Arsenic (As) Removal Using Talaromyces sp. KM-31 Isolated from As-Contaminated Mine Soil



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fungal Isolation and Molecular Identification
2.3. Arsenic Tolerance Assay
2.4. Arsenic Biosorption Experiments
2.5. Arsenic Removal by Modified Fungal Biomass
2.6. Analytical Methods
3. Results and Discussion
3.1. Isolation and Identification of anAarsenic-Tolerant Fungal Strain
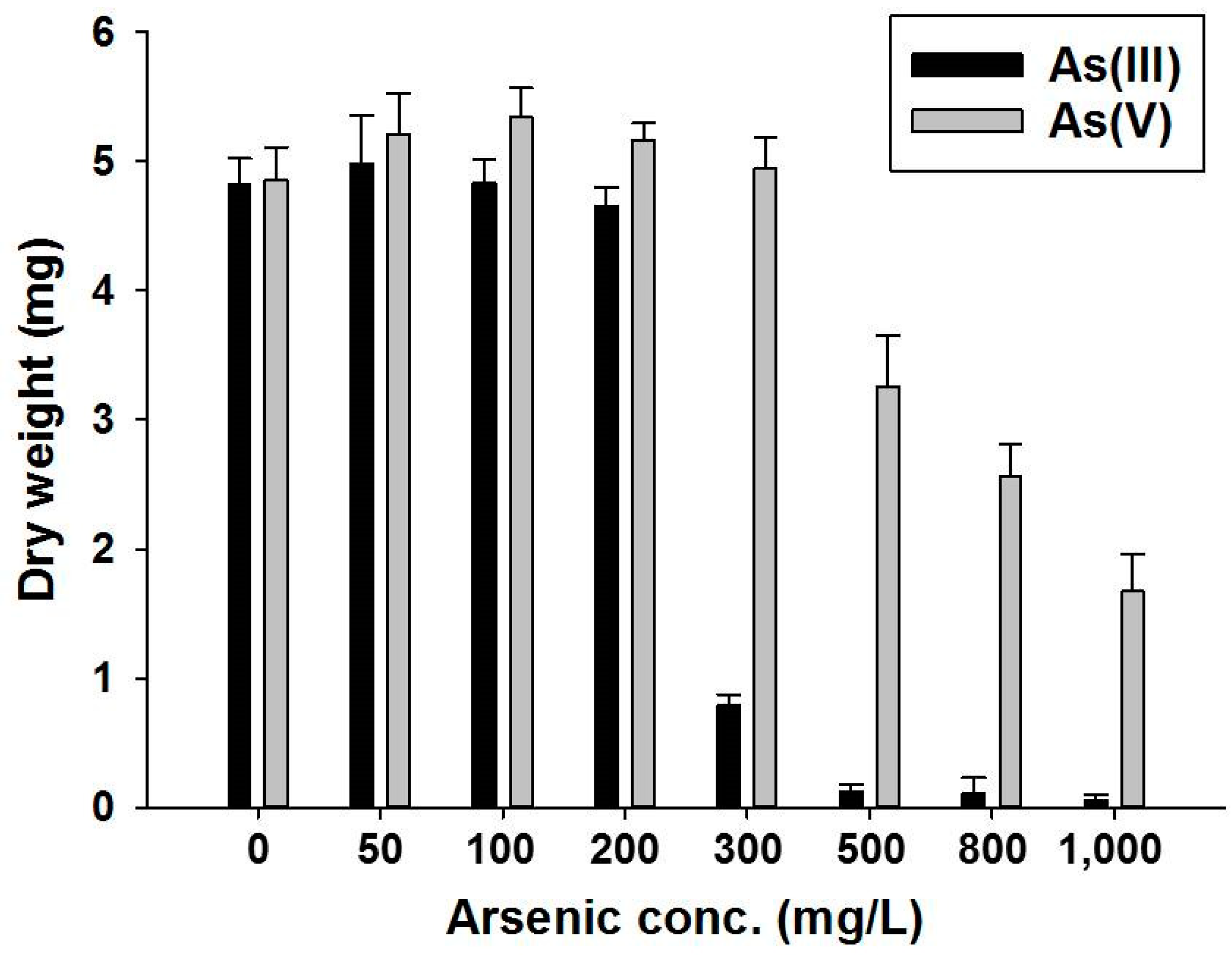
3.2. Growth and Arsenic Tolerance of Talaromyces sp. KM-31
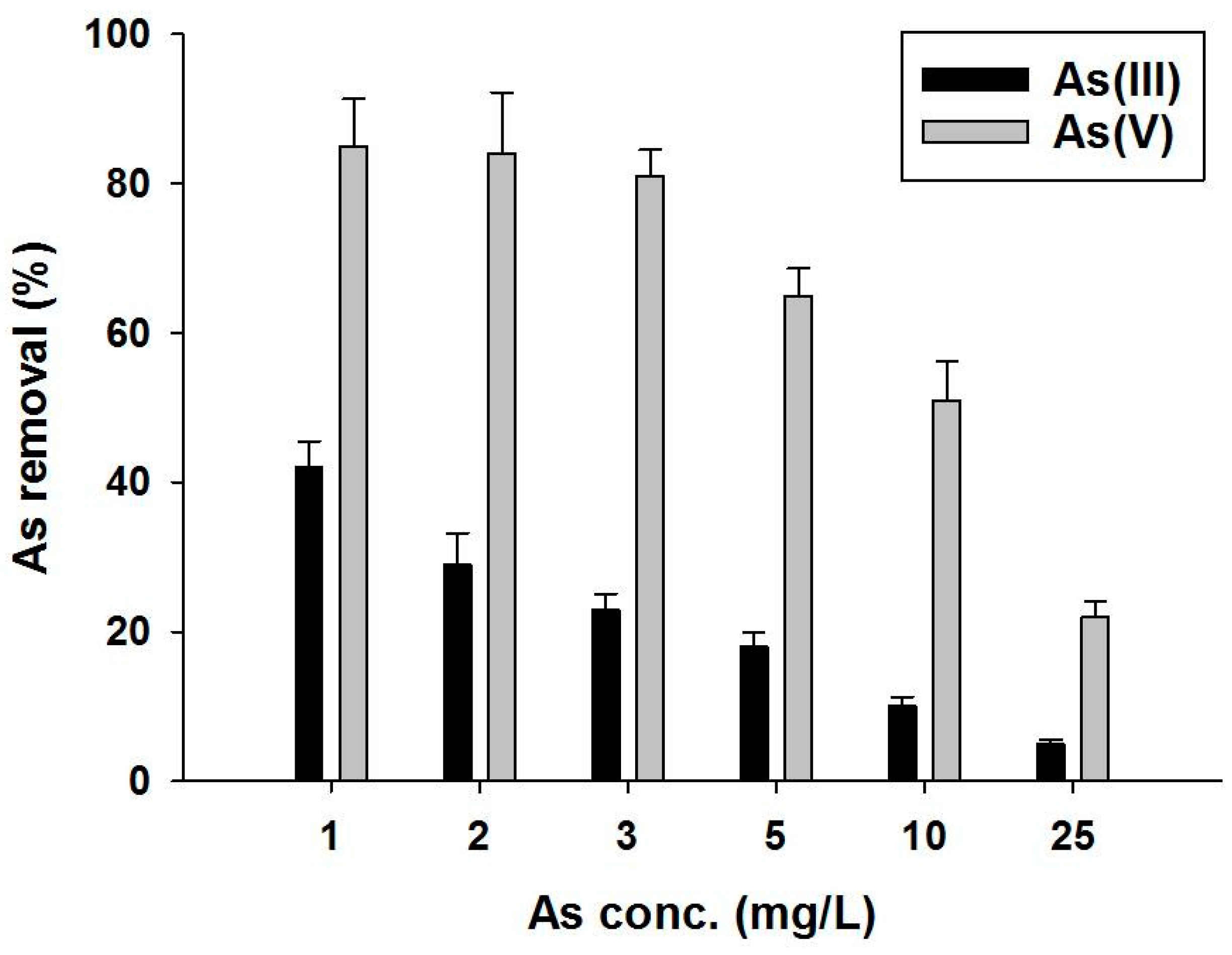
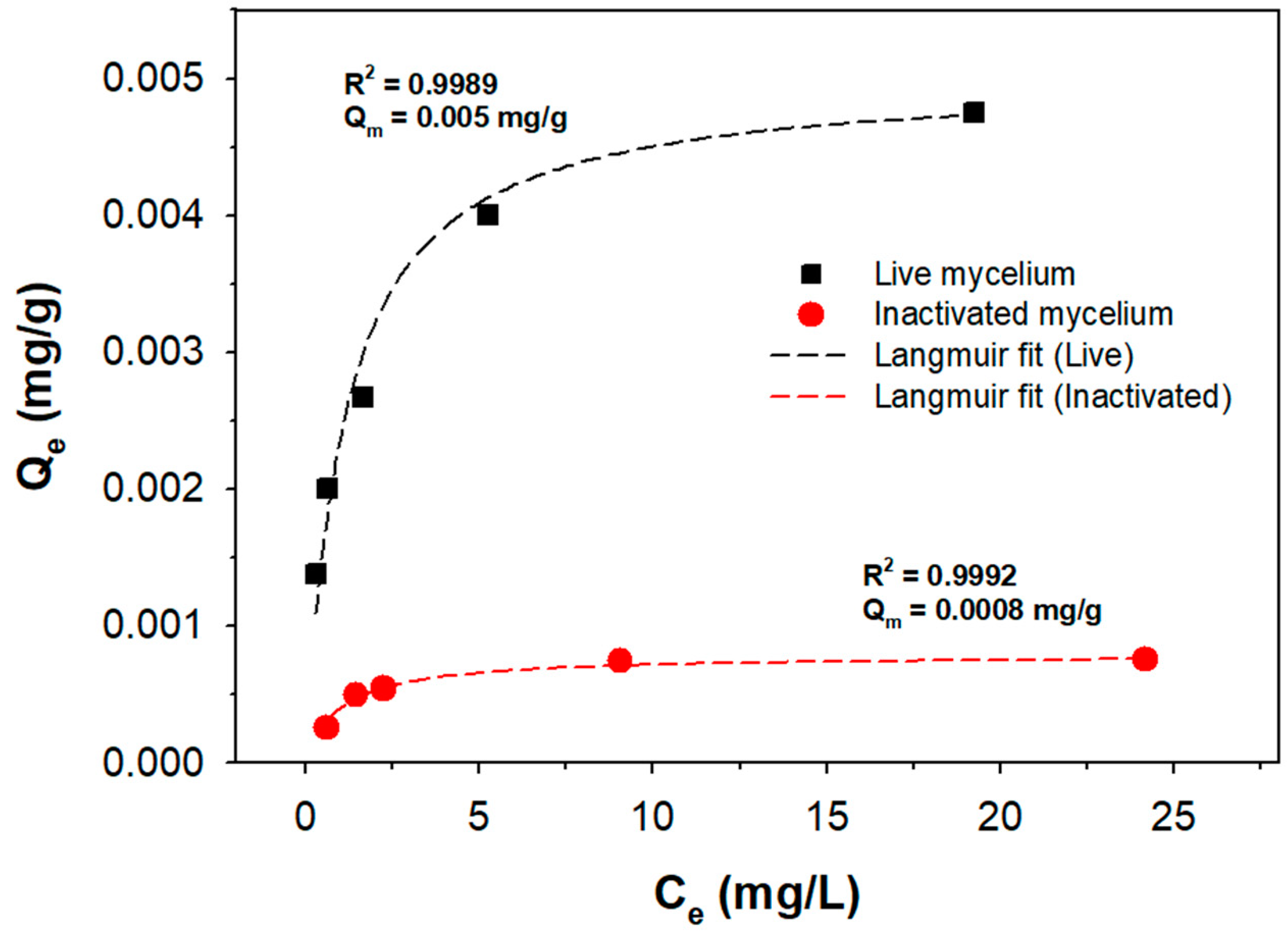
3.3. Arsenic Removal by Live Mycelial Pellets
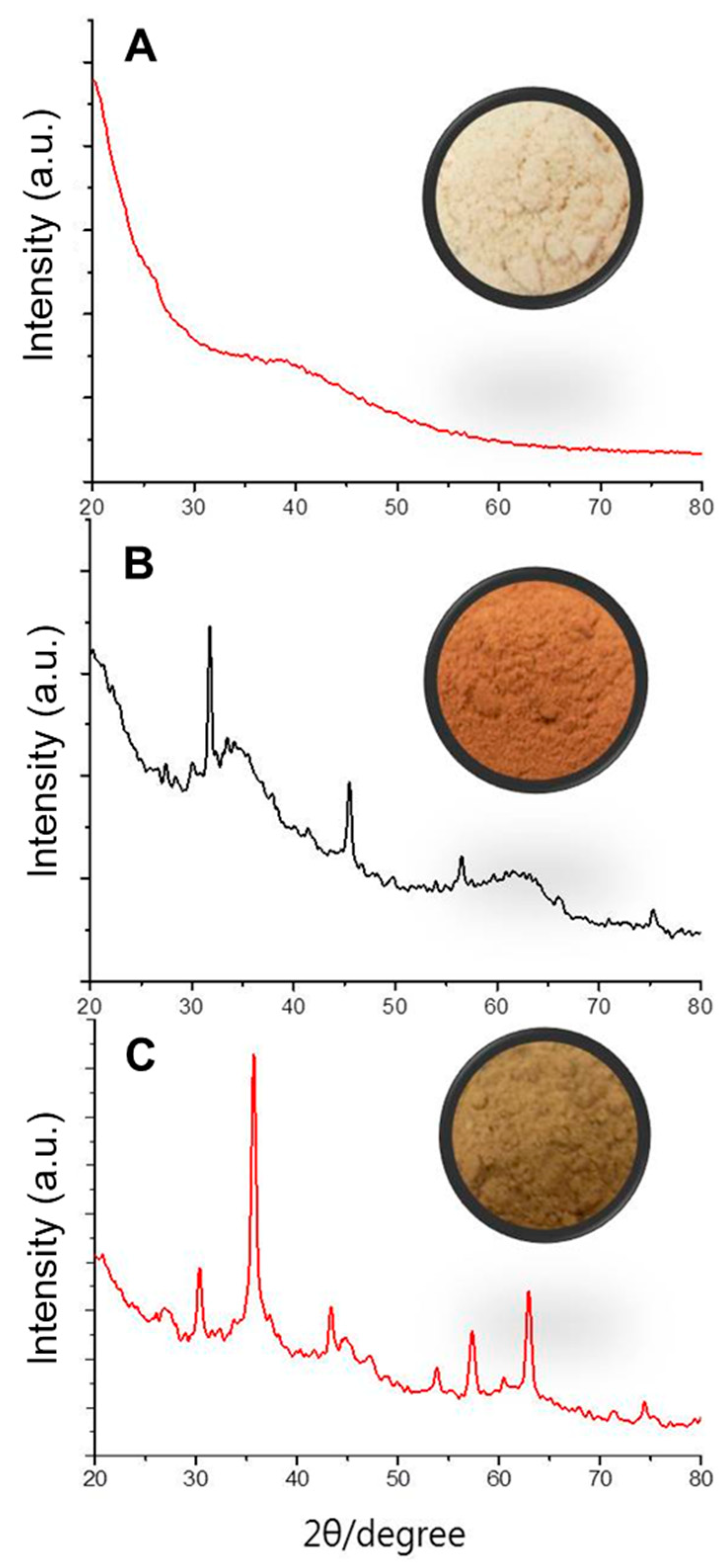
3.4. Arsenic Removal by Modified Biomass of Talaromyces sp. KM-31
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Donahoe-Christiansen, J.; D’Imperio, S.; Jackson, C.R.; Inskeep, W.P.; McDermott, T.R. Arsenite-oxidizing Hydrogenobaculum strain isolated from an acid-sulfate-chloride geothermal spring in Yellowstone National Park. Appl. Environ. Microbiol. 2004, 70, 1865–1868. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Mulligan, C.N. Occurrence of arsenic contamination in Canada: Sources, behavior and distribution. Sci. Total Environ. 2006, 366, 701–721. [Google Scholar] [CrossRef] [PubMed]
- Turpeinen, R.; Kallio, M.P.; Kairesalo, T. Role of microbes in controlling the speciation of arsenic and production of arsines in contaminated soils. Sci. Total Environ. 2002, 285, 133–145. [Google Scholar] [CrossRef]
- Tamaki, S.; Frankenberger, W.T. Environmental biochemistry of arsenic. Rev. Environ. Contam. Toxicol. 1992, 124, 79–110. [Google Scholar] [PubMed]
- Moreno-Jiménez, E.; Manzano, R.; Esteban, E.; Peñalosa, J. The fate of arsenic in soils adjacent to an old mine site (Bustarviejo, Spain): Mobility and transfer to native flora. J. Soil Sediment 2010, 10, 301–312. [Google Scholar] [CrossRef]
- Mulligan, C.N.; Yong, R.N.; Gibbs, B.F. Remediation technologies for metal-contaminated soils and groundwater: An evaluation. Eng. Geol. 2001, 60, 193–207. [Google Scholar] [CrossRef]
- Wang, S.L.; Zhao, X.Y. On the potential of biological treatment for arsenic contaminated soils and groundwater. J. Environ. Manag. 2009, 90, 2367–2376. [Google Scholar] [CrossRef]
- Gadd, G.M. Bioremedial potential of microbial mechanisms of metal mobilization and immobilization. Curr. Opin. Biotechnol. 2000, 11, 271–279. [Google Scholar] [CrossRef]
- Singh, A.; Prasad, S.M. Remediation of heavy metal contaminated ecosystem: An overview on technology advancement. Int. J. Environ. Sci. Technol. 2015, 12, 353–366. [Google Scholar] [CrossRef]
- Stolz, J.F.; Oremland, R.S. Bacterial respiration of arsenic and selenium. FEMS Microbiol. Rev. 1999, 23, 615–627. [Google Scholar] [CrossRef]
- Brierley, C.L. Bioremediation of metal-contaminated surface and groundwaters. Geomicrobiol. J. 1990, 8, 201–223. [Google Scholar] [CrossRef]
- Jaiswal, V.; Saxena, S.; Kaur, I.; Dubey, P.; Nand, S.; Naseem, M.; Singh, S.B.; Srivastava, P.K.; Barik, S.K. Application of four novel fungal strains to remove arsenic from contaminated water in batch and column modes. J. Hazard. Mater. 2018, 356, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Pumpel, T.; Schinner, F. Native fungal pellets as biosorbent for heavy metals. FEMS Microbiol. Rev. 1993, 11, 159–164. [Google Scholar] [CrossRef]
- Volesky, B.; Holan, Z.R. Biosorption of heavy metals. Biotechnol. Prog. 1995, 11, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Cernansky, S.; Urik, M.; Sevc, J.; Khun, K. Biosorption and biovolatilization of arsenic by heat-resistant fungi. Environ. Sci. Pollut. Res. 2007, 14, 31–35. [Google Scholar] [CrossRef]
- Pearce, R.B.; Callow, M.E.; Macaskie, L.E. Fungal volatilization of arsenic and antimony and the sudden infant death syndrome. FEMS Microbiol. Lett. 1998, 158, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Granchinho, S.C.R.; Franz, C.M.; Polishchuk, E.; Cullen, W.R.; Reimer, K.J. Transformation of arsenic(V) by the fungus Fusarium oxysporum melonis isolated from the alga Fucus gardneri. Appl. Organomet. Chem. 2002, 16, 721–726. [Google Scholar] [CrossRef]
- Carrasco, J.A.; Armario, P.; Pajuelo, E.; Burgos, A.; Caviedes, M.A.; Lόpez, R.; Chamber, M.A.; Palomares, A.J. Isolation and characterization of symbiotically effective Rhizobium resistant to arsenic and heavy metals after the toxic spill at the Aznalcóllar pyrite mine. Soil Biol. Biochem. 2005, 37, 1131–1140. [Google Scholar] [CrossRef]
- Vala, A.K. Tolerance and removal of arsenic by a facultative marine fungus Aspergillus candidus. Bioresour. Technol. 2010, 101, 2565–2567. [Google Scholar] [CrossRef]
- Srivastava, P.K.; Vaish, A.; Dwivedi, S.; Chakrabarty, D.; Singh, N.; Tripathi, R.D. Biological removal of arsenic pollution by soil fungi. Sci. Total Environ. 2011, 409, 2430–2442. [Google Scholar] [CrossRef]
- Rodríguez, I.A.; Martínez-Juárez, V.M.; Cárdenas-González, J.F.; Moctezuma-Zárate, M.G. Biosorption of arsenic(III) from aqueous solutions by modified fungal biomass of Paecilomyces sp. Bioinorg. Chem. Appl. 2013, 376780, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Murad, F.N.; Kabbashi, N.A.; Alam, M.Z.; Al-Khatib, M.F.R. Immobilization of fungal biomass with multi-walled carbon nanotubes as biosorbent. Malays. J. Microbiol. 2018, 14, 113–119. [Google Scholar]
- Afkhami, A.; Saber-Tehrani, M.; Bagheri, H. Modified maghemite nanoparticles as an efficient adsorbent for removing some cationic dyes from aqueous solution. Desalination 2010, 263, 240–248. [Google Scholar] [CrossRef]
- Xu, P.; Zeng, G.M.; Huang, D.L.; Feng, C.L.; Hu, S.; Zhao, M.H.; Lai, C.; Wei, Z.; Huang, C.; Xie, G.X.; et al. Use of iron oxide nanomaterials in wastewater treatment: A review. Sci. Total Environ. 2012, 424, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Gupta, M. Synthesis and surface engineering of iron oxide nanoparticles for biomedical applications. Biomaterials 2005, 26, 3995–4021. [Google Scholar] [CrossRef]
- Machala, J.; Zboril, R.; Gedanken, A. Amorphous iron(III) oxides: A review. J. Phys. Chem. B 2007, 111, 4003–4018. [Google Scholar] [CrossRef] [PubMed]
- Langmuir, I. The adsorption of gases on plane surfaces of glass, mica and platinum. J. Am. Chem. Soc. 1918, 40, 1361–1403. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Method 3051A: Microwave Assisted Acid Digestion of Sediments, Sludges, Soils, and Oils; U.S. Gov. Print Office: Washington, DC, USA, 2007.
- Gadd, G.M. Interactions of fungi with toxic metals. New Phytol. 1993, 124, 25–60. [Google Scholar] [CrossRef]
- Babich, H.; Stotzky, G. Heavy metal toxicity to microbe-mediated ecologic processes: A review and potential application to regulatory policies. Environ. Res. 1985, 36, 111–137. [Google Scholar] [CrossRef]
- Visagie, C.M.; Jacobs, K. Three new additions to the genus Talaromyces isolated from Atlantis sandveld fynbos soils. Persoonia 2012, 28, 14–24. [Google Scholar] [CrossRef]
- Hassan, N.; Rafiq, M.; Rehman, M.; Sajjad, W.; Hasan, F.; Abdullah, S. Fungi in acidic fire: A potential source of industrially important enzymes. Fungal Biol. Rev. 2019, 33, 58–71. [Google Scholar] [CrossRef]
- Romero, M.C.; Reinoso, E.H.; Urrutia, M.I.; Kiernan, A.M. Biosorption of heavy metals by Talaromyces helicus: A trained fungus for copper and biphenyl detoxification. Elect. J. Biotechnol. 2006, 9, 1–11. [Google Scholar] [CrossRef]
- Bengtsson, L.; Johansson, B.; Hackett, T.J.; McHale, L.; Mchale, A.P. Studies on the biosorption of uranium by Talaromyces emersonii CBS 814.70 biomass. Appl. Microbiol. Biotechnol. 1995, 42, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Tebo, B.M.; Obraztsova, A.Y. Sulfate-reducing bacterium grows with Cr(VI), U(VI), Mn(IV) and Fe(III) as electron acceptors. FEMS Microbiol. Lett. 1998, 162, 193–198. [Google Scholar] [CrossRef]
- Santini, J.M.; Sly, L.I.; Schnagl, R.D.; Macy, J.M. A new chemolithoautotrophic arsenite-oxidizing bacterium isolated from a gold mine: Phylogenetic, physiological, and preliminary biochemical studies. Appl. Environ. Microbiol. 2000, 66, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Niggemyer, A.; Spring, S.; Stackebrandt, E.; Rosenzweig, R.F. Isolation and characterization of a novel As(V)-reducing bacterium: Implications for arsenic mobilization and the genus Desulfitobacterium. Appl. Environ. Microbiol. 2001, 67, 5568–5580. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.; Aqil, F.; Ahmad, I. Metal tolerance and biosorption potential of filamentous fungi isolated from metal contaminated agricultural soil. Bioresour. Technol. 2007, 98, 2557–2561. [Google Scholar] [CrossRef] [PubMed]
- Canovas, D.; de Lorenzo, V. Osmotic stress limits arsenic hyper-tolerance in Aspergillus sp. P37. FEMS Microbiol. Ecol. 2007, 61, 258–263. [Google Scholar] [CrossRef]
- Sakurai, I. Biomethylation of arsenic is essentially detoxicating event. J. Health Sci. 2003, 49, 171–178. [Google Scholar] [CrossRef]
- Urík, M.; Čerňanský, S.; Ševc, J.; Šimonovičová, A.; Littera, P. Biovolatilization of arsenic by different fungal strains. Water Air Soil Pollut. 2007, 186, 337–342. [Google Scholar] [CrossRef]
- Gadd, G.M.; White, C. Microbial treatment of metal pollution—A working biotechnology? Trends Biotechnol. 1993, 11, 353–359. [Google Scholar] [CrossRef]
- Churchill, S.A.; Walters, J.V.; Churchill, P.F. Sorption of heavy metals by prepared bacterial cell surfaces. J. Environ. Eng. 1995, 121, 706–711. [Google Scholar] [CrossRef]
- Zhou, J.L. Zn biosorption by Rhizopus arrhizus and other fungi. Appl. Microbiol. Biotechnol. 1999, 51, 686–693. [Google Scholar] [CrossRef]
- Bai, R.S.; Abraham, T.E. Studies on chromium(VI) adsorption-desorption using immobilized fungal biomass. Bioresour. Technol. 2003, 87, 17–26. [Google Scholar] [PubMed]
- Yan, G.; Viraraghavan, T. Heavy-metal removal from aqueous solution by fungus Mucor rouxii. Water Res. 2003, 37, 4486–4496. [Google Scholar] [CrossRef]
- Gadd, G.M. Microbial influence on metal mobility and application for bioremediation. Geoderma 2004, 122, 109–119. [Google Scholar] [CrossRef]
- Pillichshammer, M.; Pumpel, T.; Poder, R.; Eller, K.; Klima, J.; Schinner, F. Biosorption of chromium to fungi. Biometals 1995, 8, 117–121. [Google Scholar] [CrossRef]
- Fogarty, R.V.; Tobin, J.M. Fungal melanins and their interactions with metals. Enzyme Microbiol. Technol. 1996, 19, 311–317. [Google Scholar] [CrossRef]
- Thirunavukkarasu, O.S.; Viraraghavan, T.; Subramanian, K.S. Arsenic removal from drinking water using iron oxide-coated sand. Water Air Soil Pollut. 2003, 142, 95–111. [Google Scholar] [CrossRef]
- Ghimire, K.N.; Inoue, K.; Makino, K.; Miyajima, T. Adsorptive removal of arsenic using orange juice residue. Sep. Sci. Technol. 2002, 37, 2785–2799. [Google Scholar] [CrossRef]
- Say, R.; Yilmaz, N.; Denizli, A. Biosorption of cadmium, lead, mercury, and arsenic ions by the fungus Penicillium purpurogenum. Sep. Sci. Technol. 2003, 38, 2039–2053. [Google Scholar] [CrossRef]
- Loukidou, M.X.; Matis, K.A.; Zouboulis, A.I.; Liakopoulou-Kyriakidou, M. Removal of As(V) from wastewaters by chemically modified fungal biomass. Water Res. 2003, 37, 4544–4552. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arsenic Tolerance Index in Solid and Liquid Media | Arsenic Removal (%) | |||||
|---|---|---|---|---|---|---|
| As (mg/L) | PDA | PDB | PDB | |||
| As(III) | As(V) | As(III) | As(V) | As(III) | As(V) | |
| 0 | 1.0 | 1.0 | 1.0 | 1.0 | 0 | 0 |
| 50 | 0.98 | 0.96 | 0.98 | 0.98 | 5.24 | 26.2 |
| 100 | 0.96 | 0.97 | 0.98 | 0.98 | 4.41 | 17.25 |
| 200 | 0.96 | 0.96 | 0.97 | 0.97 | 2.8 | 7.53 |
| 300 | 0.98 | 0.97 | 0.13 | 0.96 | 0.41 | 1.24 |
| 500 | 0.96 | 0.97 | 0.008 | 0.67 | 0 | 0.78 |
| 800 | 0.96 | 0.97 | 0.008 | 0.5 | 0 | 0.52 |
| 1000 | 0.96 | 0.98 | 0.004 | 0.35 | 0 | 0.32 |
| Treatment a | Arsenic Removal (mean ± SD b, %) | |
|---|---|---|
| As(III) | As(V) | |
| Untreated biomass | 4 ± 0.32 | 13 ± 0.97 |
| Fe(III) hydroxide-coated biomass | 69 ± 2.79 | 79 ± 3.65 |
| nFe(III) oxide-coated biomass | 91 ± 5.25 | 96 ± 4.18 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, I.-H.; Murugesan, K.; Ryu, J.; Kim, J.H. Arsenic (As) Removal Using Talaromyces sp. KM-31 Isolated from As-Contaminated Mine Soil. Minerals 2019, 9, 568. https://doi.org/10.3390/min9100568
Nam I-H, Murugesan K, Ryu J, Kim JH. Arsenic (As) Removal Using Talaromyces sp. KM-31 Isolated from As-Contaminated Mine Soil. Minerals. 2019; 9(10):568. https://doi.org/10.3390/min9100568
Chicago/Turabian StyleNam, In-Hyun, Kumarasamy Murugesan, Jungho Ryu, and Jae Hwan Kim. 2019. "Arsenic (As) Removal Using Talaromyces sp. KM-31 Isolated from As-Contaminated Mine Soil" Minerals 9, no. 10: 568. https://doi.org/10.3390/min9100568
APA StyleNam, I. -H., Murugesan, K., Ryu, J., & Kim, J. H. (2019). Arsenic (As) Removal Using Talaromyces sp. KM-31 Isolated from As-Contaminated Mine Soil. Minerals, 9(10), 568. https://doi.org/10.3390/min9100568