The Role of the Substrate on the Mineralization Potential of Microbial Mats in A Modern Freshwater River (Paris Basin, France)

 , , , , ,
, , , , , 
Abstract
:1. Introduction
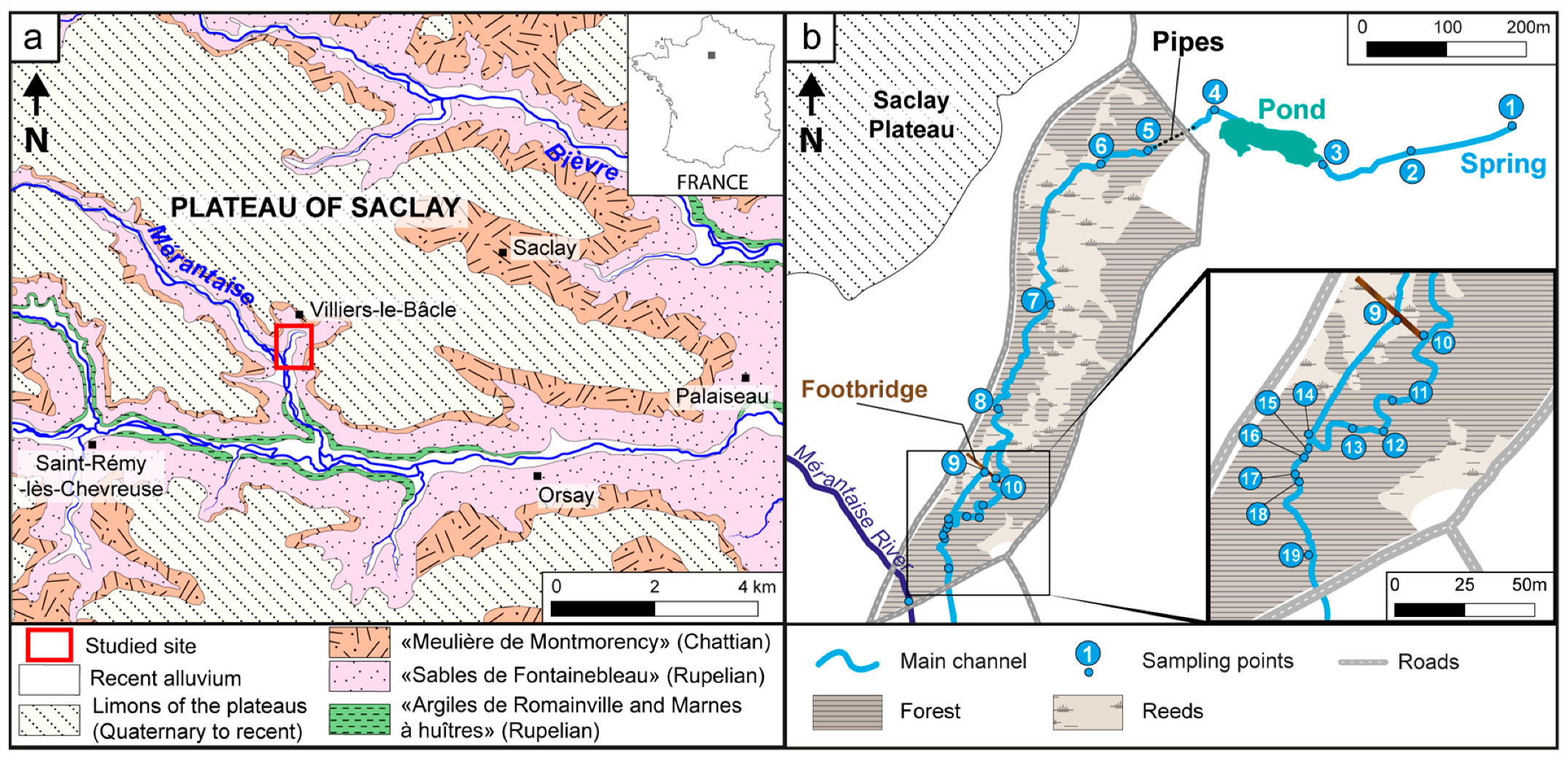
2. Geographical, Geological and Climatic Context
3. Materials and Methods
4. Results
4.1. Physicochemical Parameters of the Stream
4.1.1. Stream Morphology and Associated Vegetation
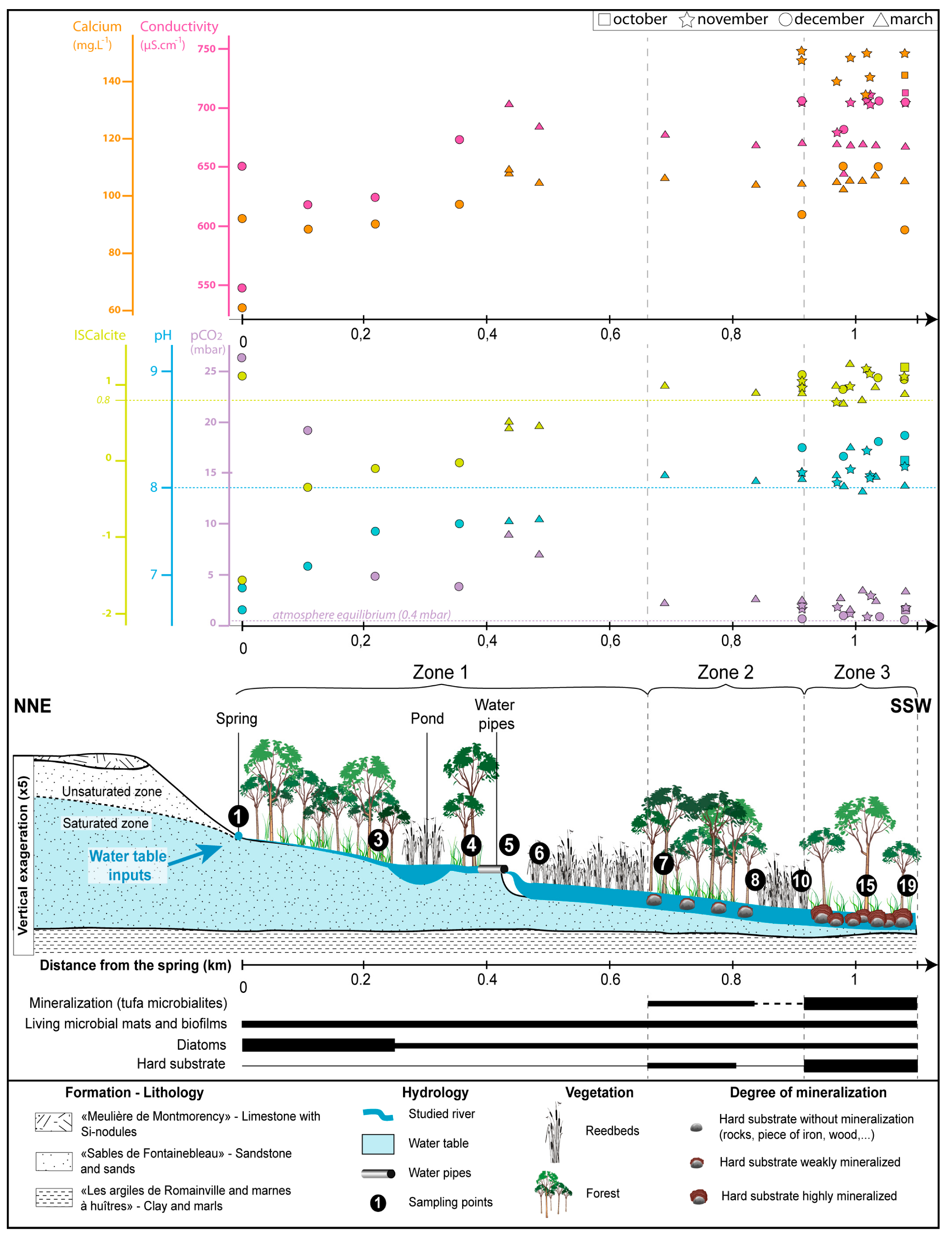
4.1.2. Hydrology and Water Chemistry
4.2. Organosedimentary Structures
4.2.1. Microbial Mats and Biofilms
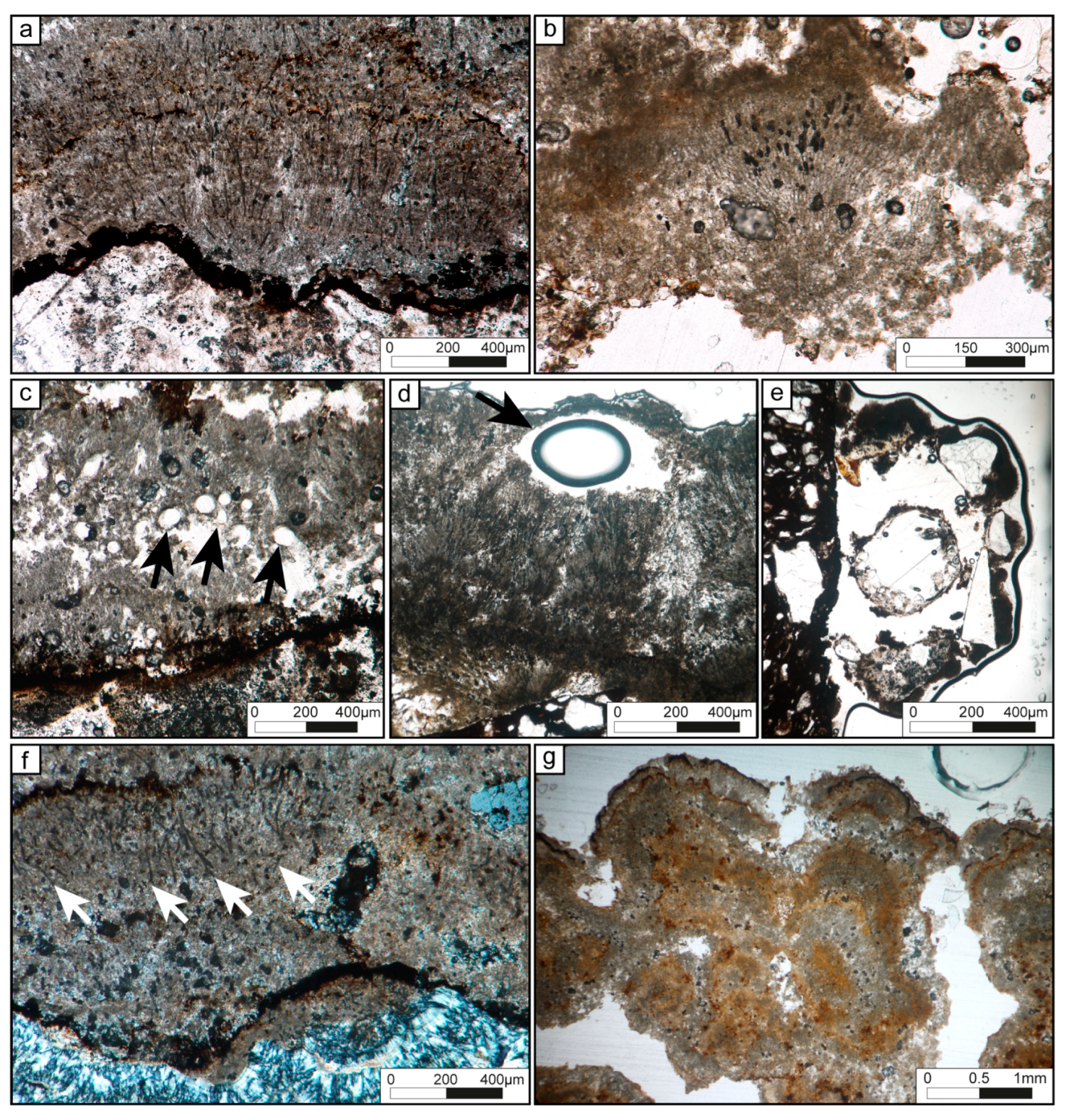
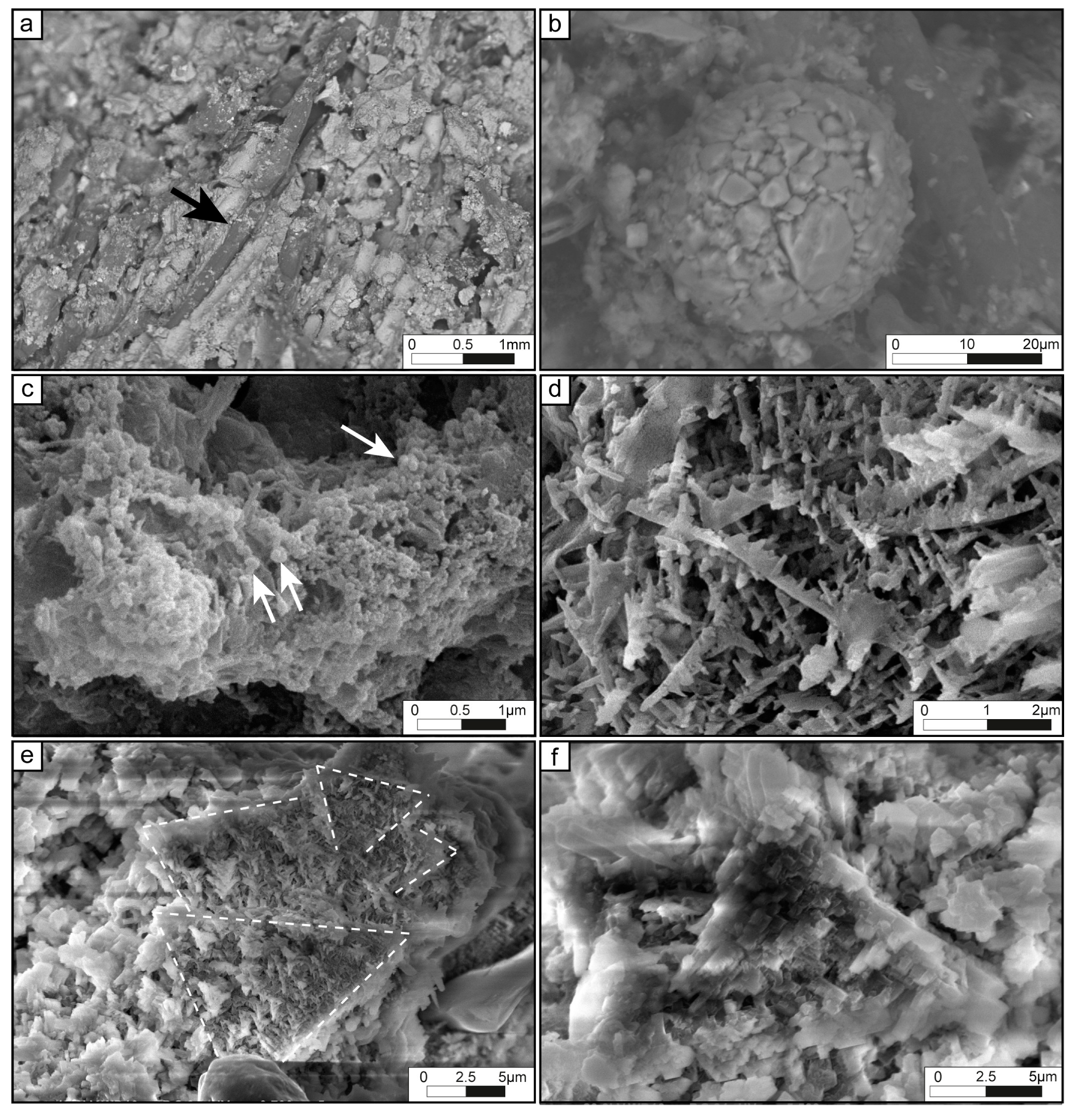
4.2.2. Tufa Microbialites
4.3. Zonation of the Mineralization Intensity
4.3.1. Zone 1: No Mineralization
4.3.2. Zone 2: Low Mineralization
4.3.3. Zone 3: High Mineralization
5. Discussion
5.1. External Controlling Processes on Mineralization
5.2. Link Between Living-Microbial and Mineralized Organosedimentary Structures
5.3. Role of Substrate on the Spatial Distribution of Tufa
5.3.1. Physical Role of the Substrate
5.3.2. Chemical Role of the Substrate
5.4. External Control on the tufa Microbialite Fabric
6. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pedley, H.M. Classification and environmental models of cool freshwater tufas. Sediment. Geol. 1990, 68, 143–154. [Google Scholar] [CrossRef]
- Ford, T.D.; Pedley, H.M. A review of tufa and travertine deposits of the world. Earth-Sci. Rev. 1996, 41, 117–175. [Google Scholar] [CrossRef]
- Della Porta, G. Carbonate build-ups in lacustrine, hydrothermal and fluvial settings: Comparing depositional geometry, fabric types and geochemical signature. Geol. Soc. Lond. Spec. Publ. 2015, 418, 17–68. [Google Scholar] [CrossRef]
- Manzo, E.; Perri, E.; Tucker, M.E. Carbonate deposition in a fluvial tufa system: processes and products (Corvino Valley-southern Italy): Carbonate fluvial tufa deposition, processes and products. Sedimentology 2012, 59, 553–577. [Google Scholar] [CrossRef]
- Shiraishi, F.; Reimer, A.; Bissett, A.; de Beer, D.; Arp, G. Microbial effects on biofilm calcification, ambient water chemistry and stable isotope records in a highly supersaturated setting (Westerhöfer Bach, Germany). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 262, 91–106. [Google Scholar] [CrossRef]
- Visscher, P.T.; Stolz, J.F. Microbial mats as bioreactors: Populations, processes, and products. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 87–100. [Google Scholar] [CrossRef]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth-Sci. Rev. 2009, 96, 141–162. [Google Scholar] [CrossRef]
- Benzerara, K.; Skouri-Panet, F.; Li, J.; Ferard, C.; Gugger, M.; Laurent, T.; Couradeau, E.; Ragon, M.; Cosmidis, J.; Menguy, N.; et al. Intracellular Ca-carbonate biomineralization is widespread in cyanobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, 10933–10938. [Google Scholar] [CrossRef] [Green Version]
- Bougeault, C.; Vennin, E.; Durlet, C.; Muller, E.; Mercuzot, M.; Chavez, M.; Gérard, E.; Ader, M.; Virgone, A.; Gaucher, E.C. Biotic-abiotic influences on modern Ca-Si-rich hydrothermal spring mounds of the Pastos Grandes volcanic caldera (Bolivia). Minerals, Under revisions.
- Gradziński, M. Factors controlling growth of modern tufa: results of a field experiment. Geol. Soc. Lond. Spec. Publ. 2010, 336, 143–191. [Google Scholar] [CrossRef]
- Stal, L.J.; van Gemerden, H.; Krumbein, W.E. Structure and development of a benthic marine microbial mat. FEMS Microbiol. Ecol. 1985, 1, 111–125. [Google Scholar] [CrossRef]
- Ginsburg, R.N.; Planavsky, N.J. Diversity of Bahamian Microbialite Substrates. In Links Between Geological Processes, Microbial Activities&Evolution of Life: Microbes and Geology; Dilek, Y., Furnes, H., Muehlenbachs, K., Eds.; Modern Approaches in Solid Earth Sciences; Springer: Dordrecht, The Netherlands, 2008; pp. 177–195. ISBN 978-1-4020-8306-8. [Google Scholar] [CrossRef]
- Dromart, G.; Gaillard, C.; Jansa, L.F. Deep-Marine Microbial Structures in the Upper Jurassic of Western Tethys. In Phanerozoic Stromatolites II; Bertrand-Sarfati, J., Monty, C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 295–318. ISBN 978-94-011-1124-9. [Google Scholar] [CrossRef]
- Benson, L. Carbonate deposition, Pyramid Lake subbasin, Nevada: 1. Sequence of formation and elevational distribution of carbonate deposits (Tufas). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 109, 55–87. [Google Scholar] [CrossRef]
- Chafetz, H.; Rush, P.F.; Utech, N.M. Microenvironmental controls on mineralogy and habit of CaCO3 precipitates: An example from an active travertine system. Sedimentology 1991, 38, 107–126. [Google Scholar] [CrossRef]
- Emeis, K.-C.; Richnow, H.-H.; Kempe, S. Travertine formation in Plitvice National Park, Yugoslavia: chemical versus biological control. Sedimentology 1987, 34, 595–609. [Google Scholar] [CrossRef]
- Janssen, A.; Swennen, R.; Podoor, N.; Keppens, E. Biological and diagenetic influence in Recent and fossil tufa deposits from Belgium. Sediment. Geol. 1999, 126, 75–95. [Google Scholar] [CrossRef]
- Merz-Preiss, M.; Riding, R. Cyanobacterial tufa calcification in two freshwater streams: Ambient environment, chemical thresholds and biological processes. Sediment. Geol. 1999, 126, 103–124. [Google Scholar] [CrossRef]
- Folk, R. Interaction between bacteria, nannobacteria, and mineral precipitation in hot springs of central Italy. Géographie Phys. Quat. 1994, 48, 233–246. [Google Scholar] [CrossRef]
- Parsiegla, K.I.; Katz, J.L. Calcite growth inhibition by copper (II): I. Effect of supersaturation. J. Cryst. Growth 1999, 200, 213–226. [Google Scholar] [CrossRef]
- Parsiegla, K.I.; Katz, J.L. Calcite growth inhibition by copper (II): II. Effect of solution composition. J. Cryst. Growth 2000, 213, 368–380. [Google Scholar] [CrossRef]
- Baumgartner, L.K.; Spear, J.R.; Buckley, D.H.; Pace, N.R.; Reid, R.P.; Dupraz, C.; Visscher, P.T. Microbial diversity in modern marine stromatolites, Highborne Cay, Bahamas. Environ. Microbiol. 2009, 11, 2710–2719. [Google Scholar] [CrossRef]
- Visscher, P.T.; Dupraz, C.; Braissant, O.; Gallagher, K.L.; Glunk, C.; Casillas, L.; Reed, R.E.S. Biogeochemistry of Carbon Cycling in Hypersaline Mats: Linking the Present to the Past through Biosignatures. In Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems; Seckbach, J., Oren, A., Eds.; Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 2010; pp. 443–468. ISBN 978-90-481-3799-2. [Google Scholar] [CrossRef]
- Perri, E.; Manzo, E.; Tucker, M.E. Multi-scale study of the role of the biofilm in the formation of minerals and fabrics in calcareous tufa. Sediment. Geol. 2012, 263–264, 16–29. [Google Scholar] [CrossRef]
- Sforna, M.C.; Daye, M.; Philippot, P.; Somogyi, A.; van Zuilen, M.A.; Medjoubi, K.; Gérard, E.; Jamme, F.; Dupraz, C.; Braissant, O.; et al. Patterns of metal distribution in hypersaline microbialites during early diagenesis: Implications for the fossil record. Geobiology 2017, 15, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Perri, E.; Tucker, M.E.; Słowakiewicz, M.; Whitaker, F.; Bowen, L.; Perrotta, I.D. Carbonate and silicate biomineralization in a hypersaline microbial mat (Mesaieed sabkha, Qatar): Roles of bacteria, extracellular polymeric substances and viruses. Sedimentology 2018, 65, 1213–1245. [Google Scholar] [CrossRef]
- Pedley, H.M.; Rogerson, M. In vitro investigations of the impact of different temperature and flow velocity conditions on tufa microfabric. Geol. Soc. Lond. Spec. Publ. 2010, 336, 193–210. [Google Scholar] [CrossRef]
- Pedley, M.; Rogerson, M.; Middleton, R. Freshwater calcite precipitates from in vitro mesocosm flume experiments: A case for biomediation of tufas. Sedimentology 2009, 56, 511–527. [Google Scholar] [CrossRef]
- Dupraz, C.; Visscher, P.T.; Baumgartner, L.K.; Reid, R.P. Microbe–mineral interactions: early carbonate precipitation in a hypersaline lake (Eleuthera Island, Bahamas). Sedimentology 2004, 51, 745–765. [Google Scholar] [CrossRef]
- Pomerol, C.; Afchain, C.; Labesse, C.; Rampon, G.; Renard, M.; Fédoroff, N.; Bournérias, M.; Jovet, P.; Cavelier, C.; Perreau, M.; et al. Notice de la carte géologique de France (1/50000), feuille Rambouillet (218); BRGM: Orléans, France, 1975. [Google Scholar]
- Lefebvre, K. Diagnostic et quantification des flux nappe-rivière: Modélisations hydrodynamique et géochimique du bassin versant de l’Yvette amont (France). Ph.D. Thesis, Paris-Saclay University, Orsay, France, December 2015. [Google Scholar]
- Corcho Alvarado, J.A.; Purtschert, R.; Barbecot, F.; Chabault, C.; Rueedi, J.; Schneider, V.; Aeschbach-Hertig, W.; Kipfer, R.; Loosli, H.H. Constraining the age distribution of highly mixed groundwater using 39Ar: A multiple environmental tracer (3H/3He, 85Kr, 39Ar, and 14C) study in the semiconfined Fontainebleau Sands Aquifer (France): CONSTRAINING THE AGE OF GROUNDWATER. Water Resour. Res. 2007, 43. [Google Scholar] [CrossRef]
- Renard, F.; Tognelli, A. A new quasi-3D unsaturated–saturated hydrogeologic model of the Plateau de Saclay (France). J. Hydrol. 2016, 535, 495–508. [Google Scholar] [CrossRef]
- Schneider, V. Apports de l’hydrodynamique et de la géochimie à la caractérisation des nappes de l’Oligocène et de l’Eocène et à la reconnaissance de leurs relations actuelles et passées: origine de la dégradation de la nappe de l’Oligocène (sud-ouest du bassin parisien). Ph.D. Thesis, Paris-Saclay University, Orsay, France, November 2005. [Google Scholar]
- Gran, G. Equivalence volumes in potentiometric titrations. Anal. Chim. Acta 1988, 206, 111–123. [Google Scholar] [CrossRef]
- Parkhurst, D.L.; Appelo, C.A.J. User’s guide to PHREEQC—A computer program for speciation, reaction-path, 1D-transport, and inverse geochemical calculations. US Geol. Surv. Water-Resour. Investig. Rep. 1999, 99, 4259. [Google Scholar] [CrossRef]
- Charlton, S.R.; Parkhurst, D.L. PHREEQCI–A Graphical User Interface to the Geochemical Model PHREEQC; U.S. Geological Survey: Denver, CO, USA, 2002. [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Piper, A.M. A graphic procedure in the geochemical interpretation of water-analyses. Eos Trans. Am. Geophys. Union 1944, 25, 914–928. [Google Scholar] [CrossRef]
- Bouton, A.; Vennin, E.; Pace, A.; Bourillot, R.; Dupraz, C.; Thomazo, C.; Brayard, A.; Désaubliaux, G.; Visscher, P.T. External controls on the distribution, fabrics and mineralization of modern microbial mats in a coastal hypersaline lagoon, Cayo Coco (Cuba). Sedimentology 2016, 63, 972–1016. [Google Scholar] [CrossRef]
- Ley, R.E.; Harris, J.K.; Wilcox, J.; Spear, J.R.; Miller, S.R.; Bebout, B.M.; Maresca, J.A.; Bryant, D.A.; Sogin, M.L.; Pace, N.R. Unexpected Diversity and Complexity of the Guerrero Negro Hypersaline Microbial Mat. Appl Env. Microbiol 2006, 72, 3685–3695. [Google Scholar] [CrossRef] [Green Version]
- Decho, A.W. Microbial biofilms in intertidal systems: An overview. Cont. Shelf Res. 2000, 20, 1257–1273. [Google Scholar] [CrossRef]
- Krumbein, W.E.; Brehm, U.; Gerdes, G.; Gorbushina, A.A.; Levit, G.; Palinska, K.A. Biofilm, Biodictyon, Biomat Microbialites, Oolites, Stromatolites Geophysiology, Global Mechanism, Parahistology. In Fossil and Recent Biofilms: A Natural History of Life on Earth; Krumbein, W.E., Paterson, D.M., Zavarzin, G.A., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 1–27. ISBN 978-94-017-0193-8. [Google Scholar] [CrossRef]
- Neu, T.R. Biofilms and microbial mats. In Biostabilization of Sediments; Bibliotheks und Informationssystem der Carl von Ossietzky Universitat: Oldenburg, Germany, 1994; pp. 9–15. [Google Scholar]
- Noffke, N. Geobiology Microbial Mats in Sandy Deposits from the Archean Era to Today; Springer: Berlin, Germany; London, UK, 2010; ISBN 978-3-642-12772-4. [Google Scholar] [CrossRef]
- Jørgensen, B.B. Distribution of colorless sulfur bacteria (Beggiatoa spp.) in a coastal marine sediment. Mar. Biol. 1977, 41, 19–28. [Google Scholar] [CrossRef]
- Kelly, D.P.; Postgate, J.R.; Kelly, D.P. Biochemistry of the chemolithotrophic oxidation of inorganic sulphur. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1982, 298, 499–528. [Google Scholar] [CrossRef] [PubMed]
- Marvasi, M.; Visscher, P.T.; Casillas Martinez, L. Exopolymeric substances (EPS) from Bacillus subtilis: polymers and genes encoding their synthesis. FEMS Microbiol. Lett. 2010, 313, 1–9. [Google Scholar] [CrossRef]
- Diatoms of North America. Available online: https://diatoms.org/ (accessed on 13 May 2019).
- Freytet, P.; Plet, A. Modern freshwater microbial carbonates: The Phormidium stromatolites (tufa-travertine) of southeastern Burgundy (Paris Basin, France). Facies 1996, 34, 219. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Al-Horani, F.A.; Farmer, J.D.; Ludwig, R.; Wade, B.D. Balance between microbial calcification and metazoan bioerosion in modern stromatolitic oncolites. Geobiology 2004, 2, 49–57. [Google Scholar] [CrossRef]
- Berrendero, E.; Arenas, C.; Mateo, P.; Jones, B. Cyanobacterial diversity and related sedimentary facies as a function of water flow conditions: Example from the Monasterio de Piedra Natural Park (Spain). Sediment. Geol. 2016, 337, 12–28. [Google Scholar] [CrossRef]
- Leggitt, V.L.; Cushman, R.A. Complex caddisfly-dominated bioherms from the Eocene Green River Formation. Sediment. Geol. 2001, 145, 377–396. [Google Scholar] [CrossRef]
- Andrews, J.E.; Brasier, A.T. Seasonal records of climatic change in annually laminated tufas: Short review and future prospects. J. Quat. Sci. 2005, 20, 411–421. [Google Scholar] [CrossRef]
- Kleinteich, J.; Golubic, S.; Pessi, I.S.; Velázquez, D.; Storme, J.-Y.; Darchambeau, F.; Borges, A.V.; Compère, P.; Radtke, G.; Lee, S.-J.; et al. Cyanobacterial Contribution to Travertine Deposition in the Hoyoux River System, Belgium. Microb. Ecol. 2017, 74, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Kano, A.; Matsuoka, J.; Kojo, T.; Fujii, H. Origin of annual laminations in tufa deposits, southwest Japan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 191, 243–262. [Google Scholar] [CrossRef]
- Arp, G.; Bissett, A.; Brinkmann, N.; Cousin, S.; De Beer, D.; Friedl, T.; Mohr, K.I.; Neu, T.R.; Reimer, A.; Shiraishi, F.; et al. Tufa-forming biofilms of German karstwater streams: microorganisms, exopolymers, hydrochemistry and calcification. Geol. Soc. Lond. Spec. Publ. 2010, 336, 83–118. [Google Scholar] [CrossRef]
- Arenas, C.; Jones, B. Temporal and environmental significance of microbial lamination: Insights from Recent fluvial stromatolites in the River Piedra, Spain. Sedimentology 2017, 64, 1597–1629. [Google Scholar] [CrossRef]
- Kawai, T.; Kano, A.; Matsuoka, J.; Ihara, T. Seasonal variation in water chemistry and depositional processes in a tufa-bearing stream in SW-Japan, based on 5 years of monthly observations. Chem. Geol. 2006, 232, 33–53. [Google Scholar] [CrossRef]
- Kempe, S. Long-term records of CO2 pressure fluctuations in fresh waters. In Transport of Carbon and Minerals in Major World Rivers; Mitt. Geol.-Paläont. Inst. Univ. Hamburg, SCOPE/UNEP Sonderband: Hamburg, Germany, 1982; Volume 52, pp. 91–332. [Google Scholar]
- Arp, G.; Wedemeyer, N.; Reitner, J. Fluvial tufa formation in a hard-water creek (Deinschwanger Bach, Franconian Alb, Germany). Facies 2001, 44, 1–22. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.D.; Wang, S.; Xiao, T.; Huang, R. Factors controlling tufa deposition in natural waters at waterfall sites. Sediment. Geol. 2004, 166, 353–366. [Google Scholar] [CrossRef]
- Shiraishi, F.; Okumura, T.; Takahashi, Y.; Kano, A. Influence of microbial photosynthesis on tufa stromatolite formation and ambient water chemistry, SW Japan. Geochim. Cosmochim. Acta 2010, 74, 5289–5304. [Google Scholar] [CrossRef]
- Arenas, C.; Vázquez-Urbez, M.; Auqué, L.; Sancho, C.; Osácar, C.; Pardo, G. Intrinsic and extrinsic controls of spatial and temporal variations in modern fluvial tufa sedimentation: A thirteen-year record from a semi-arid environment. Sedimentology 2014, 61, 90–132. [Google Scholar] [CrossRef]
- Drysdale, R.; Gillieson, D. Micro-erosion meter measurements of travertine deposition rates: a case study from Louie Creek, Northwest Queensland, Australia. Earth Surf. Process. Landf. J. Br. Geomorphol. Group 1997, 22, 1037–1051. [Google Scholar] [CrossRef]
- Vázquez-Urbez, M.; Arenas, C.; Sancho, C.; Osácar, C.; Auqué, L.; Pardo, G. Factors controlling present-day tufa dynamics in the Monasterio de Piedra Natural Park (Iberian Range, Spain): depositional environmental settings, sedimentation rates and hydrochemistry. Int. J. Earth Sci. 2010, 99, 1027–1049. [Google Scholar] [CrossRef]
- Auqué, L.; Arenas, C.; Osácar, C.; Pardo, G.; Sancho, C.; Vázquez-Urbez, M. Current tufa sedimentation in a changing-slope valley: The River Añamaza (Iberian Range, NE Spain). Sediment. Geol. 2014, 303, 26–48. [Google Scholar] [CrossRef]
- Pedley, M. Ambient Temperature Freshwater Microbial Tufas. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 179–186. ISBN 978-3-662-04036-2. [Google Scholar] [CrossRef]
- Pentecost, A. Travertine; Springer Science & Business Media: New York, NY, USA, 2005; ISBN 978-1-4020-3523-4. [Google Scholar]
- Pentecost, A. Calcium Carbonate Deposition by Blue–Green Algae. Ph.D. Thesis, University College of North Wales, Bangor, UK, 1975. [Google Scholar]
- Zhang, D.D.; Zhang, Y.; Zhu, A.; Cheng, X. Physical Mechanisms of River Waterfall Tufa (Travertine) Formation. J. Sediment. Res. 2001, 71, 205–216. [Google Scholar] [CrossRef]
- Visscher, P.T.; Gemerden, H. van Production and Consumption of Dimethylsulfoniopropionate in Marine Microbial Mats. Appl. Env. Microbiol. 1991, 57, 3237–3242. [Google Scholar]
- Sand-Jensen, K. Photosynthetic Carbon Sources of Stream Macrophytes. J. Exp. Bot. 1983, 34, 198–210. [Google Scholar] [CrossRef]
- Winsborough, B.M. Diatoms and Benthic Microbial Carbonates. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 76–83. ISBN 978-3-662-04036-2. [Google Scholar] [CrossRef]
- Turner, E.C.; Jones, B. Microscopic calcite dendrites in cold-water tufa: Implications for nucleation of micrite and cement. Sedimentology 2005, 52, 1043–1066. [Google Scholar] [CrossRef]
- Pedley, M. Freshwater (phytoherm) reefs: The role of biofilms and their bearing on marine reef cementation. Sediment. Geol. 1992, 79, 255–274. [Google Scholar] [CrossRef]
- Rittmann, B. Detachment from biofilms. In Structure and Function of Biofilms; John Wiley & Sons: New York, NY, USA, 1989; pp. 49–58. [Google Scholar]
- Stolzenbach, K.D. Particle transport and attachment. In Structure and Function of Biofilms; Characklis, W.G., Wilderer, P.A., Eds.; J. Wiley & Sons: Chichester, UK, 1989; pp. 33–47. [Google Scholar] [CrossRef]
- Primc-Habdija, B.; Habdija, I.; Plenković-Moraj, A. elka Tufa deposition and periphyton overgrowth as factors affecting the ciliate community on travertine barriers in different current velocity conditions. Hydrobiologia 2001, 457, 87–96. [Google Scholar] [CrossRef]
- Cox, E.J. Identification of Freshwater Diatoms from Live Material; Chapman & Hall: London, UK, 1996; ISBN 978-0-412-49380-5. [Google Scholar]
- Lavoie, I.; Hamilton, P.B. Guide D’identification des Diatomées des Rivières de l’Est du Canada; PUQ: Québec, QC, Canada, 2008; ISBN 978-2-7605-1984-8. [Google Scholar]
- Brasier, A.T.; Andrews, J.E.; Kendall, A.C. Diagenesis or dire genesis? The origin of columnar spar in tufa stromatolites of central Greece and the role of chironomid larvae. Sedimentology 2011, 58, 1283–1302. [Google Scholar] [CrossRef]
- Butcher, R.W. Studies in the Ecology of Rivers: VI. The Algal Growth in Certain Highly Calcareous Streams. J. Ecol. 1946, 33, 268–283. [Google Scholar] [CrossRef]
- Golubić, S.; Marčenko, E. Über Konvergenzerscheinungen bei Standortsformen der Blaualgen unter extremen Lebensbedingungen. Schweiz. Z. Für Hydrol. 1965, 27, 207–217. [Google Scholar] [CrossRef]
- Golubić, S. The relationship between blue–green algae and carbonate deposits. In The Biology of Blue–green Algae; Carr, N.G., Whitton, B.A., Eds.; Basil Blackwell: Oxford, UK, 1973; pp. 434–472. [Google Scholar]
- Reid, R.P.; Visscher, P.T.; Decho, A.W.; Stolz, J.F.; Bebout, B.M.; Dupraz, C.; Macintyre, I.G.; Paerl, H.W.; Pinckney, J.L.; Prufert-Bebout, L.; et al. The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 2000, 406, 989. [Google Scholar] [CrossRef] [PubMed]
- Trichet, J.; Défarge, C.; Tribble, J.; Tribble, G.; Sansone, F. Christmas Island lagoonal lakes, models for the deposition of carbonate–evaporite–organic laminated sediments. Sediment. Geol. 2001, 140, 177–189. [Google Scholar] [CrossRef]
- Dupraz, C.; Visscher, P.T. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payandi-Rolland, D.; Roche, A.; Vennin, E.; Visscher, P.T.; Amiotte-Suchet, P.; Thomas, C.; Bundeleva, I. Carbonate precipitation in freshwater cyanobacterial biofilms forming microbial tufa. Minerals, Submitted.
- Chekroun, K.B.; Rodríguez-Navarro, C.; González-Muñoz, M.T.; Arias, J.M.; Cultrone, G.; Rodríguez-Gallego, M. Precipitation and Growth Morphology of Calcium Carbonate Induced by Myxococcus Xanthus: Implications for Recognition of Bacterial Carbonates. J. Sediment. Res. 2004, 74, 868–876. [Google Scholar] [CrossRef]
- Folk, R.L. Nannobacteria and the precipitation of carbonate in unusual environments. Sediment. Geol. 1999, 126, 47–55. [Google Scholar] [CrossRef]
- Pedley, M. The morphology and function of thrombolitic calcite precipitating biofilms: A universal model derived from freshwater mesocosm experiments. Sedimentology 2014, 61, 22–40. [Google Scholar] [CrossRef]
- Rogerson, M.; Pedley, H.M.; Middleton, R. Microbial influence on macroenvironment chemical conditions in alkaline (tufa) streams: Perspectives from in vitro experiments. Geol. Soc. Lond. Spec. Publ. 2010, 336, 65–81. [Google Scholar] [CrossRef]
- Visscher, P.T.; Reid, R.P.; Bebout, B.M.; Hoeft, S.E.; Macintyre, I.G.; Thompson, J.A. Formation of lithified micritic laminae in modern marine stromatolites (Bahamas); the role of sulfur cycling. Am. Mineral. 1998, 83, 1482–1493. [Google Scholar] [CrossRef]
- Winsborough, B.M.; Seeler, J.-S.; Golubic, S.; Folk, R.L.; Maguire, B. Recent Fresh-Water Lacustrine Stromatolites, Stromatolitic Mats and Oncoids from Northeastern Mexico. In Phanerozoic Stromatolites II; Bertrand-Sarfati, J., Monty, C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 71–100. ISBN 978-94-011-1124-9. [Google Scholar] [CrossRef]
- Casanova, J. Stromatolites from the East African Rift: A Synopsis. In Phanerozoic Stromatolites II; Bertrand-Sarfati, J., Monty, C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 193–226. ISBN 978-94-011-1124-9. [Google Scholar] [CrossRef]
- Moore, L.S.; Burne, R.V. The Modern Thrombolites of Lake Clifton, Western Australia. In Phanerozoic Stromatolites II; Bertrand-Sarfati, J., Monty, C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 3–29. ISBN 978-94-011-1124-9. [Google Scholar] [CrossRef]
- Humphreys, W.F.; Awranmik, S.M.; Jebb, M.H.P. Freshwater biogenic tufa dams in Madang Province, Papua New Guinea. J. R. Soc. West. Aust. 1995, 13. [Google Scholar] [CrossRef]
- Drysdale, R.N. The sedimentological significance of hydropsychid caddis-fly larvae (order; Trichoptera) in a travertine-depositing stream; Louie Creek, Northwest Queensland, Australia. J. Sediment. Res. 1999, 69, 145–150. [Google Scholar] [CrossRef]
- Bertrand-Sarfati, J.; Freytet, P.; Plaziat, J.C. Microstructures in Tertiary Nonmarine Stromatolites (France). Comparison with Proterozoic. In Phanerozoic Stromatolites II; Bertrand-Sarfati, J., Monty, C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 155–191. ISBN 978-94-011-1124-9. [Google Scholar] [CrossRef]
- Roche, A.; Vennin, E.; Bouton, A.; Olivier, N.; Wattinne, A.; Bundeleva, I.; Deconinck, J.-F.; Virgone, A.; Gaucher, E.C.; Visscher, P.T. Oligo-Miocene lacustrine microbial and metazoan buildups from the Limagne Basin (French Massif Central). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 504, 34–59. [Google Scholar] [CrossRef]
- Bouton, A.; Vennin, E.; Boulle, J.; Pace, A.; Bourillot, R.; Thomazo, C.; Brayard, A.; Désaubliaux, G.; Goslar, T.; Yokoyama, Y.; et al. Linking the distribution of microbial deposits from the Great Salt Lake (Utah, USA) to tectonic and climatic processes. Biogeosciences 2016, 13, 5511–5526. [Google Scholar] [CrossRef] [Green Version]
- Vennin, E.; Bouton, A.; Bourillot, R.; Pace, A.; Roche, A.; Brayard, A.; Thomazo, C.; Virgone, A.; Gaucher, E.C.; Desaubliaux, G.; et al. The lacustrine microbial carbonate factory of the successive Lake Bonneville and Great Salt Lake, Utah, USA. Sedimentology 2019, 66, 165–204. [Google Scholar] [CrossRef]
- Vanden Berg, M.D. Domes, Rings, Ridges, and Polygons: Characteristics of Microbialites from Utah’s Great Salt Lake. Sediment. Rec. 2019, 17, 4–10. [Google Scholar] [CrossRef]
- Paterson, D.M.; Aspden, R.J.; Visscher, P.T.; Consalvey, M.; Andres, M.S.; Decho, A.W.; Stolz, J.; Reid, R.P. Light-Dependant Biostabilisation of Sediments by Stromatolite Assemblages. PLoS ONE 2008, 3, e3176. [Google Scholar] [CrossRef]
- Jahnert, R.J.; Collins, L.B. Significance of subtidal microbial deposits in Shark Bay, Australia. Mar. Geol. 2011, 286, 106–111. [Google Scholar] [CrossRef]
- Bailey, J.V.; Orphan, V.J.; Joye, S.B.; Corsetti, F.A. Chemotrophic Microbial Mats and Their Potential for Preservation in the Rock Record. Astrobiology 2009, 9, 843–859. [Google Scholar] [CrossRef] [Green Version]
- Tuchman, M.L.; Stevenson, R.J. Comparison of clay tile, sterilized rock, and natural substrate diatom communities in a small stream in Southeastern Michigan, USA. Hydrobiologia 1980, 75, 73–79. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Resh, V.H. Distribution of benthic algae and macroinvertebrates along a thermal stream gradient. Hydrobiologia 1985, 128, 13–21. [Google Scholar] [CrossRef]
- Harmsworth, G.C.; Sleigh, M.A. Colonization of non-living surfaces in streams by peritrich ciliates. Eur. J. Protistol. 1993, 29, 294–301. [Google Scholar] [CrossRef]
- Hellal, J.; Michel, C.; Barsotti, V.; Laperche, V.; Garrido, F.; Joulian, C. Representative sampling of natural biofilms: influence of substratum type on the bacterial and fungal communities structure. SpringerPlus 2016, 5, 822. [Google Scholar] [CrossRef] [PubMed]
- García-Meza, J.V.; Barrangue, C.; Admiraal, W. Biofilm formation by algae as a mechanism for surviving on mine tailings. Environ. Toxicol. Chem. 2005, 24, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Vasconcelos, M.T. Cyanobacteria Metal Interactions: Requirements, Toxicity, and Ecological Implications. Crit. Rev. Microbiol. 2006, 32, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.; Rodriguez-Eaton, S.; Ekanemesang, U.M.; Phillips, P. Characterization of metal-binding bioflocculants produced by the cyanobacterial component of mixed microbial mats. Appl. Env. Microbiol. 1994, 60, 2311–2315. [Google Scholar] [Green Version]
- Konhauser, K.O.; Ferris, F.G. Diversity of iron and silica precipitation by microbial mats in hydrothermal waters, Iceland: Implications for Precambrian iron formations. Geology 1996, 24, 323–326. [Google Scholar] [CrossRef]
- Kawai, T.; Kano, A.; Hori, M. Geochemical and hydrological controls on biannual lamination of tufa deposits. Sediment. Geol. 2009, 213, 41–50. [Google Scholar] [CrossRef]
- Kano, A.; Hagiwara, R.; Kawai, T.; Hori, M.; Matsuoka, J. Climatic Conditions and Hydrological Change Recorded in a High-Resolution Stable-Isotope Profile of a Recent Laminated Tufa on a Subtropical Island, Southern Japan. J. Sediment. Res. 2007, 77, 59–67. [Google Scholar] [CrossRef]
- Gerdes, G.; Krumbein, W.E.; Reineck, H.E. Biolaminations—Ecological versus depositional dynamics. In Cycles and Events in Stratigraphy; Springer: Berlin, Germany; New York, NY, USA, 1991; pp. 592–607. [Google Scholar]
- Chafetz, H.S.; Folk, R.L. Travertines; depositional morphology and the bacterially constructed constituents. J. Sediment. Res. 1984, 54, 289–316. [Google Scholar] [CrossRef]
- Dupraz, C.; Fowler, A.; Tobias, C.; Visscher, P.T. Stromatolitic knobs in Storr’s Lake (San Salvador, Bahamas): A model system for formation and alteration of laminae. Geobiology 2013, 11, 527–548. [Google Scholar] [CrossRef] [PubMed]
- Monty, C.L.V. Chapter 5.1 The Origin and Development of Cryptalgal Fabrics. In Developments in Sedimentology; Walter, M.R., Ed.; Stromatolites; Elsevier: Amsterdam, The Netherlands; Oxford, UK; New York, NY, USA, 1976; Volume 20, pp. 193–249. [Google Scholar] [CrossRef]
- Chafetz, H.S.; Utech, N.M.; Fitzmaurice, S.P. Differences in the delta 18O and delta 13C signatures of seasonal laminae comprising travertine stromatolites. J. Sediment. Res. 1991, 61, 1015–1028. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Phoenix, V.R.; Bottrell, S.H.; Adams, D.G.; Head, I.M. Microbial–silica interactions in Icelandic hot spring sinter: possible analogues for some Precambrian siliceous stromatolites. Sedimentology 2001, 48, 415–433. [Google Scholar] [CrossRef]
- Golubić, S.; Violante, C.; Plenković-Moraj, A.; Grgasović, T. Travertines and calcareous tufa deposits: An insight into diagenesis. Geol. Croat. 2008, 61, 363–378. [Google Scholar] [CrossRef]
- Reid, R.P.; James, N.P.; Macintyre, I.G.; Dupraz, C.P.; Burne, R.V. Shark Bay stromatolites: Microfabrics and reinterpretation of origins. Facies 2003, 49, 299. [Google Scholar] [CrossRef]
- Kano, A.; Fujii, H. Origin of the gross morphology and internal texture of tufas of Shirokawa Town, Ehime Prefecture, southwest Japan. J. Geol. Soc. Jpn. 2000, 106, 397–412. [Google Scholar] [CrossRef]
- Melón, P.; Alonso-Zarza, A.M. The Villaviciosa tufa: A scale model for an active cool water tufa system, Guadalajara (Spain). Facies 2018, 64. [Google Scholar] [CrossRef]
- Pentecost, A.; Coletta, P. The role of photosynthesis and CO2 evasion in travertine formation: a quantitative investigation at an important travertine-depositing hot spring, Le Zitelle, Lazio, Italy. J. Geol. Soc. 2007, 164, 843–853. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
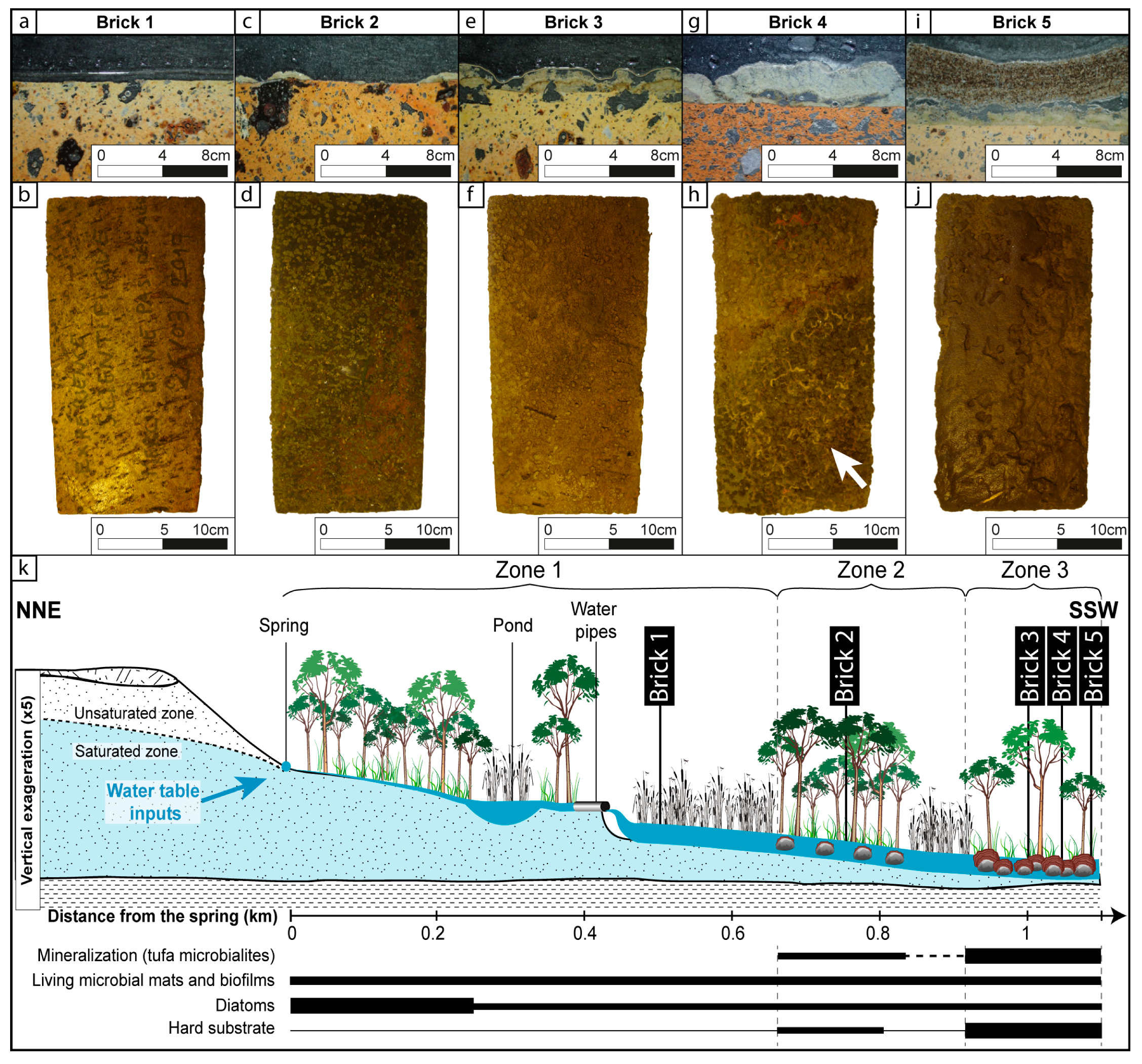{kind=link}
{kind=link}
{kind=link}
{kind=link}
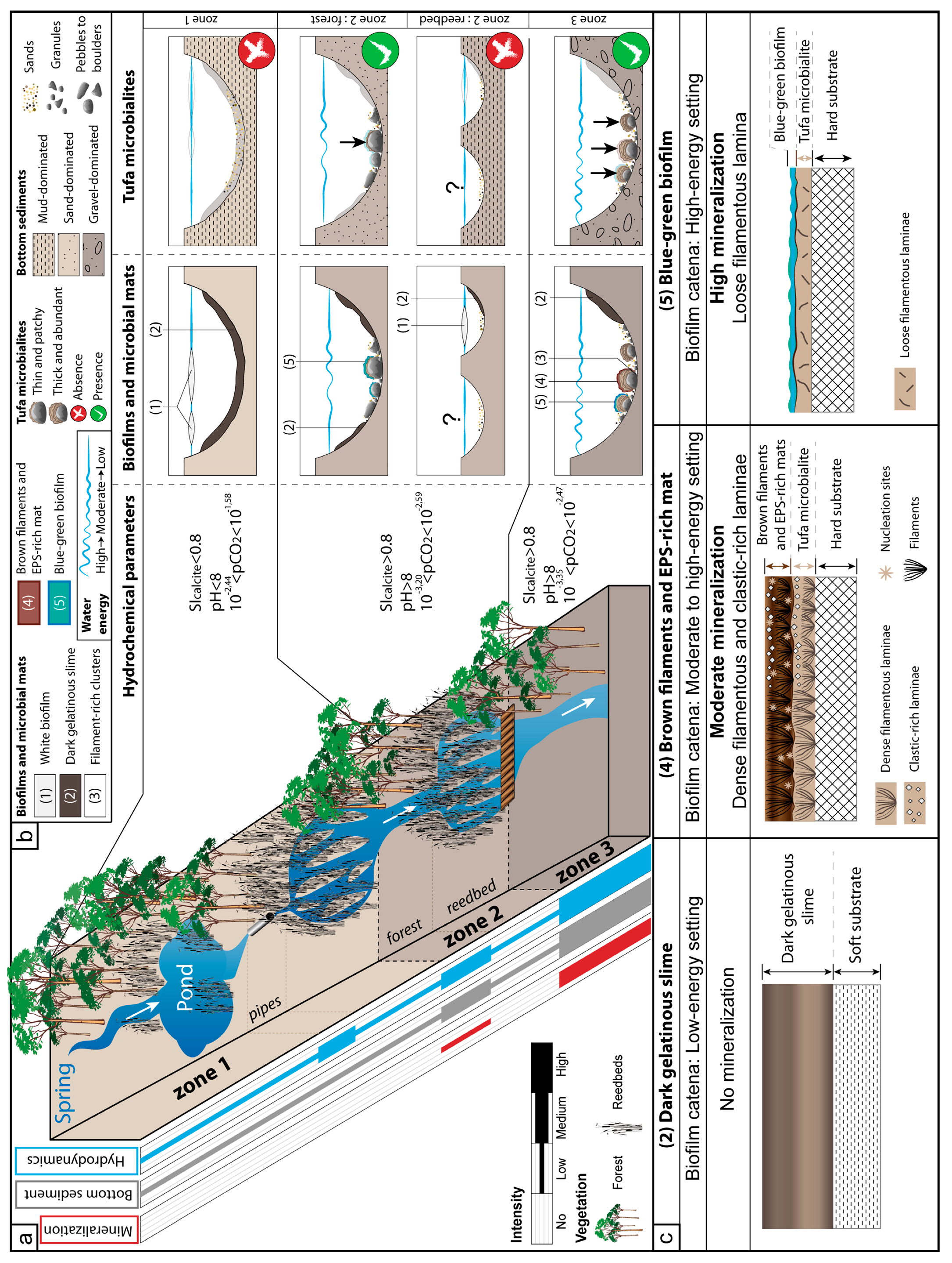{kind=link}
| Zone 1 | Zone 2 | Zone 3 | ||
|---|---|---|---|---|
| No Mineralization | Low Mineralization | High Mineralization | ||
| Sampling points | 1 to 7 | 7 to 8; 9 to 10 | 8 to 9 | 10 to 19 |
| Distance from spring | 0 to 680 m | 680 to 840 m | 840 to 970 m | 970 to 1100 m |
| Physical Parameters | ||||
| Channel | Single to multiple | Single | Multiple | Single |
| Depth | Shallow (<10 cm) | Shallow to deep (6 to 30 cm depth) | Shallow (<10 cm) | Shallow to deep (5 cm to 15 cm depth) |
| Water velocity | Slow (<0.023 m.s−1) | Slow to moderate (0.04 to 0.208 m.s−1) | Slow to moderate (0.071 to 0.137 m.s−1) | Slow to fast (0.025 to 0.585 m.s−1) |
| Topographic break | Pipes | - | - | Dams |
| Granulometry of bottom sediment | Mud to sand, few pebbles | Mud to pebbles, some blocs | Mud to sand | Mud to blocs |
| Nature of substrate | Siliceous grains (quartz), wood | Siliceous grains (chert and quartz), woods, roots or anthropogenic (pottery) | Siliceous grains (chert and quartz), wood | Siliceous grains (chert and quartz), wood, roots, anthropogenic (brick, iron, glass) |
| Biological Components | ||||
| Vegetation | Forest/Reed bed | Forest | Reed bed | Forest clearing |
| Biofilms, microbial mats | White biofilms; dark gelatinous slime | Dark gelatinous slime; blue-green biofilm; algae | Dark gelatinous slime; few white biofilms; algae | Dark gelatinous slime; brown filaments and EPS-rich mats; blue-green biofilm; algae |
| Tufa Microbialite | ||||
| Mineralized crusts | None | Thin and patchy | None | Thick and abundant |
| Chemical Parameters * | ||||
| SI calcite | −1.56 to 0.52; −0.5 | 0.89 to 1.13; 0.99 | 0.75 to 1.27; 1.01 | |
| Conductivity (μS.cm−1) | 547 to 703; 641 | 668 to 705; 685 | 644 to 712; 686 | |
| pH | 6.6 to 7.64; 7.34 | 8.08 to 8.46; 8.18 | 7.96 to 8.53; 8.25 | |
| pCO2 (mbar) | 26.30 to 3.63; 11.60 | 2.57 to 0.63; 1.95 | 3.39 to 0.45; 1.77 | |
| Zone | pH | Conductivity (μS.cm−1) | Alkalinity (meq.L−1) | TOC (ppm) | pCO2 (mbar) | pCO2 (x atm. conc.) | Anions (mg.L−1) | Cations (mg.L−1) | IScalc | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cl− | NO3− | SO42− | PO43− | HCO3_− | Na+ | K+ | Mg2+ | Ca2+ | ||||||||
| 1 | 7.34 | 641 | 4.1 | 6.34 | 11.60 | 29 | 27.29 | 23.24 | 56.78 | - | 250.1 | 15.88 | 2.82 | 9.76 | 91.63 | -0.5 |
| 2 | 8.18 | 685 | 5.38 | 6.17 | 1.95 | 5 | 27.01 | 38.22 | 50.20 | - | 328.15 | 15.39 | 3.28 | 11.33 | 115.75 | 0.99 |
| 3 | 8.25 | 686 | 5.04 | 6.14 | 1.77 | 4 | 27.71 | 47.64 | 45.59 | - | 307.24 | 16.25 | 2.72 | 11.85 | 121.72 | 1.01 |
| Microfabrics | Morphology of Laminae | Matrix | Main Components of Laminae | Other Components | Preservation/ Porosity | Substrate | Biological Affinity | Figures | |
|---|---|---|---|---|---|---|---|---|---|
| Dense filamentous | Flat or slightly wavy laminae (100 to 1000 µm thick), composed of hemispheroid and fan-shaped structures, frequently sharply truncated by dissolution surface. | Microsparite or light micrite. | Molds of filaments organized in fan-shaped and erected perpendicular to the basement. Composed of two-size types of filaments: 12 to 13 µm in diameter filaments slightly arched and 500 µm long. Segmentation may be observed every 17 to 23 µm; 2-4 µm in diameter straight filaments with and 25 to 100 µm long. Molds are empty or filled by dark matrix (oxides?). In MEB view, remnants sheath preserved in filaments molds. | Scarce detrital grains (quartz, K-feldspars, plagioclases; 100 to 200 µm) trapped in the residual porosity between the filaments; scarce circles shapes (Chlorellopsis-like?) with a diameter ranging from 50 to 100 µm within or at the top of laminations; few oxides crystals (from 10 to 30 µm-width); larval housings, Trichoptera pupal cases and diatoms. | Large vuggy to interparticular porosity; dissolution cavity is filled by detrital grains. | Micritic laminae, loose filaments laminae. Boulders of different composition. | Thick filaments: Phormidium (?) Thin filaments: Leptolyngbya sp. | Figures 4a–d, and 6d | |
| Loose filamentous | Flat to wavy laminae from 100 to 900 µm-thick, sometimes showing fan-shaped structures or concentrically organization; interlayered with micritic laminae with planar laminations of different colors; laminae are frequently sharply truncated by dissolution surfaces. | Dense micrite, locally clotted. | Molds of scattered straight filaments branching or not with diameters from 4 µm to 10 µm and 10 to 100 µm long; isolated filaments embedded in micritic matrix with local clotted organization; walls highlighted by dark lines or filled by a dark matrix; filaments are perpendicular to substrate or disorganized. In MEB view, remnant sheath preserved in filaments molds. | Few oxides crystals (10 to 40 µm); diatoms. | Weak vuggy porosity. | Dense filaments laminae, micritic laminae with or without quartz. Boulders of different composition. | Phormidium sp. | Figures 4f,g, and 6d | |
| Clastic-rich | 300 to 800 µm-thick discontinuous or continuous laminae , with dark rims at the top and dissolution surfaces. | Micritic or light microsparite. | Peloids, angular and unsorted quartz 30 to 100 µm, scarce K-feldspars or plagioclases. | Rare disorganized thin molds of filaments (4 to 6 µm). | Strong to weak vuggy porosity. | Iron-rich laminae with quartz, chert. | - | Figure 6a | |
| Micritic | Wavy laminae from 30 to 300 µm-thick, top are often limited by dark rims and dissolution surfaces. | Micritic or light microsparite. | Homogeneous dense micrite, lined by black rims enriched in iron (?). | Few oxide crystals (20 to 120 µm). | - | Mainly in contact with the boulders; dense filament laminae; iron-rich laminae with or without quartz. | - | Figure 6b,c | |
| Iron-rich | without quartz | 30 to 100 µm-thick laminae, with thin µm-thick wavy laminations. | Dark brown micrite or microsparite. | Dense dark matrix composed of goethite and siderite (?). | Small oxide crystals (20 to 40 µm). | No porosity | Micritic laminae with quartz, iron, chert. | - | Figure 6b,d |
| with quartz | 100 to 500 µm-thick laminae; delimited by irregular and black rims and dissolution surfaces. | Dark clotted micrite, grey micrite or microsparite. | Dense dark matrix composed of goethite and siderite (?); peloids, angular and unsorted quartz (20 µm to 300 µm) trapped in the matrix, scarce K-feldspars, plagioclase or siliceous-rich fragments. | Oxide crystals (10 to 30 µm), rare wood fragments. | No porosity | Micritic laminae with quartz, chert, wood. | - | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roche, A.; Vennin, E.; Bundeleva, I.; Bouton, A.; Payandi-Rolland, D.; Amiotte-Suchet, P.; Gaucher, E.C.; Courvoisier, H.; Visscher, P.T. The Role of the Substrate on the Mineralization Potential of Microbial Mats in A Modern Freshwater River (Paris Basin, France). Minerals 2019, 9, 359. https://doi.org/10.3390/min9060359
Roche A, Vennin E, Bundeleva I, Bouton A, Payandi-Rolland D, Amiotte-Suchet P, Gaucher EC, Courvoisier H, Visscher PT. The Role of the Substrate on the Mineralization Potential of Microbial Mats in A Modern Freshwater River (Paris Basin, France). Minerals. 2019; 9(6):359. https://doi.org/10.3390/min9060359
Chicago/Turabian StyleRoche, Adeline, Emmanuelle Vennin, Irina Bundeleva, Anthony Bouton, Dahédrey Payandi-Rolland, Philippe Amiotte-Suchet, Eric C. Gaucher, Hélène Courvoisier, and Pieter T. Visscher. 2019. "The Role of the Substrate on the Mineralization Potential of Microbial Mats in A Modern Freshwater River (Paris Basin, France)" Minerals 9, no. 6: 359. https://doi.org/10.3390/min9060359
APA StyleRoche, A., Vennin, E., Bundeleva, I., Bouton, A., Payandi-Rolland, D., Amiotte-Suchet, P., Gaucher, E. C., Courvoisier, H., & Visscher, P. T. (2019). The Role of the Substrate on the Mineralization Potential of Microbial Mats in A Modern Freshwater River (Paris Basin, France). Minerals, 9(6), 359. https://doi.org/10.3390/min9060359