Figure 1.
Graphical representation of seed locations. DLPFC, dorsolateral prefrontal cortex (red). vmPFC, ventromedial prefrontal cortex (yellow). dACC, dorsal anterior cingulate cortex (green). Insula (light blue). Striatum (dark blue).
Figure 1.
Graphical representation of seed locations. DLPFC, dorsolateral prefrontal cortex (red). vmPFC, ventromedial prefrontal cortex (yellow). dACC, dorsal anterior cingulate cortex (green). Insula (light blue). Striatum (dark blue).
Figure 2.
Differences in VAS-F scores. The VAS-F raw scores are plotted as a function of group, condition, and rating. 0-back condition is denoted by circles, and the 2-back condition is denoted by triangles. Red represents the HC group, and turquoise represents the MS group. Abbreviations: VAS-F, Visual Analogue Scale of Fatigue. MS, multiple sclerosis. HC, healthy control.
Figure 2.
Differences in VAS-F scores. The VAS-F raw scores are plotted as a function of group, condition, and rating. 0-back condition is denoted by circles, and the 2-back condition is denoted by triangles. Red represents the HC group, and turquoise represents the MS group. Abbreviations: VAS-F, Visual Analogue Scale of Fatigue. MS, multiple sclerosis. HC, healthy control.
Figure 3.
Group × condition × VAS-F (transformed) interaction for n-back task accuracy. For the 2-back condition, there was a positive correlation between fatigue levels and accuracy in the HC group and a negative correlation in the MS group. There were no significant relationships between fatigue levels and accuracy for the 0-back condition. 0-back condition is denoted by dotted lines, and the 2-back condition is denoted by solid lines. Abbreviations: MS, multiple sclerosis. HC, healthy control. VAS-F, Visual Analogue Scale of Fatigue.
Figure 3.
Group × condition × VAS-F (transformed) interaction for n-back task accuracy. For the 2-back condition, there was a positive correlation between fatigue levels and accuracy in the HC group and a negative correlation in the MS group. There were no significant relationships between fatigue levels and accuracy for the 0-back condition. 0-back condition is denoted by dotted lines, and the 2-back condition is denoted by solid lines. Abbreviations: MS, multiple sclerosis. HC, healthy control. VAS-F, Visual Analogue Scale of Fatigue.
Figure 4.
Dorsolateral prefrontal cortex connectivity 3-dimensional rendering. (a) Task-independent, fatigue-related connectivity in the healthy control group; (b) Task-independent, fatigue-related connectivity in the multiple sclerosis group. The seed is denoted by green spheres; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellar regions were omitted. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: SFG, superior frontal gyrus. MFG, middle frontal gyrus. IFG, inferior frontal gyrus. vmPFC, ventromedial prefrontal cortex. SMG, superior medial gyrus. PrG, precentral gyrus. PoG, postcentral gyrus. SOG, superior orbital gyrus. SPL, superior parietal lobule. AG, angular gyrus.
Figure 4.
Dorsolateral prefrontal cortex connectivity 3-dimensional rendering. (a) Task-independent, fatigue-related connectivity in the healthy control group; (b) Task-independent, fatigue-related connectivity in the multiple sclerosis group. The seed is denoted by green spheres; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellar regions were omitted. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: SFG, superior frontal gyrus. MFG, middle frontal gyrus. IFG, inferior frontal gyrus. vmPFC, ventromedial prefrontal cortex. SMG, superior medial gyrus. PrG, precentral gyrus. PoG, postcentral gyrus. SOG, superior orbital gyrus. SPL, superior parietal lobule. AG, angular gyrus.
Figure 5.
Ventromedial prefrontal cortex connectivity 3-dimensional rendering. (a) Task-independent, fatigue-related connectivity in the healthy control group. (b) Task-independent, fatigue-related connectivity in the multiple sclerosis group. The seed is denoted by green spheres; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellar regions were omitted. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: SFG, superior frontal gyrus. MFG, middle frontal gyrus. IFG, inferior frontal gyrus. vmPFC, ventromedial prefrontal cortex. MCC, middle cingulate cortex. SMA, supplementary motor area. SOG, superior orbital gyrus. MOG, middle orbital gyrus. SMG, supramarginal gyrus. STG, superior temporal gyrus. MTG, middle temporal gyrus. ITG, inferior temporal gyrus. MOcG, middle occipital gyrus. IOcG, inferior occipital gyrus.
Figure 5.
Ventromedial prefrontal cortex connectivity 3-dimensional rendering. (a) Task-independent, fatigue-related connectivity in the healthy control group. (b) Task-independent, fatigue-related connectivity in the multiple sclerosis group. The seed is denoted by green spheres; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellar regions were omitted. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: SFG, superior frontal gyrus. MFG, middle frontal gyrus. IFG, inferior frontal gyrus. vmPFC, ventromedial prefrontal cortex. MCC, middle cingulate cortex. SMA, supplementary motor area. SOG, superior orbital gyrus. MOG, middle orbital gyrus. SMG, supramarginal gyrus. STG, superior temporal gyrus. MTG, middle temporal gyrus. ITG, inferior temporal gyrus. MOcG, middle occipital gyrus. IOcG, inferior occipital gyrus.
![Diagnostics 10 00930 g005]()
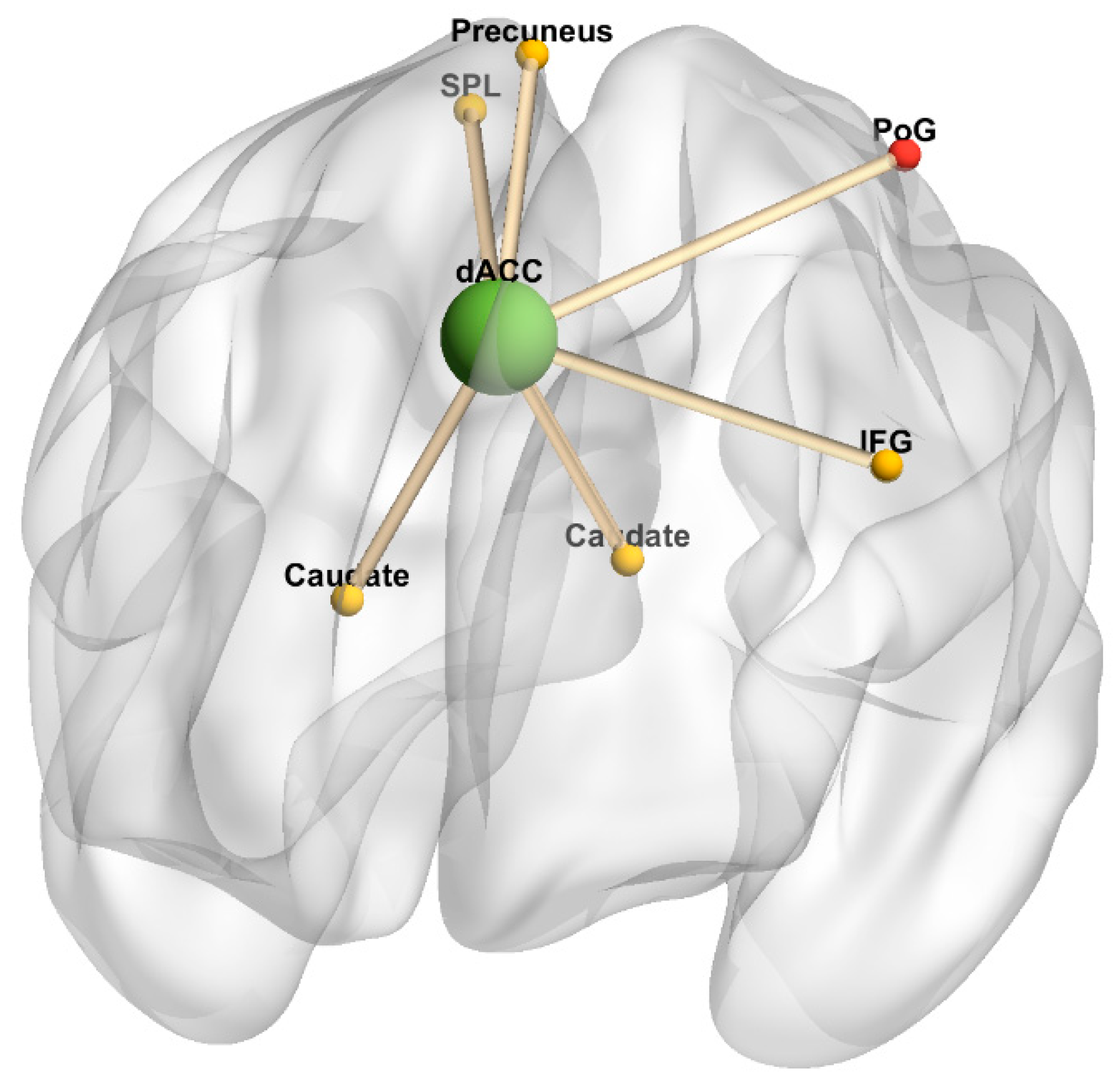
Figure 6.
Dorsal anterior cingulate cortex connectivity 3-dimensional rendering. Figure represents task-independent, fatigue-related connectivity in the multiple sclerosis group; there were no significant differences between conditions in the healthy control group. The seed is denoted by a green sphere; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellum was omitted. Anterior orientation is in the front of the figure, right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: dACC, dorsal anterior cingulate cortex. IFG, inferior frontal gyrus. PoG, postcentral gyrus. SPL, superior parietal lobule.
Figure 6.
Dorsal anterior cingulate cortex connectivity 3-dimensional rendering. Figure represents task-independent, fatigue-related connectivity in the multiple sclerosis group; there were no significant differences between conditions in the healthy control group. The seed is denoted by a green sphere; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellum was omitted. Anterior orientation is in the front of the figure, right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: dACC, dorsal anterior cingulate cortex. IFG, inferior frontal gyrus. PoG, postcentral gyrus. SPL, superior parietal lobule.
Figure 7.
Insula connectivity 3-dimensional rendering. (a) Task-independent, fatigue-related connectivity in the healthy control group. (b) Task-independent, fatigue-related connectivity in the multiple sclerosis group. The seed is denoted by green spheres; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellar regions were omitted. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: IFG, inferior frontal gyrus. vmPFC, ventromedial prefrontal cortex. SMG, superior medial gyrus. MCC, middle cingulate cortex. PcL, paracentral lobule. PoG, postcentral gyrus. MOG, middle orbital gyrus. STG, superior temporal gyrus. SPL, superior parietal lobule. SOcG, superior occipital gyrus. MOcG, middle occipital gyrus. IOcG, inferior occipital gyrus.
Figure 7.
Insula connectivity 3-dimensional rendering. (a) Task-independent, fatigue-related connectivity in the healthy control group. (b) Task-independent, fatigue-related connectivity in the multiple sclerosis group. The seed is denoted by green spheres; regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres, and regions with increased connectivity to the seed in the 0-back condition are denoted by red spheres. Cerebellar regions were omitted. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: IFG, inferior frontal gyrus. vmPFC, ventromedial prefrontal cortex. SMG, superior medial gyrus. MCC, middle cingulate cortex. PcL, paracentral lobule. PoG, postcentral gyrus. MOG, middle orbital gyrus. STG, superior temporal gyrus. SPL, superior parietal lobule. SOcG, superior occipital gyrus. MOcG, middle occipital gyrus. IOcG, inferior occipital gyrus.
![Diagnostics 10 00930 g007]()
Figure 8.
Striatum connectivity 3-dimensional rendering. Figure represents task-independent, fatigue-related connectivity in the multiple sclerosis group; there were no significant differences between conditions in the healthy control group. The seed is denoted by a green sphere, and regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres; there were no regions with increased connectivity to the seed in the 0-back condition. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: SPL, superior parietal lobule. AG, angular gyrus.
Figure 8.
Striatum connectivity 3-dimensional rendering. Figure represents task-independent, fatigue-related connectivity in the multiple sclerosis group; there were no significant differences between conditions in the healthy control group. The seed is denoted by a green sphere, and regions with increased connectivity to the seed in the 2-back condition are denoted by yellow spheres; there were no regions with increased connectivity to the seed in the 0-back condition. Anterior orientation is in the front of the figure; right hemisphere is on the left side, and left hemisphere is on the right side. Abbreviations: SPL, superior parietal lobule. AG, angular gyrus.
Table 1.
Coordinates of center location of seed regions.
Table 1.
Coordinates of center location of seed regions.
| Location | X | Y | Z |
|---|
| DLPFC | 44 | 32 | 36 |
| vmPFC | −6 | 46 | −6 |
| dACC | −4 | 20 | 46 |
| Insula | 34 | 22 | 0 |
| Striatum | 18 | 12 | 0 |
Table 2.
Demographic and disease characteristics.
Table 2.
Demographic and disease characteristics.
| Characteristic | MS (n = 26) | HC (n = 14) | MS vs. HC (p) |
|---|
| Mean (SD) | Range | Mean (SD) | Range |
|---|
| Age: years | 48.12 (10.09) | 26–63 | 42 (12.87) | 24–58 | n.s. |
| Education: years | 16.19 (2.19) | 12–21 | 16.31 (1.97) | 12–20 | n.s. |
| Illness duration: years | 11.05 (6.21) | 1–28 | - | - | - |
| | Number (%) | | Number (%) | | |
| Females | 23 (88) | - | 13 (93) | - | n.s. |
| MS phenotype | - | - | - | - | - |
| Relapsing-remitting | 19 (73) | - | - | - | - |
| Primary progressive | 2 (8) | - | - | - | - |
| Secondary progressive | 3 (12) | - | - | - | - |
| Unknown | 2 (7) | - | - | - | - |
Table 3.
Task-independent (residual) fatigue-related functional connectivity with the dorsolateral prefrontal cortex as seed.
Table 3.
Task-independent (residual) fatigue-related functional connectivity with the dorsolateral prefrontal cortex as seed.
| Dorsolateral Prefrontal Cortex Seed |
|---|
| Location | X | Y | Z | Voxels | Z Stat |
|---|
| HC |
| Increased Task-Independent Residual Connectivity in 2-back Condition |
| Superior Frontal Gyrus | −30.7 | −4.8 | 70 | 19 | 4.90 |
| Inferior Frontal Gyrus | −54.7 | 22.7 | 22 | 103 | 4.96 |
| Increased Task-Independent Residual Connectivity in 0-back Condition |
| Middle Frontal Gyrus | 27.8 | 39.9 | 46 | 27 | −4.77 |
| Ventromedial Prefrontal Cortex | 14.0 | 12.4 | −18 | 30 | −5.13 |
| Precuneus | 14.0 | −63.2 | 34 | 36 | −4.30 |
| Cuneus | 0.3 | −94.2 | 42 | 13 | −5.05 |
| MS |
| Increased Task-Independent Residual Connectivity in 2-back Condition |
| Superior Frontal Gyrus | −16.9 | 70.8 | 18 | 14 | 3.77 |
| Middle Frontal Gyrus | 27.8 | 39.9 | 30 | 14 | 4.72 |
| Middle Frontal Gyrus | −41.0 | 43.3 | 30 | 29 | 5.56 |
| Inferior Frontal Gyrus | 62.1 | 12.4 | 14 | 16 | 3.91 |
| Inferior Frontal Gyrus | 58.7 | 39.9 | 10 | 25 | 4.72 |
| Superior Medial Gyrus | 0.3 | 53.7 | 22 | 61 | 6.21 |
| Precentral Gyrus | 38.1 | 2.1 | 34 | 15 | 5.00 |
| Superior Parietal Lobule | 20.9 | −80.4 | 50 | 35 | 5.80 |
| Angular Gyrus | −37.6 | −70.1 | 54 | 18 | 4.66 |
| Increased Task-Independent Residual Connectivity in 0-back Condition |
| Superior Frontal Gyrus | −23.8 | 9.0 | 70 | 54 | −6.26 |
| Middle Frontal Gyrus | 45.0 | 9.0 | 58 | 15 | −4.36 |
| Superior Orbital Gyrus | 7.1 | 60.5 | −22 | 24 | −4.73 |
| Precentral Gyrus | −41.0 | −11.7 | 70 | 13 | −4.59 |
| Postcentral Gyrus | 34.6 | −28.9 | 78 | 21 | −4.43 |
| Postcentral Gyrus/Superior Parietal Lobule | 45.0 | −42.6 | 66 | 42 | −4.53 |
| Superior Parietal Lobule | −37.6 | −70.1 | 66 | 14 | −6.01 |
| Hippocampus | −16.9 | −35.7 | 14 | 15 | −4.75 |
| Cerebellum Crus I | −51.3 | −49.5 | −38 | 45 | −6.36 |
| Cerebellum Crus I | −37.6 | −90.7 | −30 | 15 | −4.51 |
Table 4.
Task-independent (residual) fatigue-related functional connectivity with the ventromedial prefrontal cortex as seed.
Table 4.
Task-independent (residual) fatigue-related functional connectivity with the ventromedial prefrontal cortex as seed.
| Ventromedial Prefrontal Cortex Seed |
|---|
| Location | X | Y | Z | Voxels | Z Stat |
|---|
| HC |
| Increased Task-independent Residual Connectivity in 2-back Condition |
| Middle Orbital Gyrus | −10.1 | 60.5 | −10 | 110 | 5.45 |
| Inferior Frontal Gyrus | −41.0 | 36.5 | −18 | 68 | 5.45 |
| Middle Cingulate Cortex | −3.2 | −25.4 | 46 | 30 | 4.84 |
| Superior Temporal Gyrus/Insula | −47.9 | 5.5 | −10 | 21 | 4.78 |
| Superior Temporal Gyrus | 69.0 | −28.9 | 22 | 20 | 4.72 |
| Putamen | −20.4 | −1.4 | −6 | 16 | 5.11 |
| Cerebellar Vermis | 7.1 | −77.0 | −18 | 20 | 5.29 |
| Increased Task-independent Residual Connectivity in 0-back Condition |
| Superior Frontal Gyrus/Superior Orbital Gyrus | −27.2 | 67.4 | −2 | 19 | −5.10 |
| Middle Frontal Gyrus | −47.9 | 36.5 | 34 | 208 | −6.12 |
| Middle Frontal Gyrus | 34.6 | 15.8 | 62 | 18 | −4.64 |
| Inferior Frontal Gyrus | −20.4 | 26.1 | −22 | 24 | −4.32 |
| Temporal Pole | −54.7 | 19.3 | −22 | 22 | −4.26 |
| Middle Temporal Gyrus | −58.2 | 2.1 | −26 | 27 | −4.06 |
| Inferior Temporal Gyrus | 58.7 | −39.2 | −18 | 19 | −4.00 |
| MS |
| Increased Task-independent Residual Connectivity in 2-back Condition |
| Inferior Frontal Gyrus | 45.0 | 12.4 | 6 | 40 | 5.20 |
| Inferior Frontal Gyrus | 38.1 | 9.0 | 30 | 21 | 4.42 |
| Supplemental Motor Area | 14.0 | 12.4 | 66 | 39 | 5.10 |
| Insula | −37.6 | 19.3 | 6 | 13 | 4.43 |
| Middle Temporal Gyrus | 65.6 | −49.5 | 2 | 31 | 4.14 |
| Inferior Temporal Gyrus | −61.6 | −56.4 | −6 | 13 | 4.15 |
| Supramarginal Gyrus | 58.7 | −25.4 | 46 | 31 | 4.30 |
| Middle Occipital Gyrus | 34.6 | −63.2 | 34 | 53 | 5.42 |
| Cerebellum Crus I | −51.3 | −49.5 | −30 | 37 | 5.38 |
| Increased Task-independent Residual Connectivity in 0-back Condition |
| Inferior Temporal Gyrus | 31.2 | 2.1 | −50 | 20 | −5.51 |
| Inferior Occipital Gyrus | −54.7 | −73.6 | −18 | 21 | −4.66 |
Table 5.
Task-independent (residual) fatigue-related functional connectivity with the dorsal anterior cingulate cortex as seed.
Table 5.
Task-independent (residual) fatigue-related functional connectivity with the dorsal anterior cingulate cortex as seed.
| Dorsal Anterior Cingulate Cortex Seed |
|---|
| Location | X | Y | Z | Voxels | Z Stat |
|---|
| MS |
| Increased Task-independent Residual Connectivity in 2-back Condition |
| Inferior Frontal Gyrus | −47.9 | 5.5 | 30 | 50 | 4.64 |
| Superior Parietal Lobule | 24.3 | −73.6 | 58 | 43 | 4.82 |
| Precuneus | 7.1 | −63.2 | 66 | 30 | 4.63 |
| Caudate Nucleus | −16.9 | 9.0 | 18 | 24 | 4.14 |
| Caudate Nucleus | 14.0 | 22.7 | 14 | 18 | 4.38 |
| Increased Task-independent Residual Connectivity in 0-back Condition |
| Postcentral Gyrus | −41.0 | −28.9 | 62 | 46 | −5.34 |
| Cerebellum Crus 1 | 41.5 | −83.9 | −34 | 16 | −4.98 |
Table 6.
Task-independent (residual) fatigue-related functional connectivity with the insula as seed.
Table 6.
Task-independent (residual) fatigue-related functional connectivity with the insula as seed.
| Insula Seed |
|---|
| Location | X | Y | Z | Voxels | Z Stat |
|---|
| HC |
| Increased Task-independent Residual Connectivity in 2-back Condition |
| Inferior Frontal Gyrus | −51.3 | 26.1 | −2 | 51 | 5.45 |
| Inferior Frontal Gyrus | −47.9 | 22.7 | 30 | 18 | 3.81 |
| Middle Orbital Gyrus | −6.6 | 70.8 | −14 | 15 | 4.18 |
| Superior Temporal Gyrus | 55.3 | −25.4 | 2 | 18 | 4.96 |
| Increased Task-independent Residual Connectivity in 0-back Condition |
| Middle Cingulate Cortex | 14.0 | −32.3 | 42 | 20 | −4.47 |
| Paracentral Lobule | −10.1 | −39.2 | 82 | 13 | −5.33 |
| Postcentral Gyrus | −41.0 | −46.1 | 66 | 17 | −4.84 |
| Superior Parietal Lobule | −30.7 | −42.6 | 74 | 22 | −4.46 |
| Precuneus | 7.1 | −63.2 | 66 | 51 | −4.60 |
| Superior Occipital Gyrus | 24.3 | −70.1 | 46 | 16 | −3.95 |
| Middle Occipital Gyrus | −37.6 | −77.0 | 34 | 34 | 5.46 |
| Calcarine Gyrus | 20.9 | −77.0 | 14 | 13 | −4.60 |
| Cerebellum lobule VIII | 24.3 | −35.7 | −50 | 22 | −4.74 |
| MS |
| Increased Task-independent Residual Connectivity in 2-back Condition |
| Middle Orbital Gyrus | 0.3 | 64.0 | −10 | 68 | 5.09 |
| Superior Medial Gyrus | −6.6 | 57.1 | 46 | 21 | 5.11 |
| Insula | 45.0 | 5.5 | −6 | 26 | 4.24 |
| Caudate Nucleus | 10.6 | 12.4 | 18 | 13 | 5.91 |
| Increased Task-independent Residual Connectivity in 0-back Condition |
| Ventromedial Prefrontal Cortex | −6.6 | 5.5 | −22 | 15 | −6.21 |
| Fusiform Gyrus | 27.8 | −66.7 | −6 | 13 | −4.09 |
| Inferior Occipital Gyrus | 31.2 | −87.3 | −2 | 19 | −3.70 |
| Calcarine Gyrus | 3.7 | −70.1 | 18 | 15 | −4.03 |
| Cuneus | 14.0 | −73.6 | 30 | 26 | −5.39 |
| Lingual Gyrus | −6.6 | −83.9 | −2 | 39 | −4.88 |
| Cerebellum Crus I | −54.7 | −56.4 | −38 | 15 | −4.62 |
| Cerebellum Lobule VI | −20.4 | −66.7 | −14 | 14 | −4.85 |
| Cerebellum Lobule VIII | −16.9 | −63.2 | −62 | 26 | −4.46 |
Table 7.
Task-independent (residual) fatigue-related functional connectivity with the striatum as seed.
Table 7.
Task-independent (residual) fatigue-related functional connectivity with the striatum as seed.
| Striatum Seed |
|---|
| Location | X | Y | Z | Voxels | Z Stat |
|---|
| MS |
| Increased Task-independent Residual Connectivity in 2-back Condition |
| Superior Parietal Lobule | −30.7 | −80.4 | 58 | 16 | 4.73 |
| Angular Gyrus/Superior Parietal Lobule | 41.5 | −80.4 | 42 | 16 | 4.58 |

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}