
Identification of piRNA Targets in Urinary Extracellular Vesicles for the Diagnosis of Prostate Cancer

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Isolation and Confirmation of Urinary EVs
2.3. EVs RNA Extraction
2.4. Transmission Electron Microscopy
2.5. Western Blot Analysis
2.6. Small RNA Sequencing and Sequencing Data Analysis
2.7. Reverse Transcription and RT-qPCR of Urinary EVs piRNA
2.8. Identification of Key piRNAs Potential Downstream Mechanisms
2.9. Statistical Analysis
3. Results
3.1. Patient Clinical Details
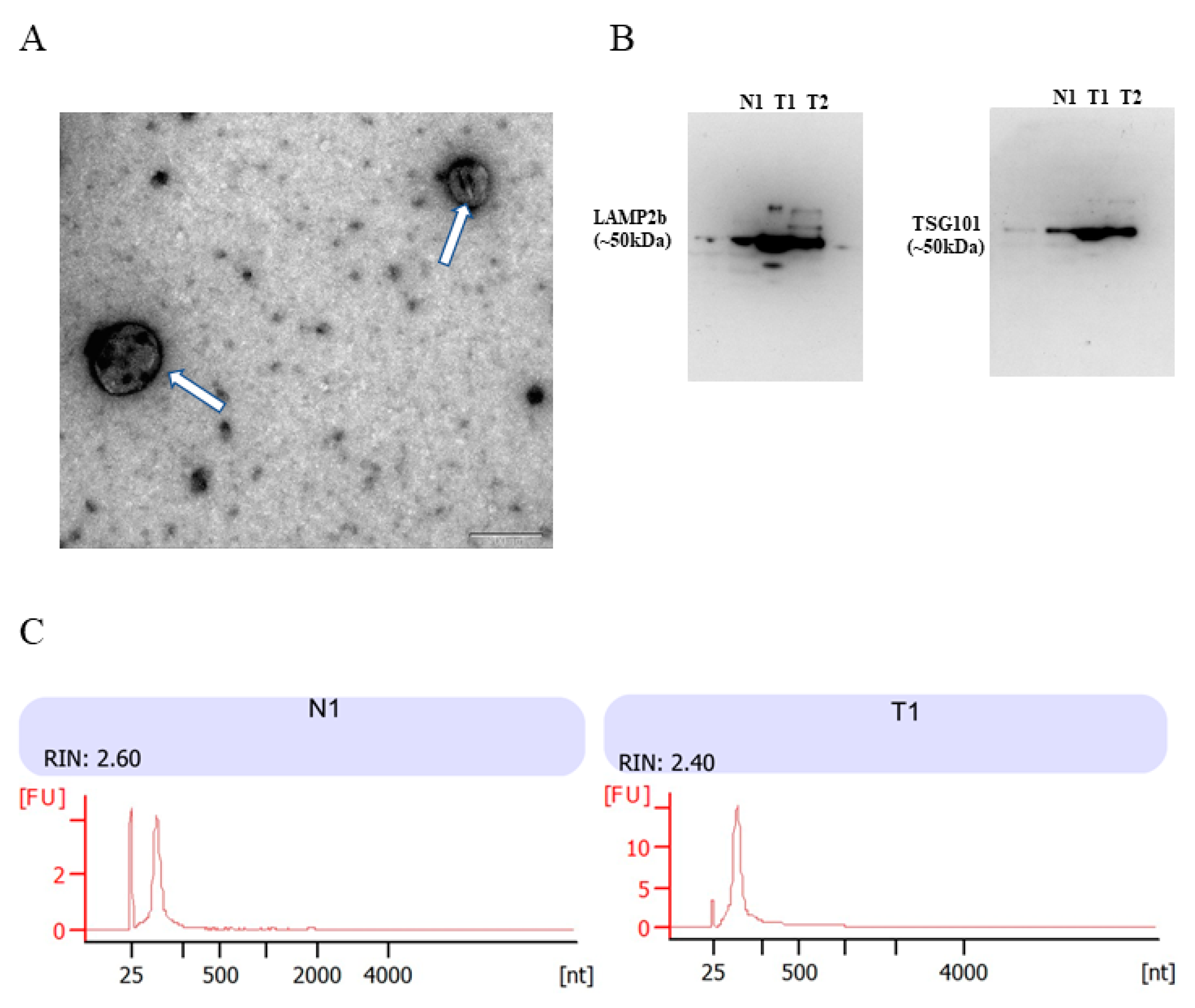
3.2. Identification of Urinary Cell-Free EVs and Characterization of Urinary EVs RNA
3.3. Small RNA Sequencing of Urinary Cell-Free EVs RNA
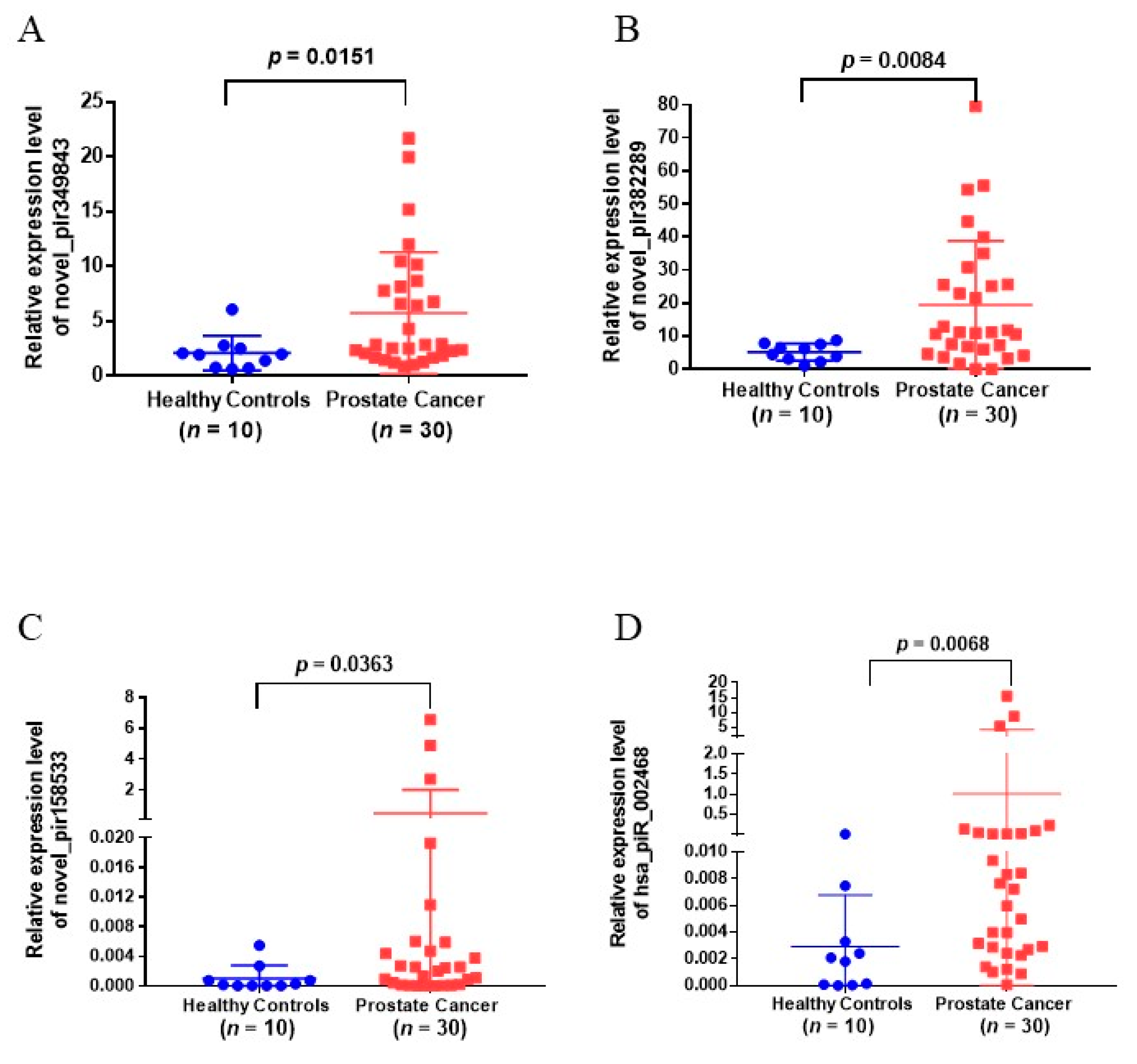
3.4. Validation of Urinary EVs piRNAs in Another Cohort of Patients
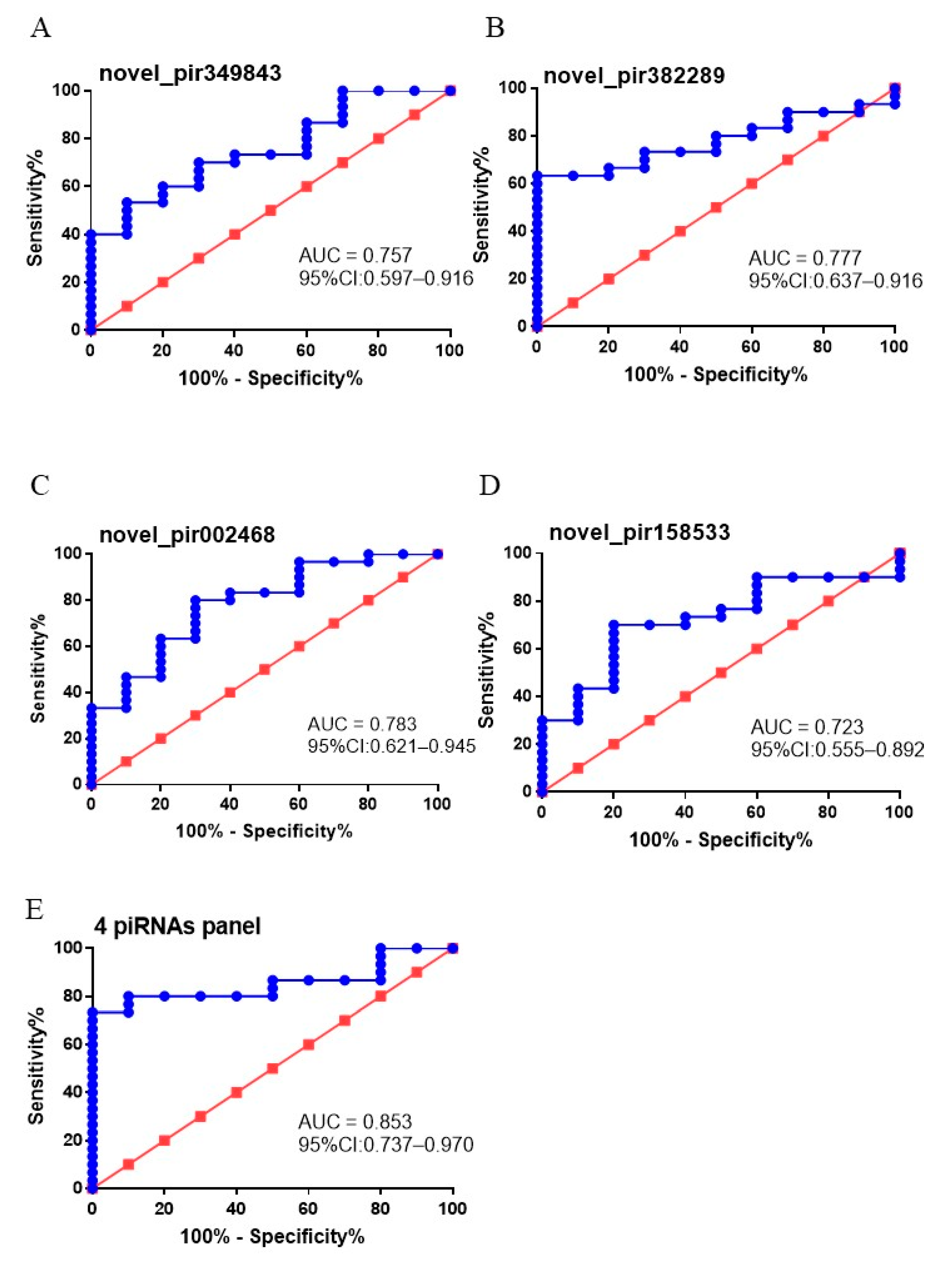
3.5. The Diagnostic Value of Putative EVs piRNA Biomarkers
3.6. Identification of the Potential Downstream Mechanisms of the Four Dysregulated piRNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Ee, I.B.; Hagedoorn, M.; Smits, C.H.M.; Kamper, A.M.; Honkoop, H.A.; Slaets, J.P.J. This is an older men’s world: A qualitative study of men’s experiences with prostate cancer. Eur. J. Oncol. Nurs. 2018, 37, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Jean-Pierre, G. Advice about Screening for Prostate Cancer With Prostate-Specific Antigen. J. Adv. Pract. Oncol. 2017, 8, 639–645. [Google Scholar] [PubMed]
- Crowley, E.; Di Nicolantonio, F.; Loupakis, F.; Bardelli, A. Liquid biopsy: Monitoring cancer-genetics in the blood. Nat. Rev. Clin. Oncol. 2013, 10, 472–484. [Google Scholar] [CrossRef]
- Tkach, M.; Thery, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [Green Version]
- Lorenc, T.; Klimczyk, K.; Michalczewska, I.; Slomka, M.; Kubiak-Tomaszewska, G.; Olejarz, W. Exosomes in Prostate Cancer Diagnosis, Prognosis and Therapy. Int. J. Mol. Sci. 2020, 21, 2118. [Google Scholar] [CrossRef] [Green Version]
- Ramalho-Carvalho, J.; Fromm, B.; Henrique, R.; Jeronimo, C. Deciphering the function of non-coding RNAs in prostate cancer. Cancer Metastasis Rev. 2016, 35, 235–262. [Google Scholar] [CrossRef]
- Mattick, J.S. Non-coding RNAs: The architects of eukaryotic complexity. EMBO Rep. 2001, 2, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Kirino, Y.; Mourelatos, Z. Mouse Piwi-interacting RNAs are 2′-O-methylated at their 3′ termini. Nat. Struct. Mol. Biol. 2007, 14, 347–348. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Sakaguchi, Y.; Suzuki, T.; Ueda, H.; Miyauchi, K.; Suzuki, T. The 3′ termini of mouse Piwi-interacting RNAs are 2′-O-methylated. Nat. Struct. Mol. Biol. 2007, 14, 349–350. [Google Scholar] [CrossRef]
- Liu, Y.; Dou, M.; Song, X.; Dong, Y.; Liu, S.; Liu, H.; Tao, J.; Li, W.; Yin, X.; Xu, W. The emerging role of the piRNA/piwi complex in cancer. Mol. Cancer 2019, 18, 123. [Google Scholar] [CrossRef] [Green Version]
- Ku, H.Y.; Lin, H. PIWI proteins and their interactors in piRNA biogenesis, germline development and gene expression. Natl. Sci. Rev. 2014, 1, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Chalbatani, G.M.; Dana, H.; Memari, F.; Gharagozlou, E.; Ashjaei, S.; Kheirandish, P.; Marmari, V.; Mahmoudzadeh, H.; Mozayani, F.; Maleki, A.R.; et al. Biological function and molecular mechanism of piRNA in cancer. Pract. Lab. Med. 2019, 13, e00113. [Google Scholar] [CrossRef]
- Kurth, H.M.; Mochizuki, K. 2′-O-methylation stabilizes Piwi-associated small RNAs and ensures DNA elimination in Tetrahymena. RNA 2009, 15, 675–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Hong, J.; Hong, M.; Wang, Y.; Yu, T.; Zang, S.; Wu, Q. piRNA-823 delivered by multiple myeloma-derived extracellular vesicles promoted tumorigenesis through re-educating endothelial cells in the tumor environment. Oncogene 2019, 38, 5227–5238. [Google Scholar] [CrossRef] [PubMed]
- Vychytilova-Faltejskova, P.; Stitkovcova, K.; Radova, L.; Sachlova, M.; Kosarova, Z.; Slaba, K.; Kala, Z.; Svoboda, M.; Kiss, I.; Vyzula, R.; et al. Circulating PIWI-Interacting RNAs piR-5937 and piR-28876 Are Promising Diagnostic Biomarkers of Colon Cancer. Cancer Epidemiol. Prev. Biomark. 2018, 27, 1019–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, N.; Zhang, F.; Jin, S.; Dong, Y.; Dong, X.; Chen, Y.; Kong, X.; Tong, Y.; Mi, Q.; et al. PIWI-interacting RNAs are aberrantly expressed and may serve as novel biomarkers for diagnosis of lung adenocarcinoma. Thorac. Cancer 2021, 12, 2468–2477. [Google Scholar] [CrossRef]
- Itou, D.; Shiromoto, Y.; Yukiho, S.Y.; Ishii, C.; Nishimura, T.; Ogonuki, N.; Ogura, A.; Hasuwa, H.; Fujihara, Y.; Kuramochi-Miyagawa, S.; et al. Induction of DNA methylation by artificial piRNA production in male germ cells. Curr. Biol. 2015, 25, 901–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Wang, Q.; Jiang, W.; Bian, Y.; Zhou, Y.; Gou, A.; Zhang, W.; Fu, K.; Shi, W. Emerging roles of piRNAs in cancer: Challenges and prospects. Aging (Albany NY) 2019, 11, 9932–9946. [Google Scholar] [CrossRef]
- Qi, T.; Cao, H.; Sun, H.; Feng, H.; Li, N.; Wang, C.; Wang, L. piR-19166 inhibits migration and metastasis through CTTN/MMPs pathway in prostate carcinoma. Aging (Albany NY) 2020, 12, 18209–18220. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, X.; Li, D.; Han, X. piR-001773 and piR-017184 promote prostate cancer progression by interacting with PCDH9. Cell Signal. 2020, 76, 109780. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Clark, D.; Mao, L. Novel dimensions of piRNAs in cancer. Cancer Lett. 2013, 336, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.Y.; Shin, H.; Moon, H.W.; Park, Y.H.; Park, J.; Lee, J.Y. Urinary exosomal microRNA profiling in intermediate-risk prostate cancer. Sci. Rep. 2021, 11, 7355. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Bajo-Santos, C.; Hessvik, N.P.; Lorenz, S.; Fromm, B.; Berge, V.; Sandvig, K.; Line, A.; Llorente, A. Identification of non-invasive miRNAs biomarkers for prostate cancer by deep sequencing analysis of urinary exosomes. Mol. Cancer 2017, 16, 156. [Google Scholar] [CrossRef]
- Goh, W.S.; Falciatori, I.; Tam, O.H.; Burgess, R.; Meikar, O.; Kotaja, N.; Hammell, M.; Hannon, G.J. piRNA-directed cleavage of meiotic transcripts regulates spermatogenesis. Genes Dev. 2015, 29, 1032–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Ji, G.; Huang, R.; Chen, H.; Gao, Y.; Wang, W.; Sun, X.; Zhang, J.; Zheng, J.; Wei, Q. PIWI-interacting RNAs piR-13643 and piR-21238 are promising diagnostic biomarkers of papillary thyroid carcinoma. Aging (Albany NY) 2020, 12, 9292–9310. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.F.; Yeung, R.; Chiu, P.K.; Lam, N.Y.; Chow, J.; Chan, B. The role of urine prostate cancer antigen 3 mRNA levels in the diagnosis of prostate cancer among Hong Kong Chinese patients. Hong Kong Med. J. 2012, 18, 459–465. [Google Scholar] [PubMed]
- Van Neste, L.; Hendriks, R.J.; Dijkstra, S.; Trooskens, G.; Cornel, E.B.; Jannink, S.A.; de Jong, H.; Hessels, D.; Smit, F.P.; Melchers, W.J.; et al. Detection of High-grade Prostate Cancer Using a Urinary Molecular Biomarker-Based Risk Score. Eur. Opean Urol. 2016, 70, 740–748. [Google Scholar] [CrossRef]
- Chiu, P.K.; Fung, Y.H.; Teoh, J.Y.; Chan, C.H.; Lo, K.L.; Li, K.M.; Tse, R.T.; Leung, C.H.; Wong, Y.P.; Roobol, M.J.; et al. Urine spermine and multivariable Spermine Risk Score predict high-grade prostate cancer. Prostate Cancer Prostatic Dis. 2021, 24, 542–548. [Google Scholar] [CrossRef]
- Gu, X.; Wang, C.; Deng, H.; Qing, C.; Liu, R.; Liu, S.; Xue, X. Exosomal piRNA profiling revealed unique circulating piRNA signatures of cholangiocarcinoma and gallbladder carcinoma. Acta Biochim. Biophys. Sin. 2020, 52, 475–484. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, X.; Pan, C.; Qu, F.; Gan, W.; Xiang, Z.; Han, X.; Li, D. piR-31470 epigenetically suppresses the expression of glutathione S-transferase pi 1 in prostate cancer via DNA methylation. Cell Signal. 2020, 67, 109501. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, 6478. [Google Scholar] [CrossRef]
- Vasconcelos, M.H.; Caires, H.R.; Abols, A.; Xavier, C.P.R.; Line, A. Extracellular vesicles as a novel source of biomarkers in liquid biopsies for monitoring cancer progression and drug resistance. Drug Resist. Updates 2019, 47, 100647. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal miRNAs as Biomarkers for Prostate Cancer. Front. Genet. 2013, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, V.D.; Enfield, K.S.S.; Rowbotham, D.A.; Lam, W.L. An atlas of gastric PIWI-interacting RNA transcriptomes and their utility for identifying signatures of gastric cancer recurrence. Gastric Cancer 2016, 19, 660–665. [Google Scholar] [CrossRef] [Green Version]
- Fu, A.; Jacobs, D.I.; Hoffman, A.E.; Zheng, T.; Zhu, Y. PIWI-interacting RNA 021285 is involved in breast tumorigenesis possibly by remodeling the cancer epigenome. Carcinogenesis 2015, 36, 1094–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.; Song, L.; Liu, C.; Lv, X.; Li, X.; Jie, J.; Zhao, D.; Li, D. piR-55490 inhibits the growth of lung carcinoma by suppressing mTOR signaling. Tumour Biol. 2016, 37, 2749–2756. [Google Scholar] [CrossRef]
- Weng, W.; Liu, N.; Toiyama, Y.; Kusunoki, M.; Nagasaka, T.; Fujiwara, T.; Wei, Q.; Qin, H.; Lin, H.; Ma, Y.; et al. Novel evidence for a PIWI-interacting RNA (piRNA) as an oncogenic mediator of disease progression, and a potential prognostic biomarker in colorectal cancer. Mol. Cancer 2018, 17, 16. [Google Scholar] [CrossRef]
- Yan, H.; Wu, Q.L.; Sun, C.Y.; Ai, L.S.; Deng, J.; Zhang, L.; Chen, L.; Chu, Z.B.; Tang, B.; Wang, K.; et al. piRNA-823 contributes to tumorigenesis by regulating de novo DNA methylation and angiogenesis in multiple myeloma. Leukemia 2015, 29, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Hui, G.; Yuan, L.; Shi, D.; Wang, Y.; Du, M.; Zhong, D.; Ma, L.; Tong, N.; Qin, C.; et al. Identification of novel piRNAs in bladder cancer. Cancer Lett. 2015, 356, 561–567. [Google Scholar] [CrossRef]
- Yang, J.; Xue, F.T.; Li, Y.Y.; Liu, W.; Zhang, S. Exosomal piRNA sequencing reveals differences between heart failure and healthy patients. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7952–7961. [Google Scholar] [CrossRef]
- Rouget, C.; Papin, C.; Boureux, A.; Meunier, A.C.; Franco, B.; Robine, N.; Lai, E.C.; Pelisson, A.; Simonelig, M. Maternal mRNA deadenylation and decay by the piRNA pathway in the early Drosophila embryo. Nature 2010, 467, 1128–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zheng, J.; Xue, Y.; Yu, H.; Gong, W.; Wang, P.; Li, Z.; Liu, Y. PIWIL3/OIP5-AS1/miR-367-3p/CEBPA feedback loop regulates the biological behavior of glioma cells. Theranostics 2018, 8, 1084–1105. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Cheng, E.C.; Zhong, M.; Lin, H. Retrotransposons and pseudogenes regulate mRNAs and lncRNAs via the piRNA pathway in the germline. Genome Res. 2015, 25, 368–380. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef] [PubMed]
- Augello, M.A.; Den, R.B.; Knudsen, K.E. AR function in promoting metastatic prostate cancer. Cancer Metastasis Rev. 2014, 33, 399–411. [Google Scholar] [CrossRef]
- Thompson, I.M.; Ankerst, D.P.; Chi, C.; Lucia, M.S.; Goodman, P.J.; Crowley, J.J.; Parnes, H.L.; Coltman, C.A., Jr. Operating characteristics of prostate-specific antigen in men with an initial PSA level of 3.0 ng/mL or lower. JAMA 2005, 294, 66–70. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | Screening Cohort (Small RNA Sequencing) | Validation Cohort (RT-qPCR) | ||
|---|---|---|---|---|
| PCa Patients (n = 5) | Non-PCa Patients (n = 5) | PCa Patients (n = 30) | Non-PCa Patients (n = 10) | |
| Age (years) | 69(63–71) | 71(64–74) | 69(64–71) | 65(62–72) |
| PSA (ng/mL) | 7(6.6–9.9) | 1.55(1.18–2.42) | 8.7(6.88–11.55) | 2.33(1.01–3.46) |
| TRUS-PV (ml) | 36(27.3–44) | N/A | 31.5(26.5–40.3) | N/A |
| Clinical T stage | N/A | N/A | ||
| cT1 | 2(40%) | 10(33.3%) | ||
| cT2 | 2(40%) | 15(50%) | ||
| cT3 | 1(20%) | 5(16.7%) | ||
| Pathological T stage | N/A | N/A | ||
| pT2 | 1(20%) | 12(40%) | ||
| pT3 | 1(20%) | 4(13.3%) | ||
| Gleason score | N/A | N/A | ||
| 6 | 4(80%) | 15(50%) | ||
| 7–10 | 1(20%) | 15(50%) | ||
| piRNA ID | Screening Cohort (PCa = 5, Non-PCa = 5) | Validation Cohort (PCa = 30, Non-PCa = 10) | ||||
|---|---|---|---|---|---|---|
| Expression (Non-PCa) | Expression (PCa) | Log2Ratio (PCa/Non-PCa) | p adj | Up or Down- Regulate in PCa | p | |
| novel_pir99473 | 1.139 | 65.734 | 4.71 | 0 | NS | 0.0951 |
| novel_pir349843 | 0.403 | 13.49 | 3.83 | 3.4527 × 10 −155 | Upregulate | 0.0151 * |
| novel_pir382289 | 0.527 | 15.854 | 2.91 | 1.96084 × 10 −87 | Upregulate | 0.0084 ** |
| novel_pir99492 | 0.171 | 6.326 | 3.40 | 2.00497 × 10 −46 | NS | 0.5279 |
| novel_pir4470 | 0.001 | 2.283 | 7.11 | 4.94546 × 10 −22 | NS | 0.3930 |
| novel_pir158533 | 0.001 | 4.836 | 7.92 | 3.13574 × 10 −3 | Upregulate | 0.0383 * |
| novel_pir123744 | 0.001 | 2.546 | 7.00 | 9.75296 × 10 −21 | ND | ND |
| novel_pir50190 | 0.001 | 0.088 | 4.54 | 9.67 × 10 −5 | NS | 0.6335 |
| hsa_piR_001170 | 5.655 | 1.356 | −2.04 | 2.26965 × 10 −88 | NS | 0.1397 |
| hsa_piR_002468 | 2.033 | 0.076 | −4.69 | 1.4262 × 10 −49 | Upregulate | 0.0068 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Q.; Chiu, P.K.-F.; Wong, C.Y.-P.; Cheng, C.K.-L.; Teoh, J.Y.-C.; Ng, C.-F. Identification of piRNA Targets in Urinary Extracellular Vesicles for the Diagnosis of Prostate Cancer. Diagnostics 2021, 11, 1828. https://doi.org/10.3390/diagnostics11101828
Peng Q, Chiu PK-F, Wong CY-P, Cheng CK-L, Teoh JY-C, Ng C-F. Identification of piRNA Targets in Urinary Extracellular Vesicles for the Diagnosis of Prostate Cancer. Diagnostics. 2021; 11(10):1828. https://doi.org/10.3390/diagnostics11101828
Chicago/Turabian StylePeng, Qiang, Peter Ka-Fung Chiu, Christine Yim-Ping Wong, Carol Ka-Lo Cheng, Jeremy Yuen-Chun Teoh, and Chi-Fai Ng. 2021. "Identification of piRNA Targets in Urinary Extracellular Vesicles for the Diagnosis of Prostate Cancer" Diagnostics 11, no. 10: 1828. https://doi.org/10.3390/diagnostics11101828
APA StylePeng, Q., Chiu, P. K. -F., Wong, C. Y. -P., Cheng, C. K. -L., Teoh, J. Y. -C., & Ng, C. -F. (2021). Identification of piRNA Targets in Urinary Extracellular Vesicles for the Diagnosis of Prostate Cancer. Diagnostics, 11(10), 1828. https://doi.org/10.3390/diagnostics11101828