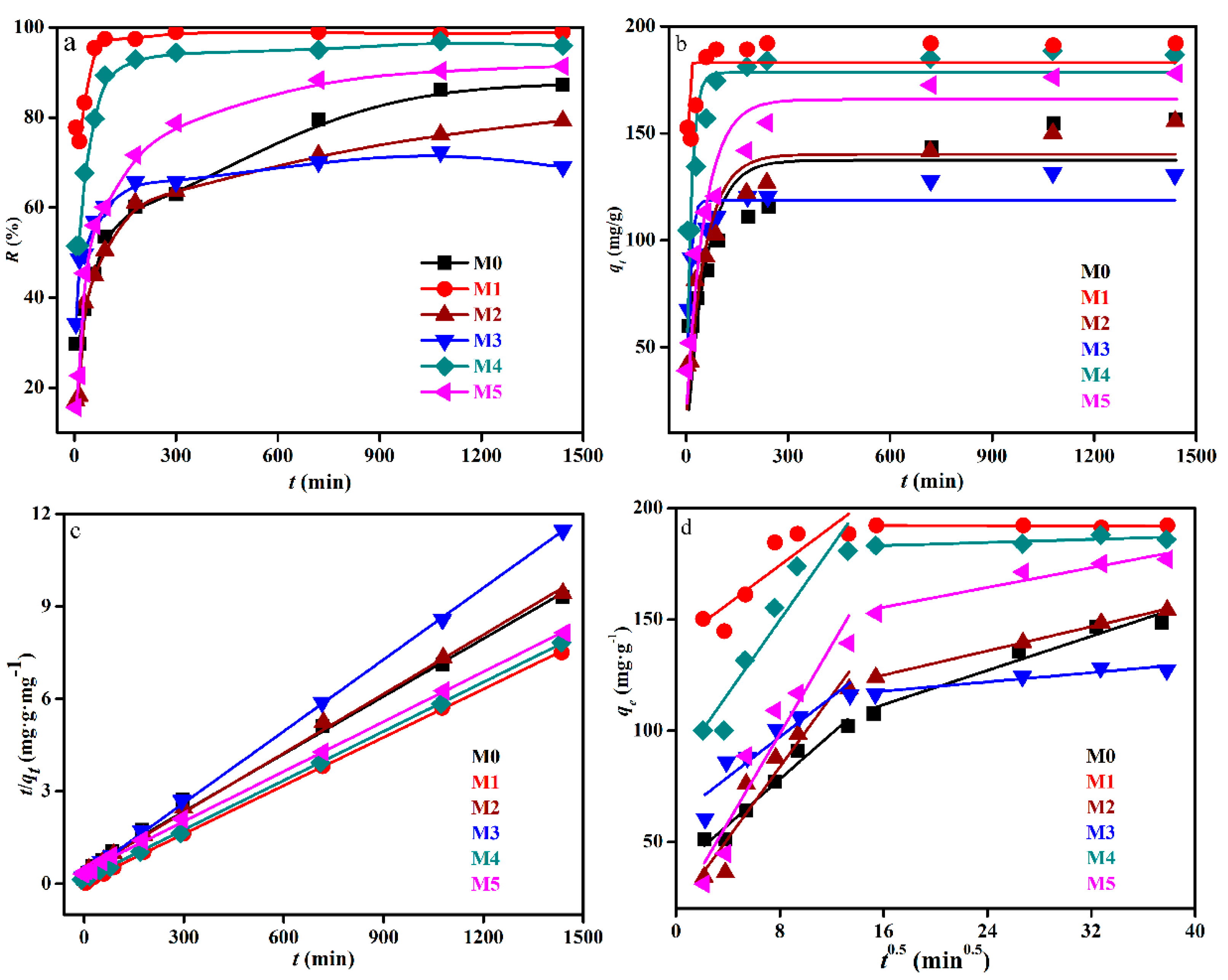
3.3. Removal Influenced by Time
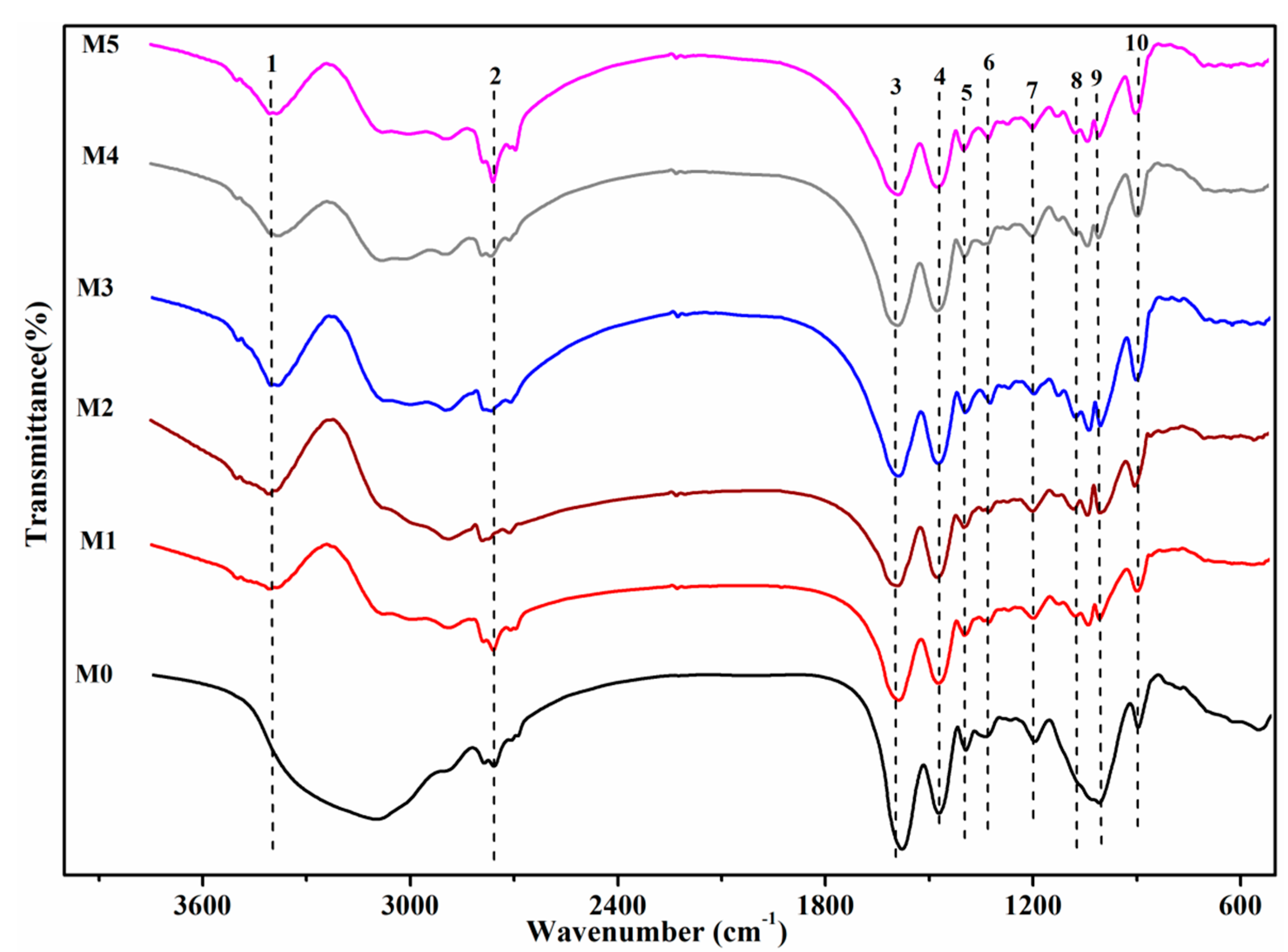
3.4. Characterization of the Surface Properties Change after U(VI) Biosorption
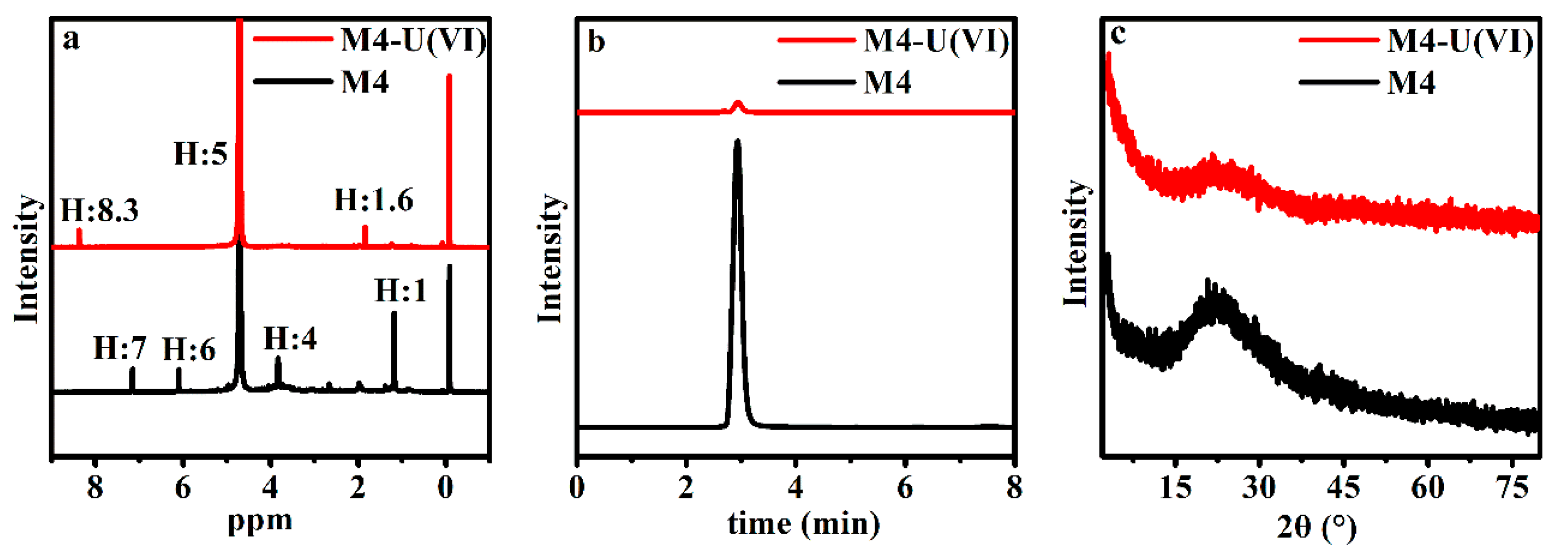
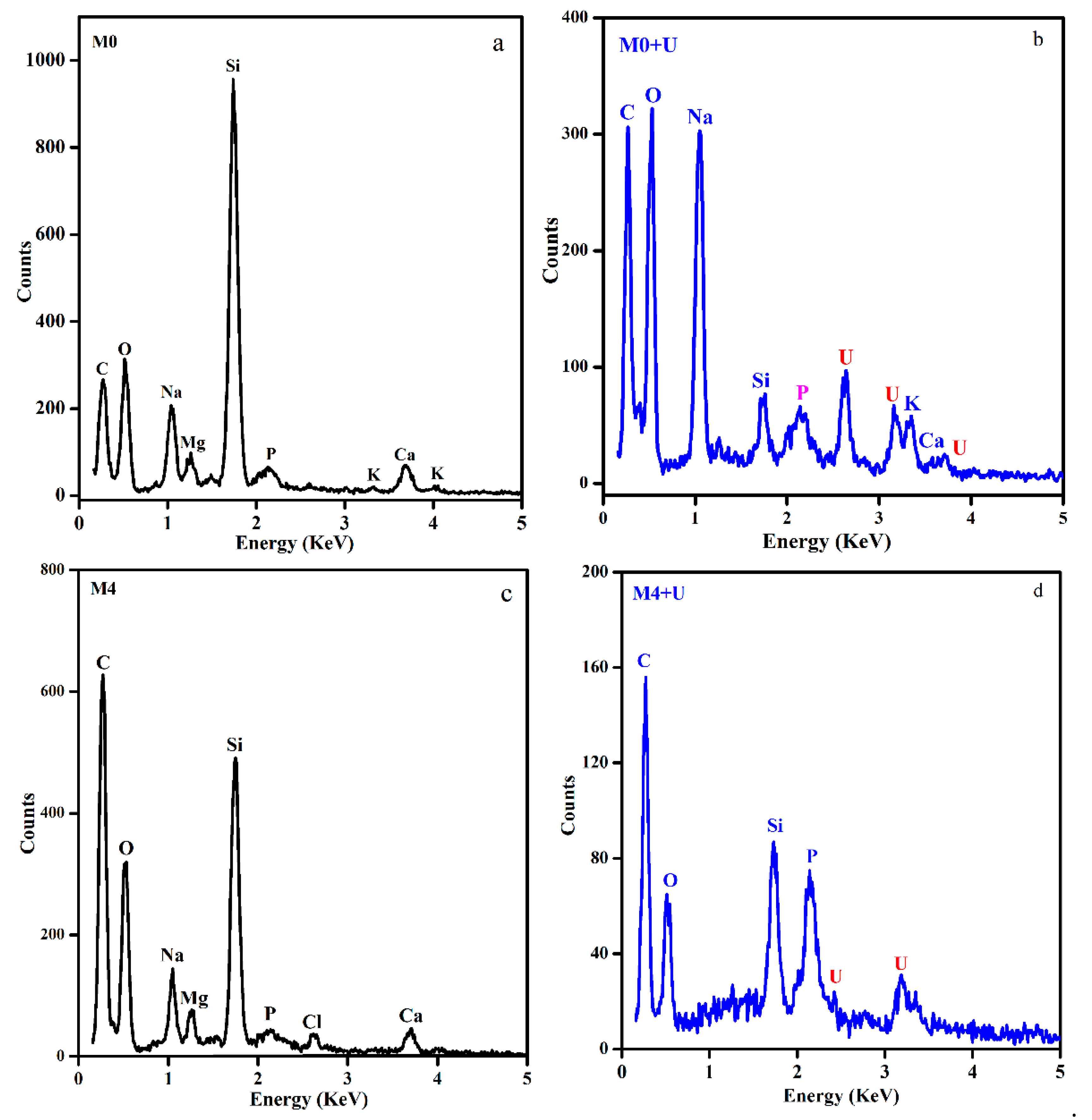
3.5. SEM-EDS and Spectroscopy Results of the Biomass after Uranium Biosorption
4. Discussion
4.1. Microscopy and Spectroscopy Observations of the Raw and Chemically Modified Biomass
4.2. Uranium Removal Influenced by Time
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nie, X.; Dong, F.; Bian, L.; Liu, M.; Ding, C.; He, H.; Yang, G.; Sun, S.; Qin, Y.; Huang, R. Uranium binding on Landoltia punctata as a result of formation of insoluble nano-U(VI) and U (IV) phosphate minerals. ACS Sustain. Chem. Eng. 2017, 5, 1494–1502. [Google Scholar] [CrossRef]
- Liu, M.; Dong, F.; Zhang, W.; Nie, X.; Wei, H.; Sun, S.; Zhong, X.; Liu, Y.; Wang, D. Contribution of surface functional groups and interface interaction to biosorption of strontium ions by Saccharomyces cerevisiae under culture conditions. RSC Adv. 2017, 7, 50880–50888. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Castro, I.; Martinez-Rodriguez, P.; Jroundi, F.; Solari, P.L.; Descostes, M.; Merroun, M.L. High-efficient microbial immobilization of solved U(VI) by the Stenotrophomonas strain Br 8. Water Res. 2020, 183, 116110. [Google Scholar] [CrossRef]
- Wang, Y.; Frutschi, M.; Suvorova, E.; Phrommavanh, V.; Descostes, M.; Osman, A.A.A.; Geipel, G.; Bernier-Latmani, R. Mobile uranium(IV)-bearing colloids in a mining-impacted wetland. Nat. Commun. 2013, 4, 2942–2947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Kelly, S.; Kemner, K.; Banfield, J.F. Nanometre-size products of uranium bioreduction. Nature 2002, 419, 134. [Google Scholar] [CrossRef]
- Tang, H.; Cheng, W.; Yi, Y.; Ding, C.; Nie, X. Nano zero valent iron encapsulated in graphene oxide for reducing uranium. Chemosphere 2021, 278, 130229. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Ballal, A.; Apte, S.K. Bioprecipitation of uranium from alkaline waste solutions using recombinant Deinococcus radiodurans. J. Hazard. Mater. 2013, 262, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.; Liu, Y.H.; Cao, X.H.; Li, M.; Huang, G.-L.; Hua, R.; Wang, C.-X.; Liu, Y.-T.; An, X.-F. Biosorption of uranium (VI) from aqueous solution by dead fungal biomass of Penicillium citrinum. Chem. Eng. J. 2011, 170, 1–6. [Google Scholar] [CrossRef]
- Daboor, S.M. Application of bacterial biomass as a potential heavy metal bio-removal agent. Afr. J. Microbiol. Res. 2014, 8, 2229–2237. [Google Scholar]
- Nie, X.; Dong, F.; Liu, N.; Liu, M.; Zhang, D.; Kang, W.; Sun, S.; Zhang, W.; Yang, J. Subcellular distribution of uranium in the roots of Spirodela punctata and surface interactions. Appl. Surf. Sci. 2015, 347, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Nie, X.; Ding, C.; Huang, R.; Dong, F.; Sun, S.; Qin, Y.; Liu, M. Kinetics and pH-dependent uranium bioprecipitation by Shewanella putrefaciens under aerobic conditions. J. Radioanal. Nucl. Chem. 2017, 312, 531–541. [Google Scholar] [CrossRef]
- Alessi, D.S.; Lezama-Pacheco, J.S.; Stubbs, J.E.; Janousch, M.; Bargar, J.R.; Persson, P.; Bernier-Latmani, R. The product of microbial uranium reduction includes multiple species with U (IV)–phosphate coordination. Geochim. Cosmochim. Acta 2014, 131, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Dong, F.; Liu, M.; He, H.; Sun, S.; Bian, L.; Yang, G.; Zhang, W.; Qin, Y.; Huang, R. Microbially mediated stable uranium phosphate nano-biominerals. J. Nanosci. Nanotechnol. 2017, 17, 6771–6780. [Google Scholar] [CrossRef]
- Lange, C.C.; Wackett, L.P.; Minton, K.W.; Daly, M.J. Engineering a recombinant Deinococcus radiodurans for organopollutant degradation in radioactive mixed waste environments. Nat. Biotechnol. 1998, 16, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Fredrickson, J.K.; Kostandarithes, H.M.; Li, S.W.; Plymale, A.E.; Daly, M.J. Reduction of Fe (III), Cr (VI), U(VI), and Tc (VII) by Deinococcus radiodurans R1. Appl. Environ. Microbiol. 2000, 66, 2006–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier-Latmani, R.; Veeramani, H.; Vecchia, E.D.; Junier, P.; Lezama-Pacheco, J.S.; Suvorova, E.I.; Sharp, J.O.; Wigginton, N.S.; Bargar, J.R. Non-uraninite products of microbial U(VI) reduction. Environ. Sci. Technol. 2010, 44, 9456–9462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plathe, K.L.; Lee, S.W.; Tebo, B.M.; Bargar, J.R.; Bernier-Latmani, R. Impact of microbial Mn oxidation on the remobilization of bioreduced U (IV). Environ. Sci. Technol. 2013, 47, 3606–3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stylo, M.; Alessi, D.S.; Shao, P.P.Y.; Lezama-Pacheco, J.S.; Bargar, J.R.; Bernier-Latmani, R. Biogeochemical controls on the product of microbial U(VI) reduction. Environ. Sci. Technol. 2013, 47, 12351–12358. [Google Scholar] [CrossRef] [Green Version]
- Stylo, M.; Neubert, N.; Roebbert, Y.; Weyer, S.; Bernier-Latmani, R. Mechanism of uranium reduction and immobilization in Desulfovibrio vulgaris biofilms. Environ. Sci. Technol. 2015, 49, 10553–10561. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Sar, P. Uranium biomineralization by a metal resistant Pseudomonas aeruginosa strain isolated from contaminated mine waste. J. Hazard. Mater. 2011, 186, 336–343. [Google Scholar] [CrossRef]
- Liang, X.; Csetenyi, L.; Gadd, G.M. Uranium bioprecipitation mediated by yeasts utilizing organic phosphorus substrates Appl. Microbiol. Biotechnol. 2016, 100, 5141–5151. [Google Scholar] [CrossRef]
- Bai, R.S.; Abraham, T.E. Studies on enhancement of Cr (VI) biosorption by chemically modified biomass of Rhizopus nigricans. Water Res. 2002, 36, 1224–1236. [Google Scholar] [CrossRef]
- Kapoor, A.; Viraraghavan, T. Heavy metal biosorption sites in Aspergillus niger. Bioresour. Technol. 1997, 61, 221–227. [Google Scholar] [CrossRef]
- Holan, Z.R.; Volesky, B. Accumulation of cadmium, lead, and nickel by fungal and wood biosorbents. Appl. Biochem. Biotechnol. 1995, 53, 133–146. [Google Scholar] [CrossRef]
- Tran, H.N.; You, S.-J.; Hosseini-Bandegharaei, A.; Chao, H.-P. Mistakes and inconsistencies regarding adsorption of contaminants from aqueous solutions: A critical review. Water Res. 2017, 120, 88–116. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, S.; Wang, J. Poly Amidoxime functionalized carbon nanotube as an efficient adsorbent for removal of uranium from aqueous solution. J. Mol. Liq. 2020, 319, 114288. [Google Scholar] [CrossRef]
- Zhong, X.; Liang, W.; Lu, Z.; Qui, M.; Hu, B. Ultra-high capacity of graphene oxide conjugated covalent organic framework nanohybrid for U(VI) and Eu (III) adsorption removal. J. Mol. Liq. 2021, 323, 114603. [Google Scholar] [CrossRef]
- Farci, D.; Bowler, M.W.; Kirkpatrick, J.; McSweeney, S.; Tramontano, E.; Piano, D. New features of the cell wall of the radio-resistant bacterium Deinococcus radiodurans. Biochim. Biophys. Acta Biomembr. 2014, 183, 1978–1984. [Google Scholar] [CrossRef]
- Müller, D.J.; Baumeister, W.; Engel, A. Conformational change of the hexagonally packed intermediate layer of Deinococcus radiodurans monitored by atomic force microscopy. J. Bacteriol. Res. 1996, 17, 3025–3030. [Google Scholar] [CrossRef] [Green Version]
- Baumeister, W.; Barth, M.; Hegerl, R.; Guckenberger, R.; Hahn, M.; Saxton, W.O. Three-dimensional structure of the regular surface layer (HPI layer) of Deinococcus radiodurans. J. Mol. Biol. 1986, 18, 241–250. [Google Scholar] [CrossRef]
- Work, E. Chemical Structure of Bacterial Cell Walls: Amino-acids of Walls of Micrococcus radiodurans. Nature 1964, 201, 1107–1109. [Google Scholar] [CrossRef]
- Knivett, V.A.; Cullen, J.; Jackson, M.J. Odd-numbered fatty acids in Micrococcus radiodurans. Biochemistry 1965, 96, 2C–3C. [Google Scholar] [CrossRef] [Green Version]
- Rothfuss, H.; Lara, J.C.; Schmid, A.K.; Lidstrom, M.E. Involvement of the S-layer proteins Hpi and SlpA in the maintenance of cell envelope integrity in Deinococcus radiodurans R1. Microbiology 2006, 152, 2779–2787. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.S.; McKay, G. Pseudo-second order model for sorption processes. Process Biochem. 1999, 34, 451–465. [Google Scholar] [CrossRef]
- Chen, J.P.; Yang, L. Chemical modification of Sargassum sp. for prevention of organic leaching and enhancement of uptake during metal biosorption. Ind. Eng. Chem. Res. 2005, 44, 9931–9942. [Google Scholar]
- Čejka, J. 12. Infrared Spectroscopy and Thermal Analysis of the Uranyl Minerals. In Uranium; De Gruyter: Berlin, Germany, 2018; pp. 521–622. [Google Scholar]
- Li, X.; Ding, C.; Liao, J.; Lan, T. Biosorption of uranium on Bacillus sp. dwc-2: Preliminary investigation on mechanism. J. Environ. Radioact. 2014, 135, 6–12. [Google Scholar] [CrossRef]
- Yang, L.; Chen, J.P. Biosorption of hexavalent chromium onto raw and chemically modified Sargassum sp. Bioresour. Technol. 2008, 99, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Dong, F.; Liu, N. An investigation on the subcellular distribution and compartmentalization of uranium in Phaseolus vulgaris L. J. Radioanal. Nucl. Chem. 2014, 299, 1351–1357. [Google Scholar] [CrossRef]
- Huang, G.; Ng, T.W.; An, T.; Li, G.; Xia, D.; Yip, H.Y.; Zao, Y.; Wong, P.K. Probing the intracellular organic matters released from the photocatalytic inactivation of bacteria using fractionation procedure and excitation-emission-matrix fluorescence. Water Res. 2017, 110, 270–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.Y.; Ohnuki, T.; Tanaka, K.; Kozai, N.; Kamiishi, E.; Utsunomiya, S. Post-adsorption process of Yb phosphate nano-particle formation by Saccharomyces cerevisiae. Geochim. Acta 2012, 93, 30–46. [Google Scholar] [CrossRef]
- Misra, C.S.; Appukuttan, D.; Kantamreddi, V.S.S.; Rao, S.A.; Apte, S.K. Recombinant D. radiodurans cells for bioremediation of heavy metals from acidic/neutral aqueous wastes. Bioeng. Bugs 2012, 3, 44–48. [Google Scholar] [PubMed] [Green Version]
- Park, D.; Yun, Y.S.; Park, J.M. Studies on hexavalent chromium biosorption by chemically-treated biomass of Ecklonia sp. Chemosphere 2005, 60, 1356–1364. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Wave Number/cm−1 | Assignment |
|---|---|---|
| 1 | 3600 | Hydroxyl groups of the glucose and the -NH stretching |
| 2 | 2930 | C-H stretching |
| 3 | 1660 | C=O stretching, amide I band of amide bond |
| 4 | 1540 | Amide II band |
| 5 | 1450 | Symmetric bending of CH3 of the acetyl moiety |
| 6 | 1390 | C-O stretching of COOH |
| 7 | 1240 | Amide III band, δO-H of COOH |
| 8 | 1070 | Phosphate group |
| 9 | 1030 | C-OH of carbohydrate |
| 10 | 920 | Asymmetric stretching vibration of v3 UO22+ and stretching vibration of weekly bonded oxygen ligands with uranium (v U-Oligand) |
| Biomass | Pseudo-First-Order | Pseudo-Second-Order | Intraparticle Diffusion Kinetics | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| k1 | qe(cal) | R2 | k2 × 10−3 | qe(cal) | h | R2 | k3 | C | R2 | |
| (min−1) | (mg/g) | (g/(mg·min)) | (mg/g) | (mg/(g·min)) | (g/(mg·min−0.5)) | (mg/g) | ||||
| M0 | 0.017 | 134.2 | 0.6491 | 1.08 | 159.0 | 2.60 | 0.9949 | 2.95 | 55.2 | 0.9263 |
| M1 | 0.323 | 182.1 | 0.2203 | 18.77 | 191.6 | 43.46 | 0.9998 | 1.01 | 162.7 | 0.4032 |
| M2 | 0.018 | 137.2 | 0.8897 | 1.28 | 155.5 | 3.14 | 0.9982 | 3.06 | 53.4 | 0.7760 |
| M3 | 0.103 | 114.3 | 0.6397 | 4.95 | 127.7 | 8.29 | 0.9997 | 1.44 | 82.7 | 0.6710 |
| M4 | 0.066 | 177.0 | 0.6225 | 5.52 | 186.9 | 19.21 | 0.9998 | 2.10 | 125.1 | 0.5197 |
| M5 | 0.018 | 164.1 | 0.9253 | 1.27 | 182.1 | 4.01 | 0.9993 | 3.64 | 63.4 | 0.7323 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, X.; Dong, F.; Liu, M.; Cheng, W.; Ding, C.; Bian, L.; Sun, S. Bonding Behavior and Mechanism of U(VI) by Chemically Modified Deinococcus radiodurans. Minerals 2021, 11, 1108. https://doi.org/10.3390/min11101108
Nie X, Dong F, Liu M, Cheng W, Ding C, Bian L, Sun S. Bonding Behavior and Mechanism of U(VI) by Chemically Modified Deinococcus radiodurans. Minerals. 2021; 11(10):1108. https://doi.org/10.3390/min11101108
Chicago/Turabian StyleNie, Xiaoqin, Faqin Dong, Mingxue Liu, Wencai Cheng, Congcong Ding, Liang Bian, and Shiyong Sun. 2021. "Bonding Behavior and Mechanism of U(VI) by Chemically Modified Deinococcus radiodurans" Minerals 11, no. 10: 1108. https://doi.org/10.3390/min11101108
APA StyleNie, X., Dong, F., Liu, M., Cheng, W., Ding, C., Bian, L., & Sun, S. (2021). Bonding Behavior and Mechanism of U(VI) by Chemically Modified Deinococcus radiodurans. Minerals, 11(10), 1108. https://doi.org/10.3390/min11101108