
Dynamic Behaviors of an Obligate Commensal Symbiosis Model with Crowley–Martin Functional Responses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Existence and Local Stability of the Equilibria
3. Global Stability of the Equilibria
4. Nonautonomous Case
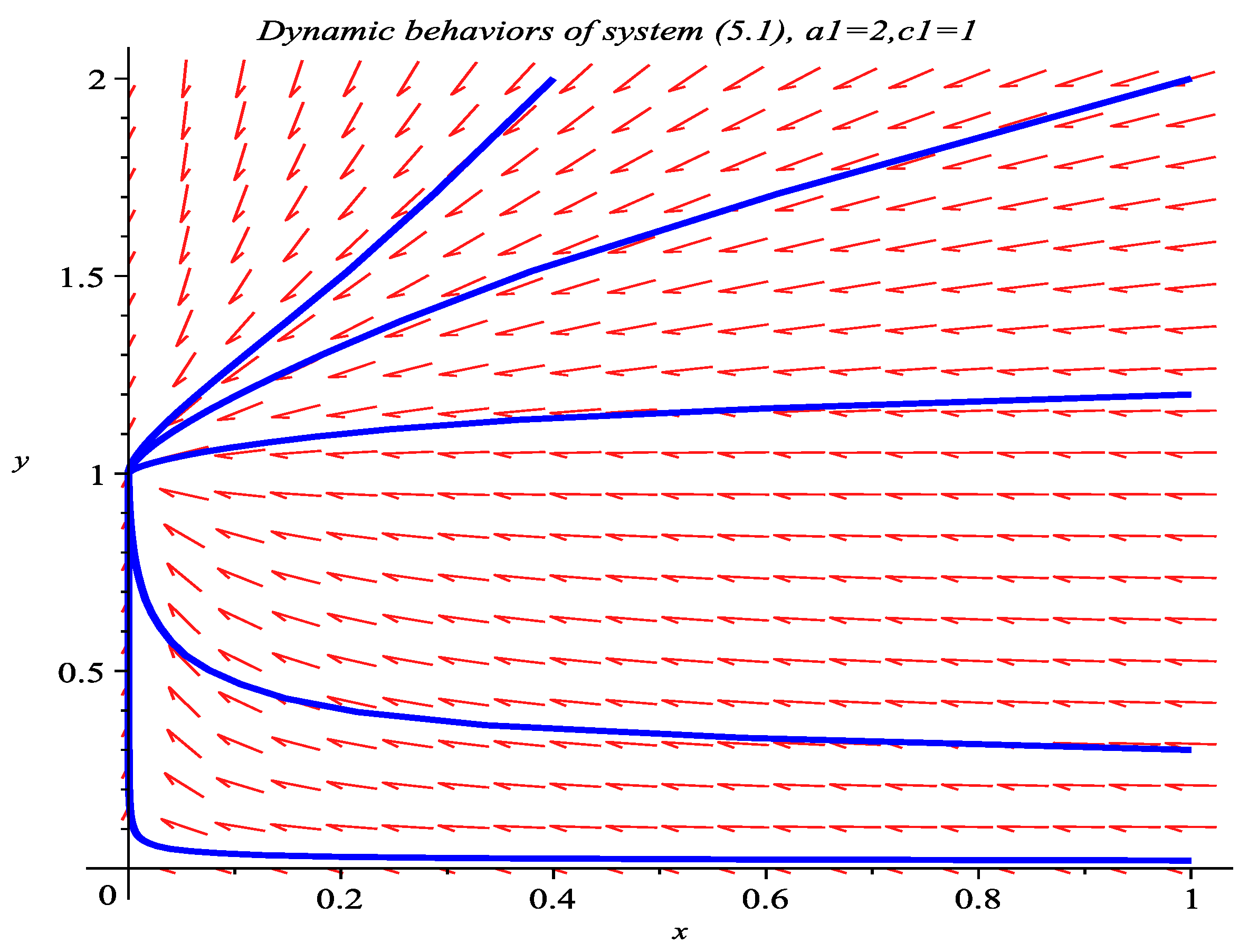
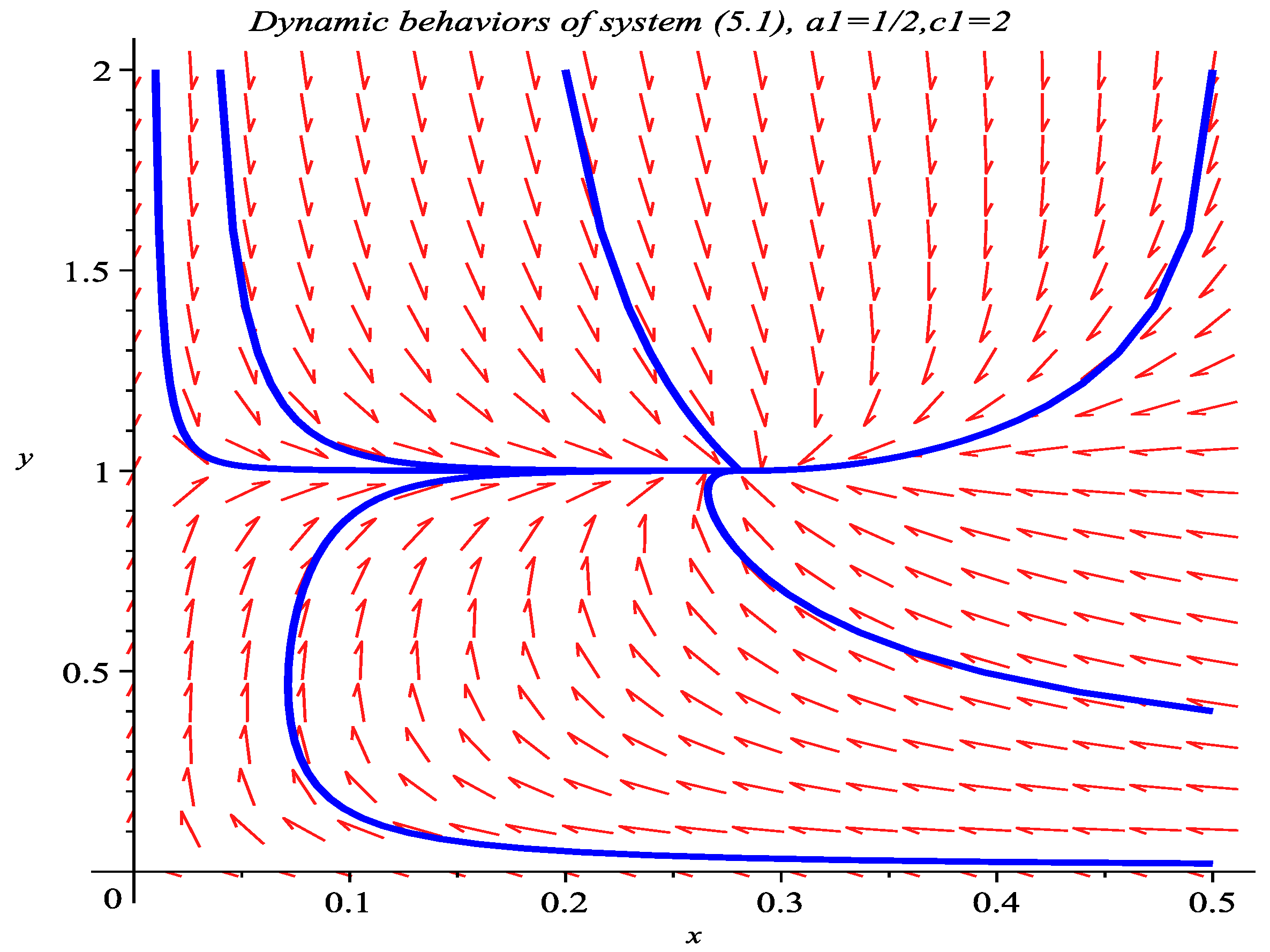
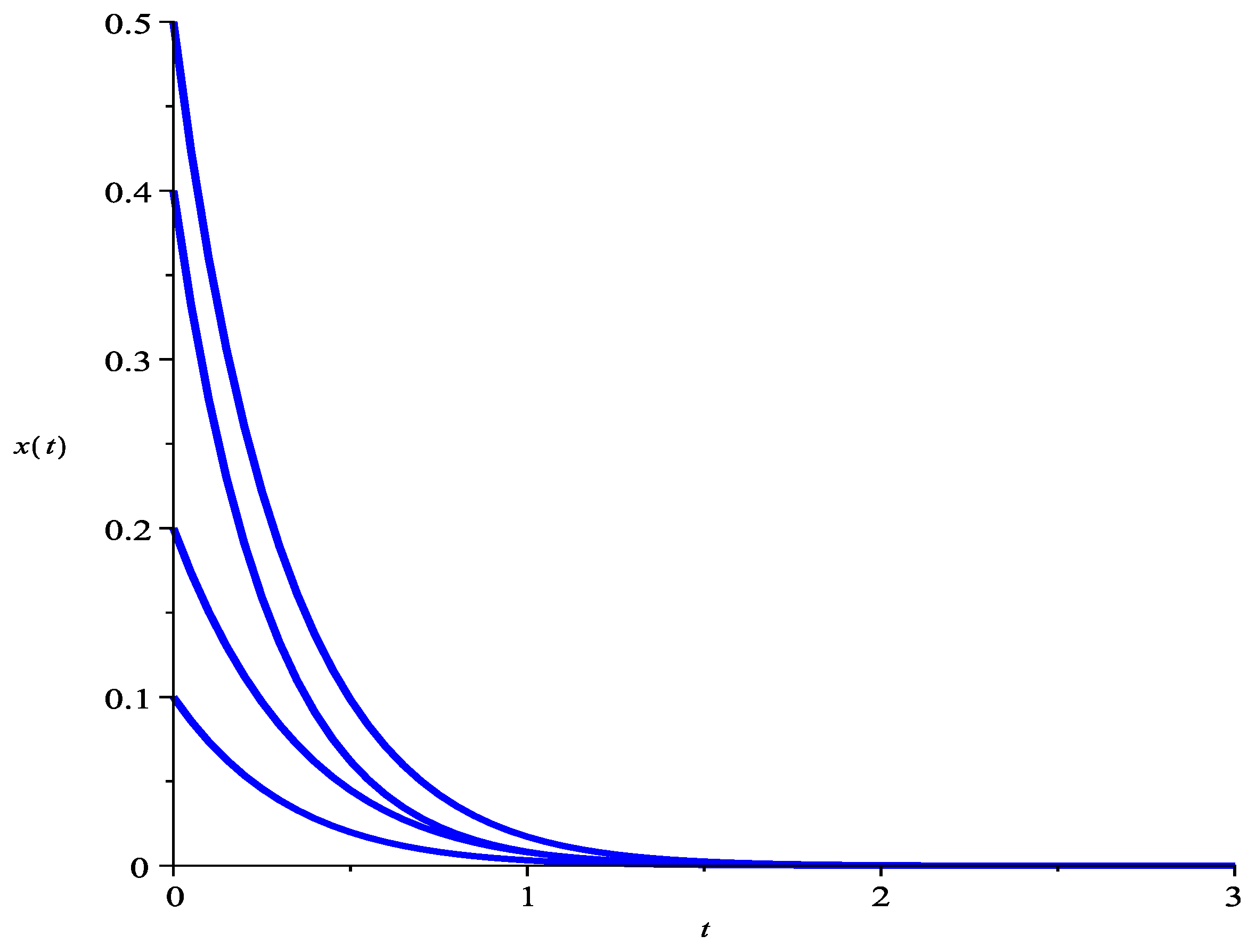
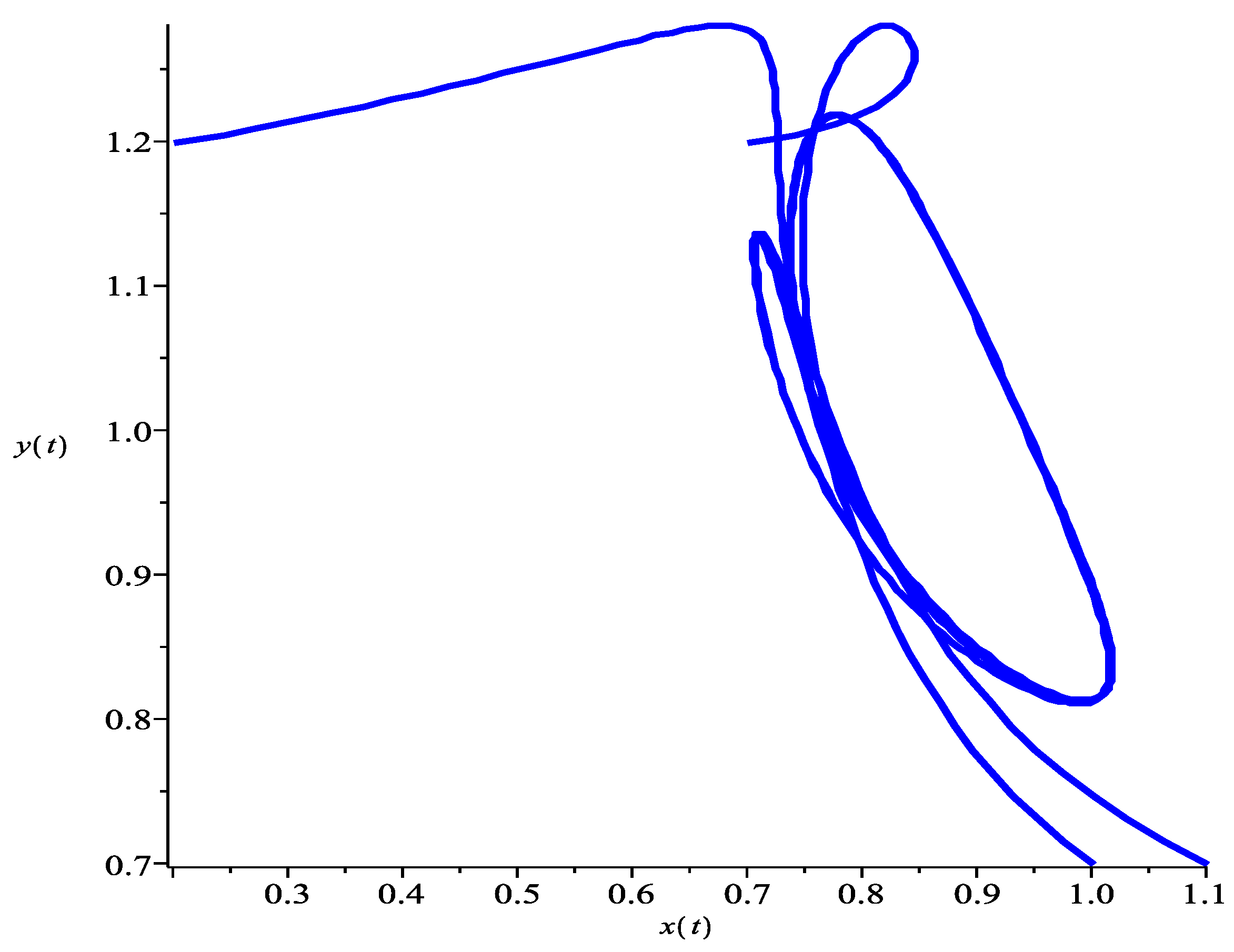
5. Numeric Simulations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, Q.; Chen, F. The influence of partial closure for the populations to a non-selective harvesting Lotka-Volterra discrete amensalism model. Adv. Differ. Equations 2019, 2019, 281. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, F.; Lai, L.; Li, Z. Dynamic behaviors of a discrete May type cooperative system incorporating Michaelis-Menten type harvesting. IAENG Int. J. Appl. Math. 2020, 50, 1–10. [Google Scholar]
- Zhu, Z.; Wu, R.; Chen, F.; Li, Z. Dynamic behaviors of a Lotka-Volterra commensal symbiosis model with non-selective Michaelis-Menten type harvesting. IAENG Int. J. Appl. Math. 2020, 50, 396–404. [Google Scholar]
- Georgescu, P.; Maxin, D.; Zhang, H. Global stability results for models of commensalism. Int. J. Biomath. 2017, 10, 1750037. [Google Scholar] [CrossRef]
- Deng, M. Stability of a stochastic delay commensalism model with Lévy jumps. Phys. A Stat. Mech. Its Appl. 2019, 527, 121061. [Google Scholar] [CrossRef]
- Gakkhar, S.; Gupta, K. A three species dynamical system involving prey-predation, competition and commensalism. Appl. Math. Comput. 2016, 273, 54–67. [Google Scholar] [CrossRef]
- Mougi, A. The roles of amensalistic and commensalistic interactions in large ecological network stability. Sci. Rep. 2016, 6, 29929. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Chen, F.; Xie, X.; Miao, Z. Global stability of May cooperative system with feedback controls. Adv. Differ. Equ. 2015, 2015, 360. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Xia, Y.; Zhang, T. Stability and bifurcation analysis of a commensal model with additive Allee effect and nonlinear growth rate. Int. J. Bifurc. Chaos 2021, 31, 2150204. [Google Scholar] [CrossRef]
- Wu, R.; Li, L.; Zhou, X. A commensal symbiosis model with Holling type functional response. J. Math. Comput. Sci. 2016, 16, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Miao, Z.; Xue, Y. Positive periodic solution of a discrete Lotka-Volterra commensal symbiosis model. Commun. Math. Biol. Neurosci. 2015, 2015, 10. [Google Scholar]
- Chen, B. The influence of commensalism on a Lotka-Volterra commensal symbiosis model with Michaelis-Menten type harvesting. Adv. Differ. Equ. 2019, 2019, 43. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xie, X.; Lin, Q. Permanence, partial survival, extinction, and global attractivity of a nonautonomous harvesting Lotka-Volterra commensalism model incorporating partial closure for the populations. Adv. Differ. Equ. 2018, 2018, 211. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Huang, X. The influence of partial closure for the populations to a harvesting Lotka-Volterra commensalism model. Commun. Math. Biol. Neurosci. 2018, 2018, 10. [Google Scholar]
- Xue, Y.; Xie, X.; Lin, Q. Almost periodic solutions of a commensalism system with Michaelis-Menten type harvesting on time scales. Open Math. 2019, 17, 1503–1514. [Google Scholar] [CrossRef]
- Lei, C. Dynamic behaviors of a stage-structured commensalism system. Adv. Differ. Equ. 2018, 2018, 301. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q. Allee effect increasing the final density of the species subject to the Allee effect in a Lotka-Volterra commensal symbiosis model. Adv. Differ. Equ. 2018, 2018, 196. [Google Scholar] [CrossRef]
- Chen, B. Dynamic behaviors of a commensal symbiosis model involving Allee effect and one party can not survive independently. Adv. Differ. Equ. 2018, 2018, 212. [Google Scholar] [CrossRef]
- Wu, R.; Li, L.; Lin, Q. A Holling type commensal symbiosis model involving Allee effect. Commun. Math. Biol. Neurosci. 2018, 2018, 6. [Google Scholar]
- Lei, C. Dynamic behaviors of a Holling type commensal symbiosis model with the first species subject to Allee effect. Commun. Math. Biol. Neurosci. 2019, 2019, 3. [Google Scholar]
- Vargas-De-León, C.; Gómez-Alcaraz, G. Global stability in some ecological models of commensalism between two species. Biomatemática 2013, 23, 139–146. [Google Scholar]
- Chen, F.; Xue, Y.; Lin, Q.; Xie, X. Dynamic behaviors of a Lotka-Volterra commensal symbiosis model with density dependent birth rate. Adv. Differ. Equ. 2018, 2018, 296. [Google Scholar] [CrossRef]
- Han, R.; Chen, F. Global stability of a commensal symbiosis model with feedback controls. Commun. Math. Biol. Neurosci. 2015, 2015, 15. [Google Scholar]
- Guan, X.; Chen, F. Dynamical analysis of a two species amensalism model with Beddington-DeAngelis functional response and Allee effect on the second species. Nonlinear Anal. Real World Appl. 2019, 48, 71–93. [Google Scholar] [CrossRef]
- Li, T.; Lin, Q.; Chen, J. Positive periodic solution of a discrete commensal symbiosis model with Holling II functional response. Commun. Math. Biol. Neurosci. 2016, 2016, 22. [Google Scholar]
- Puspitasari, N.; Kusumawinahyu, W.M.; Trisilowati, T. Dynamic analysis of the symbiotic model of commensalism and parasitism with harvesting in commensal populations. JTAM (J. Teor. Apl. Mat.) 2021, 5, 193–204. [Google Scholar] [CrossRef]
- Jawad, S. Study the dynamics of commensalism interaction with Michaels-Menten type prey harvesting. Al-Nahrain J. Sci. 2022, 25, 45–50. [Google Scholar] [CrossRef]
- Kumar, G.B.; Srinivas, M.N. Influence of spatiotemporal and noise on dynamics of a two species commensalism model with optimal harvesting. Res. J. Pharm. Technol. 2016, 9, 1717–1726. [Google Scholar] [CrossRef]
- Li, T.; Wang, Q. Stability and Hopf bifurcation analysis for a two-species commensalism system with delay. Qual. Theory Dyn. Syst. 2021, 20, 83. [Google Scholar] [CrossRef]
- Puspitasari, N.; Kusumawinahyu, W.M.; Trisilowati, T. Dynamical analysis of the symbiotic model of commensalism in four populations with Michaelis-Menten type harvesting in the first commensal population. JTAM (J. Teor. Apl. Mat.) 2021, 5, 392–404. [Google Scholar]
- Zhou, Y.C.; Jin, Z.; Qin, J.L. Ordinary Differential Equaiton and Its Application; Science Press: Beijing, China, 2003. [Google Scholar]
- Zhu, Z.; Chen, Y.; Li, Z.; Chen, F. Stability and bifurcation in a Leslie-Gower predator-prey model with Allee effect. Int. J. Bifurc. Chaos 2022, 32, 2250040. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.; Chen, F. Stability and bifurcation in a two-patch model with additive Allee effect. AIMS Math. 2022, 7, 536–551. [Google Scholar] [CrossRef]
- Yang, L.Y.; Han, R.Y.; Xue, Y.L.; Chen, F.D. On a nonautonomous obligate Lotka-Volterra model. J. Sanming Univ. 2014, 31, 15–18. [Google Scholar]
- Chen, F.D.; Lin, C.T.; Yang, L.Y. On a discrete obligate Lotka-Volterra model with one party can not surivive independently. J. Shenyang Univ. (Natural Sci.) 2015, 27, 336–338. [Google Scholar]
- Chen, F.; Pu, L.; Yang, L. Positive periodic solution of a discrete obligate Lotka-Volterra model. Commun. Math. Biol. Neurosci. 2015, 2015, 14. [Google Scholar]
- Wu, R.; Li, L. Dynamic behaviors of a commensal symbiosis model with ratio-dependent functional response and one party can not survive independently. J. Math. Comput. Sci. 2016, 16, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S. The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Entomol. Soc. Can. 1965, 97, 5–60. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Chen, F. Almost periodic solution of a modified Leslie-CGower predator-Cprey model with Holling-type II schemes and mutual interference. Int. J. Biomath. 2014, 7, 1450028. [Google Scholar] [CrossRef]
- Molla, H.; Sarwardi, S.; Sajid, M. Predator-prey dynamics with Allee effect on predator species subject to intra-specific competition and nonlinear prey refuge. J. Math. Comput. Sci. 2022, 25, 150–165. [Google Scholar] [CrossRef]
- Roy, J.; Barman, D.; Alam, S. Role of fear in a predator-prey system with ratio-dependent functional response in deterministic and stochastic environment. Biosystems 2020, 197, 104176. [Google Scholar] [CrossRef]
- Pal, S.; Majhi, S.; Mandal, S.; Pal, N. Role of fear in a predator-prey model with Beddington-DeAngelis functional response. Z. Naturforschung A 2019, 74, 581–595. [Google Scholar] [CrossRef]
- Crowley, P.H.; Martin, E.K. Functional responses and interference within and between year classes of a dragonfly population. J. N. Am. Benthol. Soc. 1989, 8, 211–221. [Google Scholar] [CrossRef]
- Tripathi, J.P.; Bugalia, S.; Tiwari, V.; Kang, Y. A predator-prey model with Crowley-Martin functional response: A nonautonomous study. Nat. Resour. Model. 2020, 33, E12287. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Xue, Y.; Xie, X.; Lin, Q. Dynamic Behaviors of an Obligate Commensal Symbiosis Model with Crowley–Martin Functional Responses. Axioms 2022, 11, 298. https://doi.org/10.3390/axioms11060298
Xu L, Xue Y, Xie X, Lin Q. Dynamic Behaviors of an Obligate Commensal Symbiosis Model with Crowley–Martin Functional Responses. Axioms. 2022; 11(6):298. https://doi.org/10.3390/axioms11060298
Chicago/Turabian StyleXu, Lili, Yalong Xue, Xiangdong Xie, and Qifa Lin. 2022. "Dynamic Behaviors of an Obligate Commensal Symbiosis Model with Crowley–Martin Functional Responses" Axioms 11, no. 6: 298. https://doi.org/10.3390/axioms11060298
APA StyleXu, L., Xue, Y., Xie, X., & Lin, Q. (2022). Dynamic Behaviors of an Obligate Commensal Symbiosis Model with Crowley–Martin Functional Responses. Axioms, 11(6), 298. https://doi.org/10.3390/axioms11060298