
Stimulation of Biomethane Productivity in Anaerobic Digestion Using Electro-Conductive Carbon-Nanotube Hollow-Fiber Media




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Seed Sludge and Feedstock
2.2. Preparation of Carbon-Nanotube Hollow-Fiber Media and Polymeric Hollow Fiber Media
2.3. Experiments
2.4. Sample Analysis
2.5. Microbial Community Analysis
3. Results and Discussion
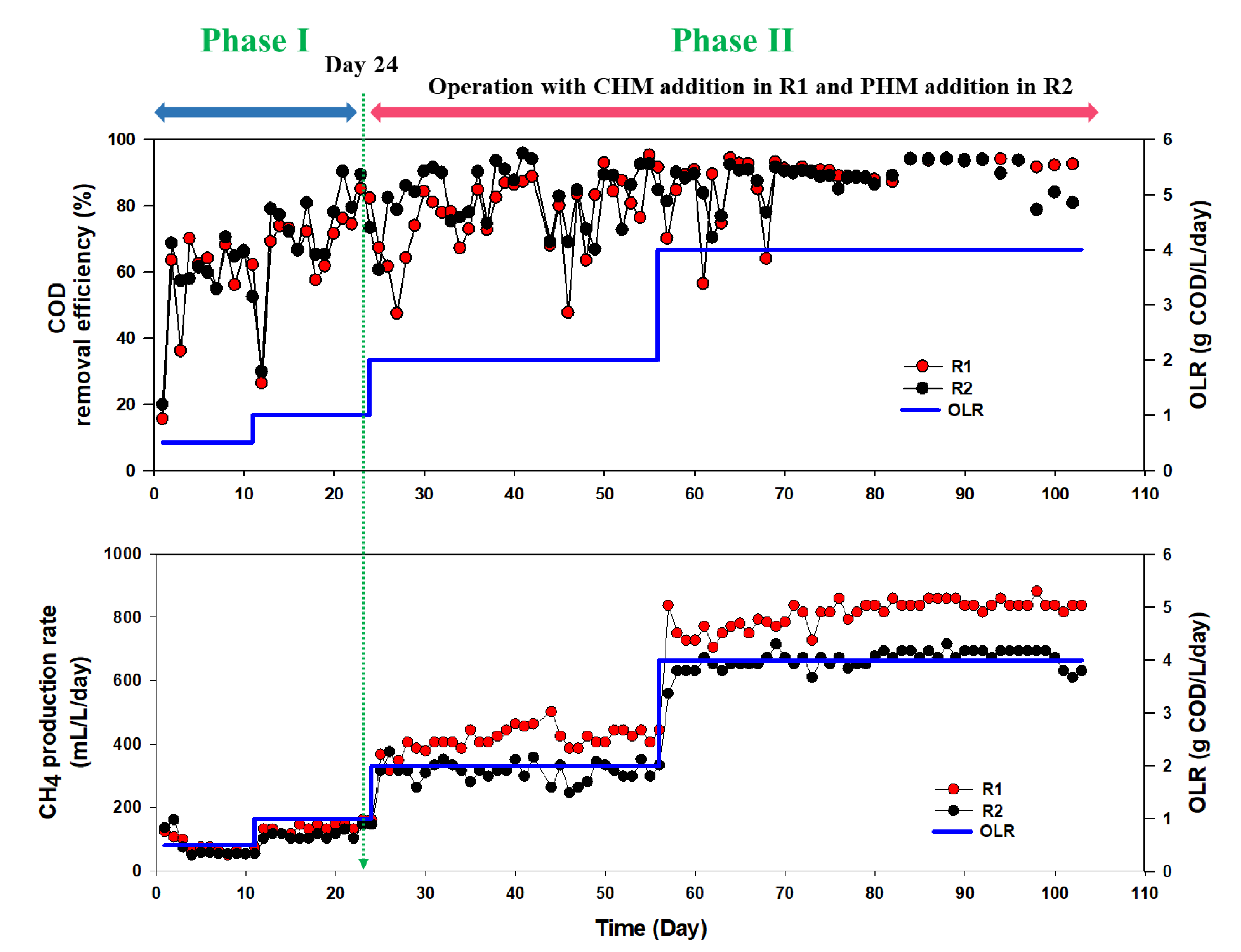
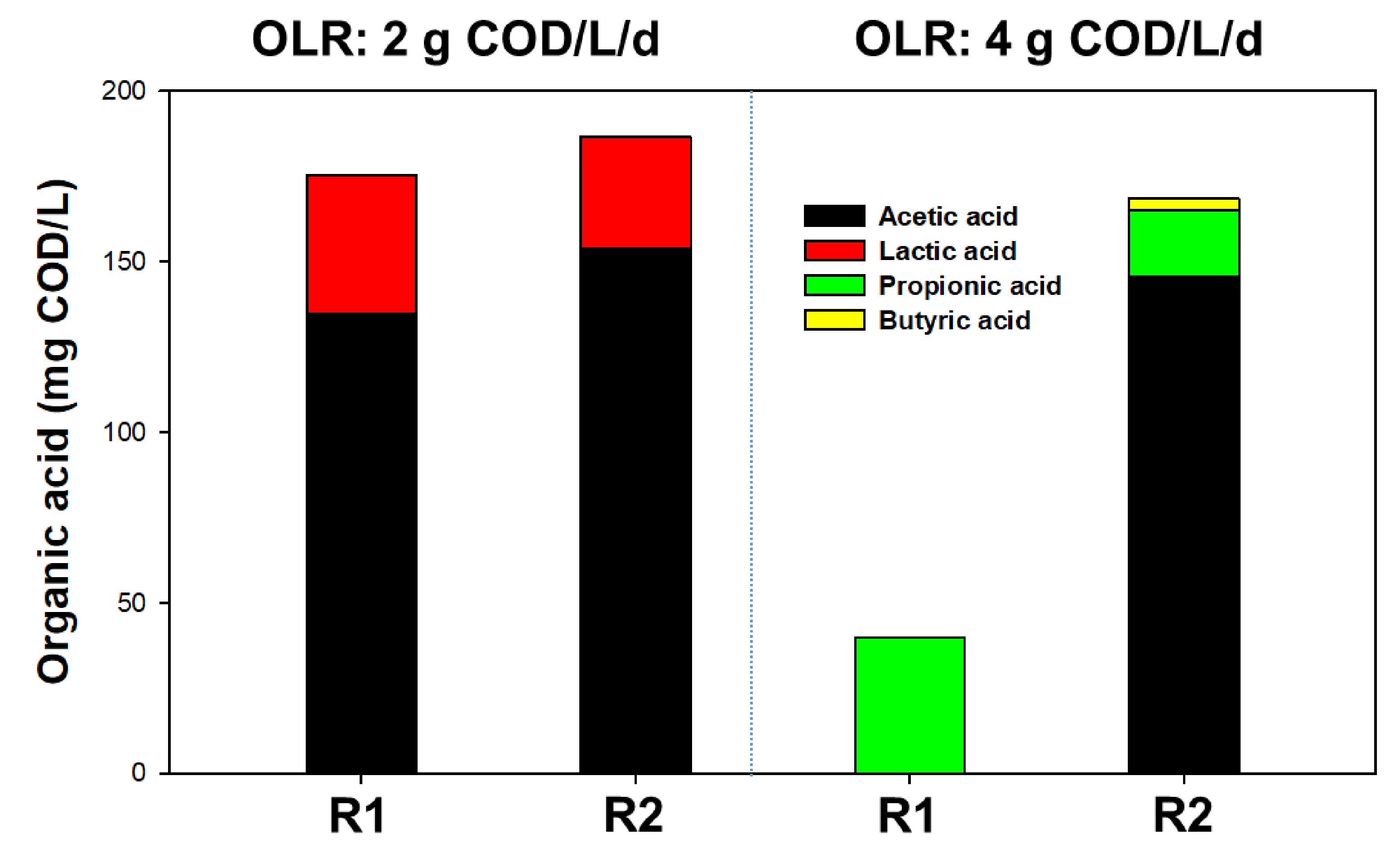
3.1. Effect of CHM on AD Performance
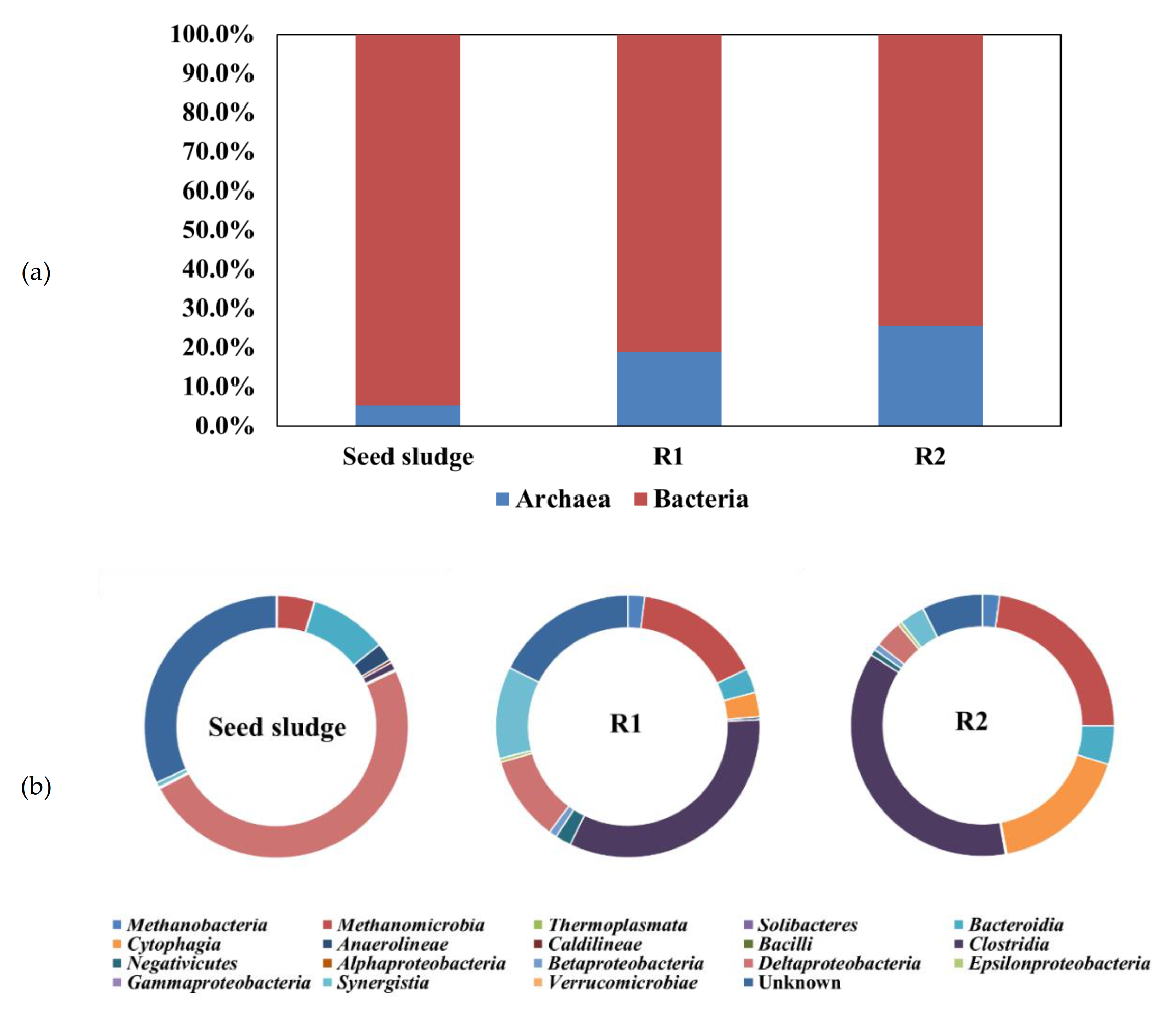
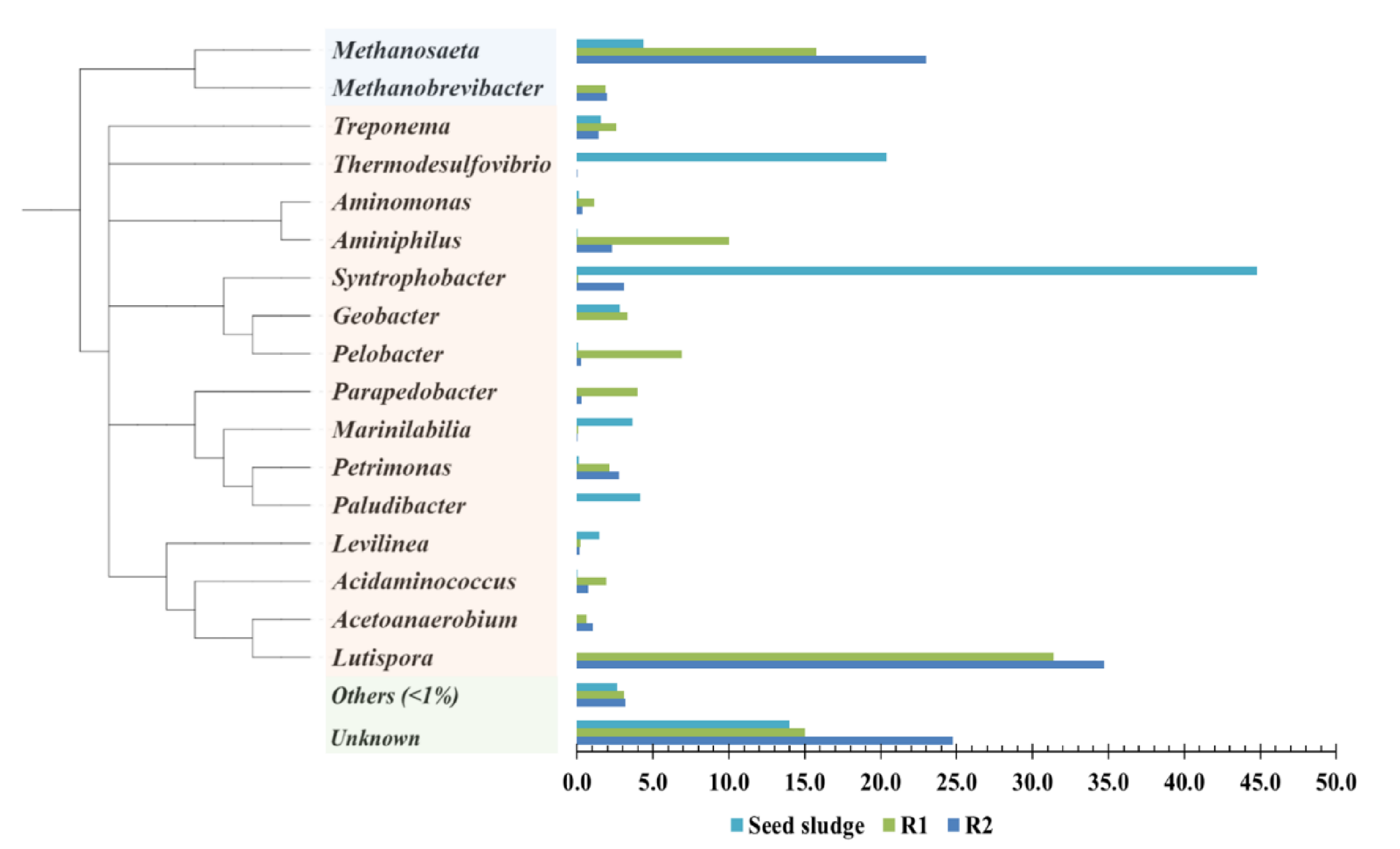
3.2. Change in Taxonomic Distribution of the Microorganisms
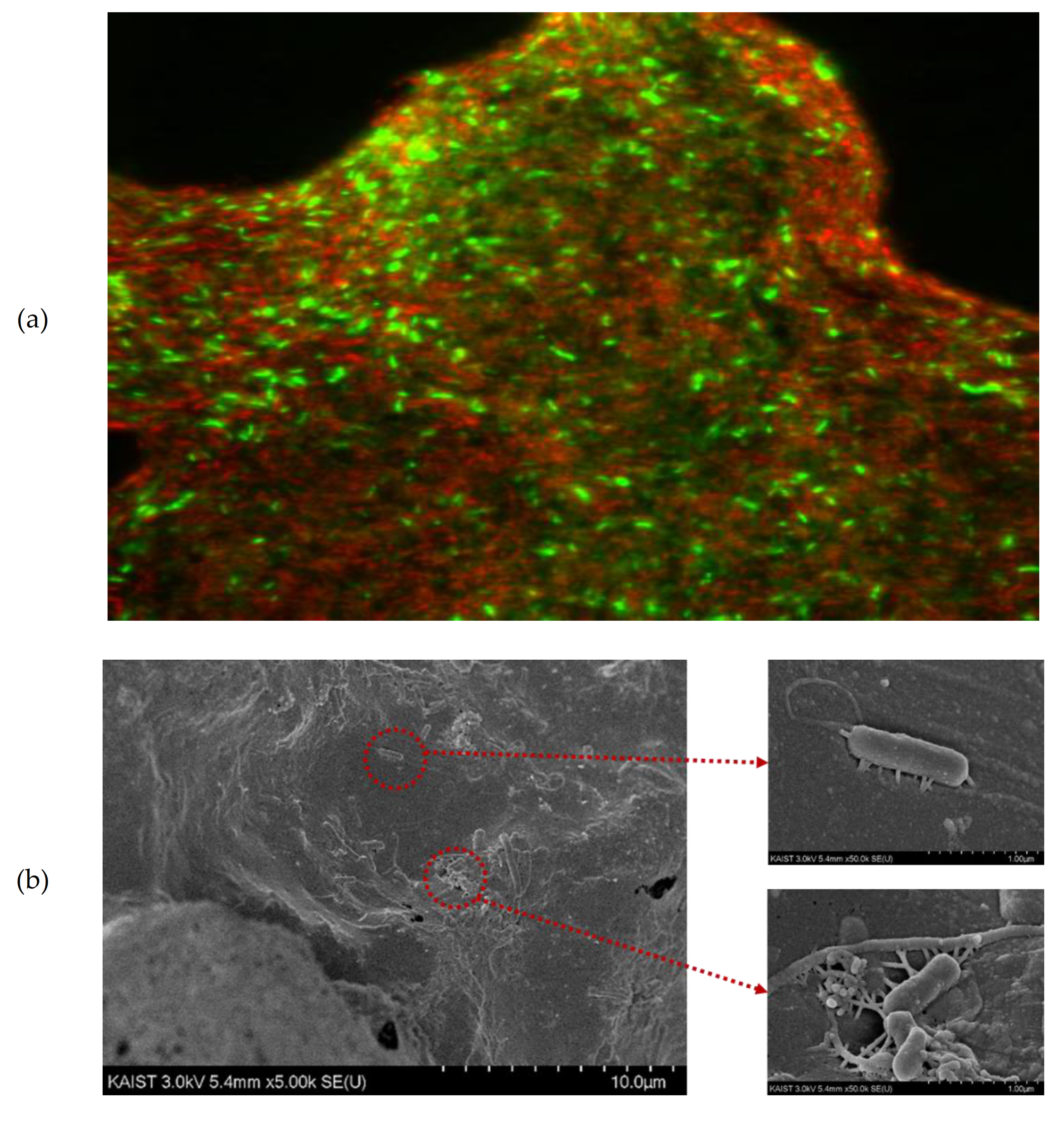
3.3. Effect of CHM on Direct Interspecies Electron Transfer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Nomenclature
| Full Name | Abbreviation | Unit |
| Anaerobic digestion | AD | |
| Carbon-nanotube hollow-fiber media | CHM | |
| Chemical oxygen demand | COD | mg COD/L |
| Confocal laser scanning microscopy | CLSM | |
| Direct interspecies electron transfer | DIET | |
| interspecies electron transfer | IET | |
| Methane production rate | MPR | |
| Operational taxonomic units | OTUs | |
| Organic loading rate | OLR | g COD/L/day |
| Polymeric hollow-fiber media | PHM |
References
- World Biogas Association. Available online: https://www.worldbiogasassociation.org/wp-content/uploads/2019/07/WBA-globalreport-56ppa4_digital.pdf (accessed on 27 December 2020).
- Zhang, P.; Yan, F.; Du, C. A comprehensive analysis of energy management strategies for hybrid electric vehicles based on bibliometrics. Renew. Sust. Energ. Rev. 2015, 48, 88–104. [Google Scholar] [CrossRef]
- Available online: https://www.fortunebusinessinsights.com/industry-reports/biogas-market-100910 (accessed on 27 December 2020).
- Rajagopal, R.; Massé, D.I.; Singh, G. A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresour. Technol. 2013, 143, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Sposob, M.; Moon, H.S.; Lee, D.; Kim, T.H.; Yun, Y.M. Comprehensive analysis of the microbial communities and operational parameters of two full-scale anaerobic digestion plants treating food waste in South Korea: Seasonal variation and effect of ammonia. J. Hazard. Mater. 2020, 398, 122975. [Google Scholar] [CrossRef]
- Baek, G.; Jung, H.; Kim, J.; Lee, C. A long-term study on the effect of magnetite supplementation in continuous anaerobic digestion of dairy effluent–magnetic separation and recycling of magnetite. Bioresour. Technol. 2017, 241, 830–840. [Google Scholar] [CrossRef]
- Baek, G.; Shi, L.; Rossi, R.; Logan, B.E. The effect of high applied voltages on bioanodes of microbial electrolysis cells in the presence of chlorides. Chem. Eng. J. 2021, 405, 126742. [Google Scholar] [CrossRef]
- Chen, S.; Rotaru, A.-E.; Shrestha, P.M.; Malvankar, N.S.; Liu, F.; Fan, W.; Nevin, K.P.; Lovley, D.R. Promoting Interspecies Electron Transfer with Biochar. Sci. Rep. 2014, 5019. [Google Scholar] [CrossRef] [Green Version]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct Exchange of Electrons Within Aggregates of an Evolved Syntrophic Coculture of Anaerobic Bacteria. Science 2010, 330, 1413–1415. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, Y.; Holmes, D.E.; Dang, Y.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Potential enhancement of direct interspecies electron transfer for syntrophic metabolism of propionate and butyrate with biochar in up-flow anaerobic sludge blanket reactors. Bioresour. Technol. 2016, 209, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Yee, M.O.; Rotaru, A.E. Extracellular electron uptake in Methanosarcinales is independent of multiheme c-type cytochromes. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. USA 2012, 109, 10042–10046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-Y.; Lee, S.-H.; Park, H.-D. Enrichment of specific electroactive microorganisms and enhancement of methane production by adding granular activated carbon in anaerobic reactors. Bioresour. Technol. 2016, 205, 205–212. [Google Scholar] [CrossRef]
- Lekawa-Raus, A.; Patmore, J.; Kurzepa, L.; Bulmer, J.; Koziol, K. Electrical Properties of Carbon Nanotube Based Fibers and Their Future Use in Electrical Wiring. Adv. Funct. Mater. 2014, 24, 3661–3682. [Google Scholar] [CrossRef]
- Wei, G.; Yu, H.; Quan, X.; Chen, S.; Zhao, H.; Fan, X. Constructing All Carbon Nanotube Hollow Fiber Membranes with Improved Performance in Separation and Antifouling for Water Treatment. Environ. Sci. Technol. 2014, 48, 8062–8068. [Google Scholar] [CrossRef]
- Matsumoto, K.; Uejima, H.; Iwasaki, T.; Sano, Y.; Sumino, H. Studies on regenerated protein fibers. III. Production of regenerated silk fibroin fiber by the self-dialyzing wet spinning method. J. Appl. Polym. Sci. 1996, 60, 503–511. [Google Scholar] [CrossRef]
- Gilcreas, F.W. Standard methods for the examination of water and waste water. Am. J. Public Health Nation’s Health 1966, 56, 387–388. [Google Scholar] [CrossRef]
- Viggi, C.C.; Rossetti, S.; Fazi, S.; Paiano, P.; Majone, M.; Aulenta, F. Magnetite Particles Triggering a Faster and More Robust Syntrophic Pathway of Methanogenic Propionate Degradation. Environ. Sci. Technol. 2014, 48, 7536–7543. [Google Scholar] [CrossRef]
- Yun, Y.M.; Sung, S.; Shin, H.S.; Han, J.I.; Kim, H.W.; Kim, D.H. Producing desulfurized biogas through removal of sulfate in the first-stage of a two-stage anaerobic digestion. Biotechnol. Bioeng. 2017, 114, 970–979. [Google Scholar] [CrossRef]
- Regueiro, L.; Veiga, P.; Figueroa, M.; Alonso-Gutierrez, J.; Stams, A.J.M.; Lema, J.M.; Carballa, M. Relationship between microbial activity and microbial community structure in six full-scale anaerobic digesters. Microbiol. Res. 2012, 167, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, F.R.; Dinsdale, R.; Hawkes, D.L.; Hussy, I. Sustainable fermentative hydrogen production: Challenges for process optimisation. Int. J. Hydrog. Energy 2002, 27, 1339–1347. [Google Scholar] [CrossRef]
- Xia, Y.; Massé, D.I.; McAllister, T.A.; Kong, Y.; Seviour, R.; Beaulieu, C. Identity and diversity of archaeal communities during anaerobic co-digestion of chicken feathers and other animal wastes. Bioresour. Technol. 2012, 110, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Du, H.; Ren, C.; Xu, Y. Illuminating Anaerobic Microbial Community and Cooccurrence Patterns across a Quality Gradient in Chinese Liquor Fermentation Pit Muds. Appl. Environ. Microbiol. 2016, 82, 2506–2515. [Google Scholar] [CrossRef] [Green Version]
- Ledezma, P.; Lu, Y.; Freguia, S. Electroactive haloalkaliphiles exhibit exceptional tolerance to free ammonia. FEMS Microbiol. Lett. 2017, 365, fnx260. [Google Scholar] [CrossRef] [PubMed]
- Moscoviz, R.; de Fouchécour, F.; Santa-Catalina, G.; Bernet, N.; Trably, E. Cooperative growth of Geobacter sulfurreducens and Clostridium pasteurianum with subsequent metabolic shift in glycerol fermentation. Sci. Rep. 2017, 7, 44334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador, A.F.; Martins, G.; Melle-Franco, M.; Serpa, R.; Stams, A.J.M.; Cavaleiro, A.J.; Pereira, M.A.; Alves, M.M. Carbon nanotubes accelerate methane production in pure cultures of methanogens and in a syntrophic coculture. Environ. Microbiol. 2017, 19, 2727–2739. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Holmes, D.E.; Zhao, Z.; Woodard, T.L.; Zhang, Y.; Sun, D.; Wang, L.-Y.; Nevin, K.P.; Lovley, D.R. Enhancing anaerobic digestion of complex organic waste with carbon-based conductive materials. Bioresour. Technol. 2016, 220, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.; Baena, S.; Fardeau, M.-L.; Patel, B.K.C. Aminiphilus circumscriptus gen. nov., sp. nov., an anaerobic amino-acid-degrading bacterium from an upflow anaerobic sludge reactor. Int. J. Syst. Evol. Microbiol. 2007, 57, 1914–1918. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Yang, Y.; Quan, X.; Zhao, Z. Potentially direct interspecies electron transfer of methanogenesis for syntrophic metabolism under sulfate reducing conditions with stainless steel. Bioresour. Technol. 2017, 234, 303–309. [Google Scholar] [CrossRef]
- Liu, F.; Rotaru, A.E.; Shrestha, P.M.; Malvankar, N.S.; Nevin, K.P.; Lovley, D.R. Promoting direct interspecies electron transfer with activated carbon. Energy Environ. Sci. 2012, 5, 8982–8989. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Mauter, M.S.; Elimelech, M. Physicochemical Determinants of Multiwalled Carbon Nanotube Bacterial Cytotoxicity. Environ. Sci. Technol. 2008, 42, 7528–7534. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Read Count | OTUs | Chao1 | Shannon |
|---|---|---|---|---|
| Seed sludge | 45,917 | 158 | 158.75 | 3.69 |
| R1 | 58,396 | 103 | 108.83 | 2.30 |
| R2 | 62,178 | 97 | 109.21 | 2.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Han, S.; Yun, Y.-M.; Kang, S. Stimulation of Biomethane Productivity in Anaerobic Digestion Using Electro-Conductive Carbon-Nanotube Hollow-Fiber Media. Minerals 2021, 11, 179. https://doi.org/10.3390/min11020179
Yang S, Han S, Yun Y-M, Kang S. Stimulation of Biomethane Productivity in Anaerobic Digestion Using Electro-Conductive Carbon-Nanotube Hollow-Fiber Media. Minerals. 2021; 11(2):179. https://doi.org/10.3390/min11020179
Chicago/Turabian StyleYang, Seongmin, Seungyeob Han, Yeo-Myeong Yun, and Seoktae Kang. 2021. "Stimulation of Biomethane Productivity in Anaerobic Digestion Using Electro-Conductive Carbon-Nanotube Hollow-Fiber Media" Minerals 11, no. 2: 179. https://doi.org/10.3390/min11020179
APA StyleYang, S., Han, S., Yun, Y. -M., & Kang, S. (2021). Stimulation of Biomethane Productivity in Anaerobic Digestion Using Electro-Conductive Carbon-Nanotube Hollow-Fiber Media. Minerals, 11(2), 179. https://doi.org/10.3390/min11020179