
On Large and Small Data Blow-Up Solutions in the Trojan Y Chromosome Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
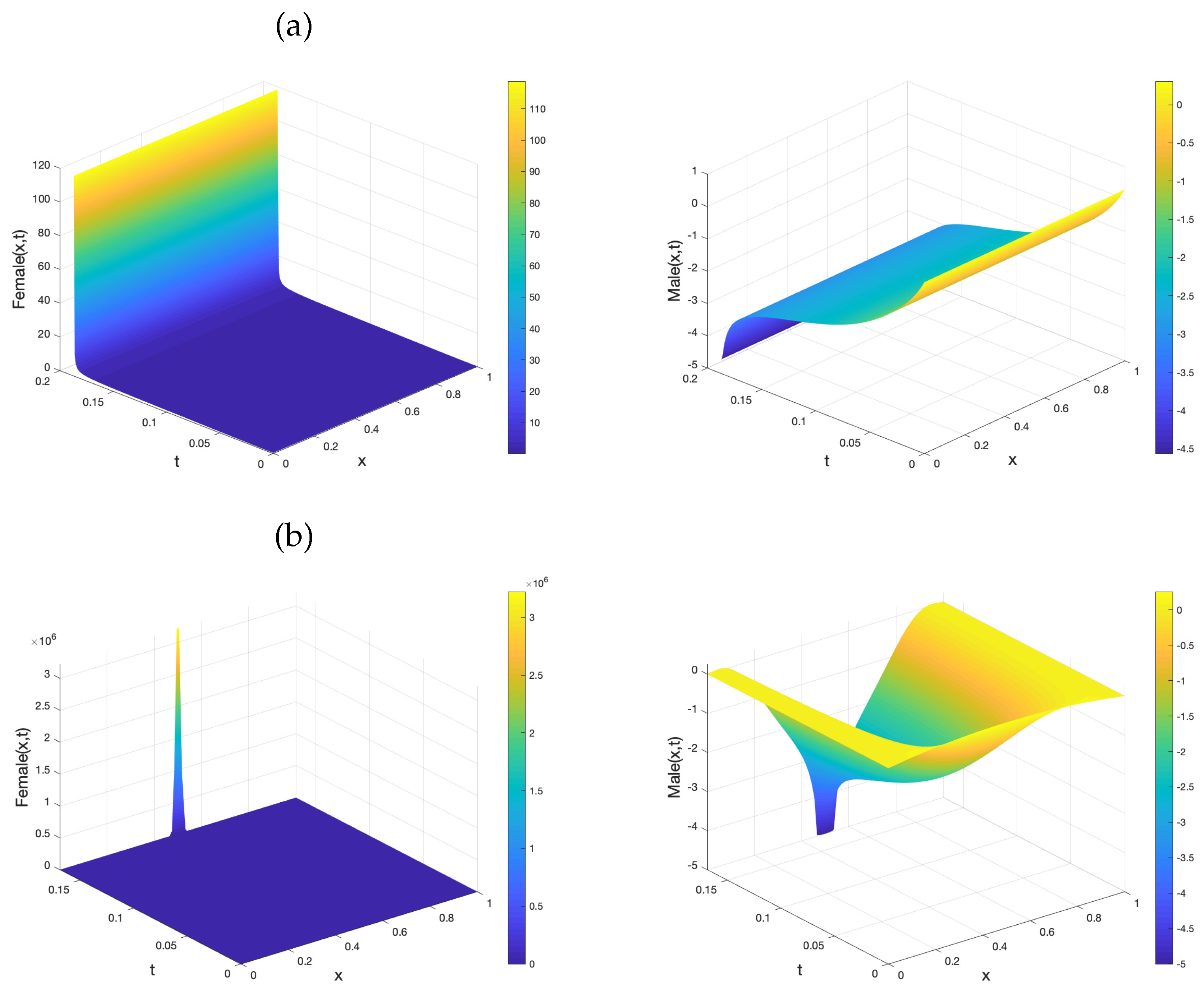
- The three species reaction diffusion TYC model, (1)–(3), results in negative solutions in the male population m. These conditions include the choice of large initial conditions () and/or large introduction rate of supermales s. This result is given via Lemma 2;
- The negativity of m may lead to finite time blow-up in of f and m. This happens for large initial data as seen via Theorem 1, but is also possible for small initial data, which is seen via Theorem 2, in the PDE model. Similar results are derived via Corollary A1, Corollary A2 for the ODE model;
- We numerically derive regions of invariance in the phase space, for the three species TYC PDE model, via Figure 2. Additionally, chemotaxis type mechanisms can damp blow-up solutions;
- We discuss the practical relevance of these results to biological control and possible restrictions.

2. Finite Time Blow up
2.1. Preliminaries
2.2. Negative Solutions
2.3. Finite Time Blow-Up in the PDE Model
3. Numerical Experiments for TYC PDE Model
3.1. Scaling of TYC PDE Model
3.2. Spatial Damping
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Finite Time Blow-Up in the ODE Model
Appendix A.1. Three Species TYC Model
Appendix A.2. Finite Time Blow-Up of Solutions
Appendix B. Scaling of Chemotaxis TYC PDE Models
References
- Arim, M.; Abades, S.; Neill, P.; Lima, M.; Marquet, P. Spread Dynamics of invasive species. Proc. Natl. Acad. Sci. USA 2006, 103, 374–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bampfylde, C.J.; Lewis, M.A. Biological control through intraguild predation: Case studies in pest control, invasive species and range expansion. Bull. Math. Biol. 2007, 69, 1031–1066. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.S.; Lewis, M.; Horvath, L. Invasion by extremes: Population spread with variation in dispersal and reproduction. Am. Nat. 2001, 157, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.H.; Simberloff, D.; Kuris, A.M.; Carey, J.R. Eradication revisited: Dealing with exotic species. Trends Ecol. Evol. 2000, 15, 316–320. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; Barro, P.J.D.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Bacher, S. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Shigesada, N.; Kawasaki, K. Biological Invasions: Theory and Practice; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Schofield, P.; Loftus, W. Nonnative fishes in Florida freshwaters: A literature review and synthesis. Rev. Fish Biol. Fisher. 2015, 25, 117–145. [Google Scholar] [CrossRef]
- Driesche, R.V.; Bellows, T. Biological Control; Springer Science & Business Media: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Parshad, R.D.; Wickramsooriya, S.; Bailey, S. A remark on “Biological control through provision of additional food to predators: A theoretical study” [Theoretical Population Biology 72 (2007) 111–120]. Theor. Popul. Biol. 2020, 132, 60–68. [Google Scholar] [CrossRef]
- Beauregard, M.A.; Parshad, R.D.; Boon, S.; Conaway, H.; Griffin, T.; Lyu, J. Optimal Control and Analysis of a Modified Trojan Y-Chromosome Strategy. Ecol. Model. 2020, 416, 108854. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, P.A.; Besson, J. Wild Trout Evaluations: MYY Brook Trout Field Evaluations 2019. 2020. Available online: https://www.researchgate.net/profile/Curtis-Roth-2/publication/351549966_Wild_Trout_Evaluations_MYY_Brook_Trout_Field_Evaluations_2019/links/609d1812299bf1259ed13ed5/Wild-Trout-Evaluations-MYY-Brook-Trout-Field-Evaluations-2019.pdf (accessed on 8 December 2021).
- Noble, C.; Min, J.; Olejarz, J.; Buchthal, J.; Chavez, A.; Smidler, A.L.; DeBenedictis, E.A.; Church, G.M.; Nowak, M.A.; Esvelt, K.M. Daisy-chain gene drives for the alteration of local populations. Proc. Natl. Acad. Sci. USA 2019, 116, 8275–8282. [Google Scholar] [CrossRef] [Green Version]
- Schill, D.J.; Meyer, K.A.; Hansen, M.J. Simulated effects of YY-male stocking and manual suppression for eradicating nonnative brook trout populations. N. Am. J. Fish. Manag. 2017, 37, 1054–1066. [Google Scholar]
- Dunham, R.A.; Su, B. Genetically Engineered Fish: Potential Impacts on Aquaculture, Biodiversity, and the Environment. In Topics in Biodiversity and Conservation; Chaurasia, A., Hawksworth, D.L., Pessoa de Miranda, M., Eds.; Springer: Cham, Switzerland, 2020; Volume 19. [Google Scholar] [CrossRef]
- Gutierrez, J.B.; Teem, J. A model describing the effect of sex-reversed YY fish in an established wild population: The use of a Trojan Y-Chromosome to cause extinction of an introduced exotic species. J. Theor. Biol. 2006, 241, 333–341. [Google Scholar] [CrossRef]
- McCormick, J.L.; Schill, D.J.; Meyer, K.A. Simulated use of YY Male stocking and suppression for eradicating Common Carp populations. N. Am. J. Fish. Manag. 2021, 41, 366–382. [Google Scholar] [CrossRef]
- Poirier, J.; Davis, B.; Harris, J. Feasibility Assessment of Stocking YY Males to Eradicate Nonnative Brook Trout from Tyee Springs. In Columbia River Fish and Wildlife Conservation Office 2019 Annual Report; 2020. Available online: https://www.fws.gov/sites/default/files/documents/Report-Feasibility-Assessment-of-Stocking-YY-Males-to-Eradicate-Nonnative-Brook-Trout-%20from-Tyee-Springs-2020-Columbia-River-FWCO.pdf (accessed on 8 December 2021).
- Teem, J.L.; Gutierrez, J.B.; Parshad, R.D. A comparison of the Trojan Y Chromosome model and daughterless carp eradication strategies. Biol. Invasions 2014, 16, 1217–1230. [Google Scholar] [CrossRef]
- Wedekind, C. Demographic and genetic consequences of disturbed sex determination. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotton, S.; Wedekind, C. Introduction of Trojan sex chromosomes to boost population growth. J. Theor. Biol. 2007, 249, 153–161. [Google Scholar] [CrossRef]
- Cotton, S.; Wedekind, C. Population consequences of environmental sex reversal. Conserv. Biol. 2009, 23, 196–206. [Google Scholar] [CrossRef]
- Wang, X.; Parshad, R.D.; Walton, J. The Stochastic Trojan Y Chromosome model for eradication of an invasive species. J. Biol. Dynam. 2016, 10, 179–199. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Liu, B.; Ning, D. Existence of global attractor for the Trojan Y Chromosome model. Electron. J. Qual. Theory Differ. Equ. 2012, 2012, 1–16. [Google Scholar] [CrossRef]
- Bhattacharyya, J.; Roelke, D.L.; Walton, J.R.; Banerjee, S. Using yy supermales to destabilize invasive fish populations. Theor. Popul. Biol. 2020, 134, 1–14. [Google Scholar] [CrossRef]
- Lyu, J.; Schofield, P.J.; Reaver, K.M.; Beauregard, M.A.; Parshad, R.D. A comparison of the Trojan Y Chromosome strategy to harvesting models for eradication of non-native species. Nat. Resour. Model. 2020, 33, e12252. [Google Scholar] [CrossRef] [Green Version]
- Parshad, R.D. On the long time behavior of a PDE model for invasive species control. Int. J. Mathemat. Anal. 2011, 5, 1991–2015. [Google Scholar]
- Wang, X.; Walton, J.R.; Parshad, R.D.; Storey, K.; Boggess, M. Analysis of Trojan Y-Chromosome eradication strategy. J. Math. Biol. 2014, 68, 1731–1756. [Google Scholar] [CrossRef] [PubMed]
- Schill, D.J.; Heindel, J.A.; Campbell, M.R.; Meyer, K.A.; Mamer, E.R. Production of a YY Male Brook Trout Broodstock for Potential Eradication of Undesired Brook Trout Populations. N. Am. J. Aquacult. 2016, 78, 72–83. [Google Scholar] [CrossRef]
- Kennedy, P.A.; Meyer, K.A.; Schill, D.J.; Campbell, M.R.; Vue, N.V. Survival and reproduction success of hatchery YY male brook trout stocked in Idaho streams. Trans. Am. Fish. Soc. 2018, 147, 419–430. [Google Scholar] [CrossRef]
- Murray, J.D. Mathematical Biology II: Spatial Models and Biomedical Applications; Springer: New York, NY, USA, 2001; Volume 3. [Google Scholar]
- Cantrell, R.S.; Lam, K.Y. Competitive exclusion in phytoplankton communities in a eutrophic water column. Discret. Contin. Dyn. Syst.-Ser. B 2021, 26, 1–13. [Google Scholar] [CrossRef]
- Gutierrez, J.B.; Hurdal, M.K.; Parshad, R.D.; Teem, J.L. Analysis of the Trojan Y Chromosome Model for Eradication of Invasive Species in a Dendritic Riverine System. J. Math. Biol. 2012, 64, 319–340. [Google Scholar] [CrossRef]
- Jiang, H.; Lam, K.Y.; Lou, Y. Three-patch Models for the Evolution of Dispersal in Advective Environments: Varying Drift and Network Topology. Bull. Math. Biol. 2021, 83, 1–46. [Google Scholar] [CrossRef]
- Parshad, R.D.; Kouachi, S.; Gutierrez, J.B. Global existence and asymptotic behavior of a model for biological control of invasive species via supermale introduction. Commun. Math. Sci. 2013, 11, 951–972. [Google Scholar]
- Parshad, R.D.; Gutierrez, J.B. On the Global Attractor of the Trojan Y-Chromosome Model. Commun. Pure Appl. Anal. 2011, 10, 339–359. [Google Scholar] [CrossRef]
- Ladyzhenskaia, O.A.; Solonnikov, V.A.; Ural’tseva, N.N. Linear and Quasi-Linear Equations of Parabolic Type; American Mathematical Society: Providence, RI, USA, 1988; Volume 23. [Google Scholar]
- Morgan, J. Global Existence for Semilinear Parabolic Systems. SIAM J. Math. Anal. 1989, 20, 1128–11441. [Google Scholar] [CrossRef]
- Quittner, P.; Souplet, P. Superlinear Parabolic Problems. Blow-Up, Global Existence and Steady States; Birkhuser Advanced Texts; Birkhuser: Basel, Switzerland, 2007. [Google Scholar]
- Pierre, M. Global existence in reaction-diffusion systems with control of mass: A survey. Milan J. Math. 2010, 78, 417–455. [Google Scholar] [CrossRef]
- Horstmann, D.; Winkler, M. Boundedness vs. blow-up in a chemotaxis system. J. Differ. Equ. 2005, 215, 52–107. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.M.; Dennis, B.; Liebhold, A.M.; Drake, J.M. The evidence for Allee effects. Popul. Ecol. 2009, 51, 341–354. [Google Scholar] [CrossRef]
- Parshad, R.D.; Antwi-Fordjour, K.; Takyi, E.M. Some novel results in two species competition. SIAM J. Appl. Math. 2021, 81, 1847–1869. [Google Scholar] [CrossRef]
- Tianran, Z.; Wang, W. Mathematical models of two-sex population dynamics. Kôkyûroku 2005, 1432, 96–104. [Google Scholar]
- Fitzgibbon, W.E.; Morgan, J.J.; Tang, B.Q.; Yin, H.M. Reaction-Diffusion-Advection Systems with Discontinuous Diffusion and Mass Control. SIAM J. Math. Anal. 2021, 53, 6771–6803. [Google Scholar] [CrossRef]
- Morgan, J.; Tang, B.Q. Boundedness for reaction–diffusion systems with lyapunov functions and intermediate sum conditions. Nonlinearity 2020, 33, 3105. [Google Scholar] [CrossRef]
- Day, C.C.; Landguth, E.L.; Simmons, R.K.; Baker, W.P.; Whiteley, A.R.; Lukacs, P.M.; Bearlin, A. Simulating effects of fitness and dispersal on the use of Trojan sex chromosomes for the management of invasive species. J. Appl. Ecol. 2020, 57, 1413–1425. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takyi, E.M.; Beauregard, M.A.; Griffin, T.; Bobo, L.; Parshad, R.D. On Large and Small Data Blow-Up Solutions in the Trojan Y Chromosome Model. Axioms 2022, 11, 120. https://doi.org/10.3390/axioms11030120
Takyi EM, Beauregard MA, Griffin T, Bobo L, Parshad RD. On Large and Small Data Blow-Up Solutions in the Trojan Y Chromosome Model. Axioms. 2022; 11(3):120. https://doi.org/10.3390/axioms11030120
Chicago/Turabian StyleTakyi, Eric M., Matthew A. Beauregard, Thomas Griffin, Landrey Bobo, and Rana D. Parshad. 2022. "On Large and Small Data Blow-Up Solutions in the Trojan Y Chromosome Model" Axioms 11, no. 3: 120. https://doi.org/10.3390/axioms11030120
APA StyleTakyi, E. M., Beauregard, M. A., Griffin, T., Bobo, L., & Parshad, R. D. (2022). On Large and Small Data Blow-Up Solutions in the Trojan Y Chromosome Model. Axioms, 11(3), 120. https://doi.org/10.3390/axioms11030120