
Dual Effect of PER2 C111G Polymorphism on Cognitive Functions across Progression from Subjective Cognitive Decline to Mild Cognitive Impairment

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Clinical Assessment
2.2. Neuropsychological Assessment
2.3. APOE ε4 and PER2 C111G Genotyping
2.4. Statistical Analysis
3. Results
3.1. Description of the Sample and Comparison of Demographic Features between G Carriers and G Non-Carriers
3.2. Comparison of Neuropsychological Composite Scores between G Carriers and G Non-Carriers at Baseline
3.3. Multivariate Analysis
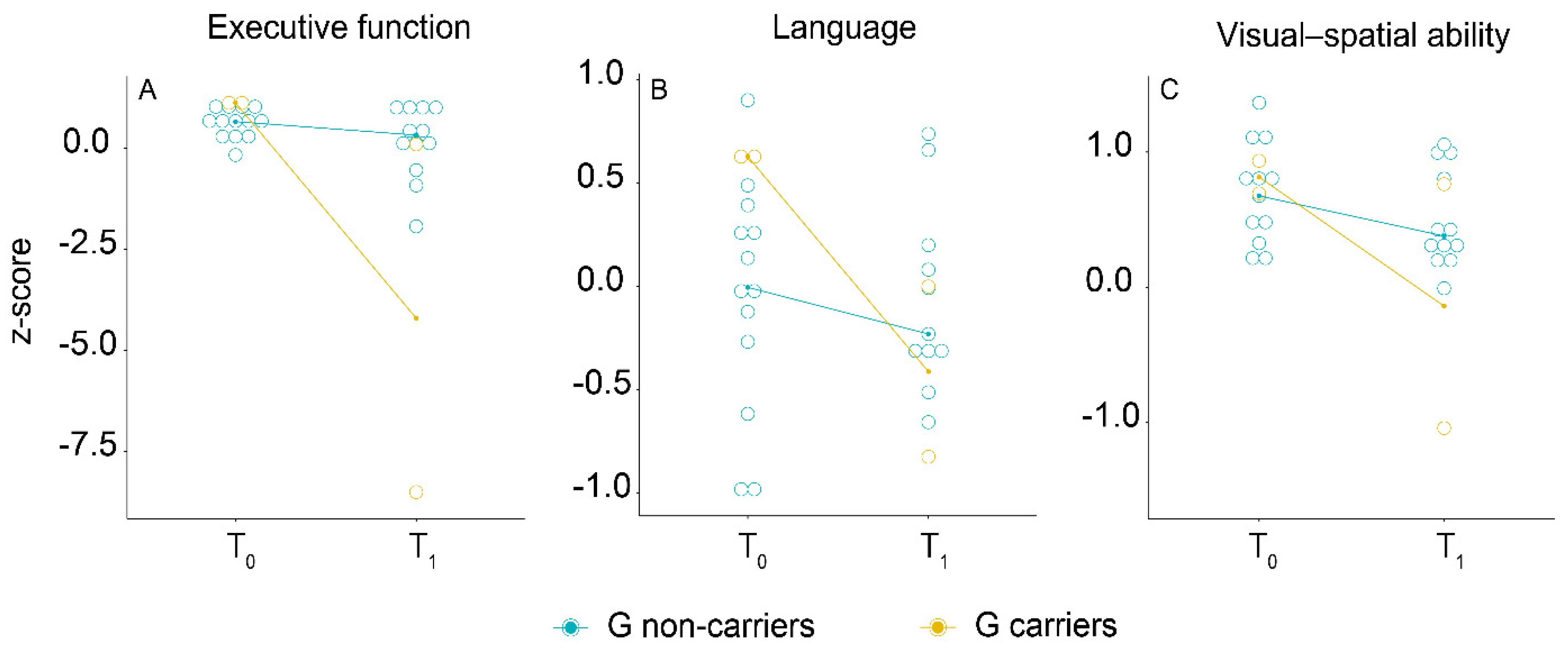
3.4. Longitudinal Association of PER2 Genotype with Cognitive Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jessen, F.; Amariglio, R.E.; van Boxtel, M.; Breteler, M.; Ceccaldi, M.; Chételat, G.; Dubois, B.; Dufouil, C.; Ellis, K.A.; van der Flier, W.M.; et al. A Conceptual Framework for Research on Subjective Cognitive Decline in Preclinical Alzheimer’s Disease. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2014, 10, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Perrotin, A.; Mormino, E.C.; Madison, C.M.; Hayenga, A.O.; Jagust, W.J. Subjective Cognition and Amyloid Deposition Imaging: A Pittsburgh Compound B Positron Emission Tomography Study in Normal Elderly Individuals. Arch. Neurol. 2012, 69, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amariglio, R.E.; Becker, J.A.; Carmasin, J.; Wadsworth, L.P.; Lorius, N.; Sullivan, C.; Maye, J.E.; Gidicsin, C.; Pepin, L.C.; Sperling, R.A.; et al. Subjective Cognitive Complaints and Amyloid Burden in Cognitively Normal Older Individuals. Neuropsychologia 2012, 50, 2880–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, R.; Godin, O.; Crivello, F.; Maillard, P.; Mazoyer, B.; Tzourio, C.; Dufouil, C. Longitudinal Neuroimaging Correlates of Subjective Memory Impairment: 4-Year Prospective Community Study. Br. J. Psychiatry 2011, 198, 199–205. [Google Scholar] [CrossRef]
- Mitchell, A.J.; Beaumont, H.; Ferguson, D.; Yadegarfar, M.; Stubbs, B. Risk of Dementia and Mild Cognitive Impairment in Older People with Subjective Memory Complaints: Meta-Analysis. Acta Psychiatr. Scand. 2014, 130, 439–451. [Google Scholar] [CrossRef]
- Parfenov, V.A.; Zakharov, V.V.; Kabaeva, A.R.; Vakhnina, N.V. Subjective Cognitive Decline as a Predictor of Future Cognitive Decline: A Systematic Review. Dement. Neuropsychol. 2020, 14, 248–257. [Google Scholar] [CrossRef]
- Mazzeo, S.; Padiglioni, S.; Bagnoli, S.; Bracco, L.; Nacmias, B.; Sorbi, S.; Bessi, V. The Dual Role of Cognitive Reserve in Subjective Cognitive Decline and Mild Cognitive Impairment: A 7-Year Follow-up Study. J. Neurol. 2019, 266, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, S.; Bessi, V.; Padiglioni, S.; Bagnoli, S.; Bracco, L.; Sorbi, S.; Nacmias, B. KIBRA T Allele Influences Memory Performance and Progression of Cognitive Decline: A 7-Year Follow-up Study in Subjective Cognitive Decline and Mild Cognitive Impairment. Neurol. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Bessi, V.; Mazzeo, S.; Padiglioni, S.; Piccini, C.; Nacmias, B.; Sorbi, S.; Bracco, L. From Subjective Cognitive Decline to Alzheimer’s Disease: The Predictive Role of Neuropsychological Assessment, Personality Traits, and Cognitive Reserve. A 7-Year Follow-Up Study. J. Alzheimer’s Dis. 2018, 63, 1523–1535. [Google Scholar] [CrossRef]
- Bessi, V.; Giacomucci, G.; Mazzeo, S.; Bagnoli, S.; Padiglioni, S.; Balestrini, J.; Tomaiuolo, G.; Piaceri, I.; Carraro, M.; Bracco, L.; et al. PER2 C111G Polymorphism, Cognitive Reserve and Cognition in Subjective Cognitive Decline and Mild Cognitive Impairment. A 10-Year Follow-up Study. Eur. J. Neurol. 2020. [Google Scholar] [CrossRef]
- Wang, L.M.-C.; Dragich, J.M.; Kudo, T.; Odom, I.H.; Welsh, D.K.; O’Dell, T.J.; Colwell, C.S. Expression of the Circadian Clock Gene Period2 in the Hippocampus: Possible Implications for Synaptic Plasticity and Learned Behaviour. ASN Neuro 2009, 1. [Google Scholar] [CrossRef] [Green Version]
- Lawton, M.P.; Brody, E.M. Assessment of Older People: Self-Maintaining and Instrumental Activities of Daily Living. Gerontologist 1969, 9, 179–186. [Google Scholar] [CrossRef]
- Albert, M.S.; DeKosky, S.T.; Dickson, D.; Dubois, B.; Feldman, H.H.; Fox, N.C.; Gamst, A.; Holtzman, D.M.; Jagust, W.J.; Petersen, R.C.; et al. The Diagnosis of Mild Cognitive Impairment Due to Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Alzheimer’s Dement. 2011, 7, 270–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The Diagnosis of Dementia Due to Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2011, 7, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Neary, D.; Snowden, J.S.; Gustafson, L.; Passant, U.; Stuss, D.; Black, S.; Freedman, M.; Kertesz, A.; Robert, P.H.; Albert, M.; et al. Frontotemporal Lobar Degeneration: A Consensus on Clinical Diagnostic Criteria. Neurology 1998, 51, 1546–1554. [Google Scholar] [CrossRef] [Green Version]
- Román, G.C.; Tatemichi, T.K.; Erkinjuntti, T.; Cummings, J.L.; Masdeu, J.C.; Garcia, J.H.; Amaducci, L.; Orgogozo, J.M.; Brun, A.; Hofman, A. Vascular Dementia: Diagnostic Criteria for Research Studies. Report of the NINDS-AIREN International Workshop. Neurology 1993, 43, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Bracco, L.; Amaducci, L.; Pedone, D.; Bino, G.; Lazzaro, M.P.; Carella, F.; D’Antona, R.; Gallato, R.; Denes, G. Italian Multicentre Study on Dementia (SMID): A Neuropsychological Test Battery for Assessing Alzheimer’s Disease. J. Psychiatr. Res. 1990, 24, 213–226. [Google Scholar] [CrossRef]
- Crook, T.H.; Feher, E.P.; Larrabee, G.J. Assessment of Memory Complaint in Age-Associated Memory Impairment: The MAC-Q. Int. Psychogeriatr. 1992, 4, 165–176. [Google Scholar] [CrossRef]
- Caffarra, P.; Vezzadini, G.; Dieci, F.; Zonato, F.; Venneri, A. Rey-Osterrieth Complex Figure: Normative Values in an Italian Population Sample. Neurol. Sci. 2002, 22, 443–447. [Google Scholar] [CrossRef]
- Baddeley, A.; Della Sala, S.; Papagno, C.; Spinnler, H. Dual-Task Performance in Dysexecutive and Nondysexecutive Patients with a Frontal Lesion. Neuropsychology 1997, 11, 187–194. [Google Scholar] [CrossRef]
- Spinnler, H.; Tognoni, G. Standardizzazione e Taratura Italiana Di Test Neuropsicologici: Gruppo Italiano per Lo Studio Neuropsicologico Dell’invecchiamento; Masson Italia Periodici: Milano, Italy, 1987. [Google Scholar]
- Giovagnoli, A.R.; Del Pesce, M.; Mascheroni, S.; Simoncelli, M.; Laiacona, M.; Capitani, E. Trail Making Test: Normative Values from 287 Normal Adult Controls. Ital. J. Neurol. Sci. 1996, 17, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Brazzelli, M.; Della Sala, S.; Laiacona, M. Calibration of the Italian Version of the Rivermead Behavioural Memory Test: A Test for the Ecological Evaluation of Memory. Boll. Psicol. Appl. 1993, 206, 33–42. [Google Scholar]
- Bozon, B.; Kelly, A.; Josselyn, S.A.; Silva, A.J.; Davis, S.; Laroche, S. MAPK, CREB and Zif268 Are All Required for the Consolidation of Recognition Memory. Philos. Trans. R. Soc. B 2003, 358, 805–814. [Google Scholar] [CrossRef] [Green Version]
- Lonze, B.E.; Ginty, D.D. Function and Regulation of CREB Family Transcription Factors in the Nervous System. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Arjona, A.; Sarkar, D.K. The Circadian Gene MPer2 Regulates the Daily Rhythm of IFN-Gamma. J. Interferon Cytokine Res. 2006, 26, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Sujino, M.; Nagano, M.; Fujioka, A.; Shigeyoshi, Y.; Inouye, S.-I.T. Temporal Profile of Circadian Clock Gene Expression in a Transplanted Suprachiasmatic Nucleus and Peripheral Tissues. Eur. J. Neurosci. 2007, 26, 2731–2738. [Google Scholar] [CrossRef]
- Albrecht, U.; Sun, Z.S.; Eichele, G.; Lee, C.C. A Differential Response of Two Putative Mammalian Circadian Regulators, Mper1 and Mper2, to Light. Cell 1997, 91, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Hood, S.; Cassidy, P.; Cossette, M.-P.; Weigl, Y.; Verwey, M.; Robinson, B.; Stewart, J.; Amir, S. Endogenous Dopamine Regulates the Rhythm of Expression of the Clock Protein PER2 in the Rat Dorsal Striatum via Daily Activation of D2 Dopamine Receptors. J. Neurosci. 2010, 30, 14046–14058. [Google Scholar] [CrossRef] [PubMed]
- Gravotta, L.; Gavrila, A.M.; Hood, S.; Amir, S. Global Depletion of Dopamine Using Intracerebroventricular 6-Hydroxydopamine Injection Disrupts Normal Circadian Wheel-Running Patterns and PERIOD2 Expression in the Rat Forebrain. J. Mol. Neurosci. 2011, 45, 162–171. [Google Scholar] [CrossRef]
- Spanagel, R.; Pendyala, G.; Abarca, C.; Zghoul, T.; Sanchis-Segura, C.; Magnone, M.C.; Lascorz, J.; Depner, M.; Holzberg, D.; Soyka, M.; et al. The Clock Gene Per2 Influences the Glutamatergic System and Modulates Alcohol Consumption. Nat. Med. 2005, 11, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Yelamanchili, S.V.; Pendyala, G.; Brunk, I.; Darna, M.; Albrecht, U.; Ahnert-Hilger, G. Differential Sorting of the Vesicular Glutamate Transporter 1 into a Defined Vesicular Pool Is Regulated by Light Signaling Involving the Clock Gene Period2. J. Biol. Chem. 2006, 281, 15671–15679. [Google Scholar] [CrossRef] [Green Version]
- Straub, R.H.; Cutolo, M. Circadian Rhythms in Rheumatoid Arthritis: Implications for Pathophysiology and Therapeutic Management. Arthritis Rheum. 2007, 56, 399–408. [Google Scholar] [CrossRef]
- Mendoza, J.; Clesse, D.; Pévet, P.; Challet, E. Serotonergic Potentiation of Dark Pulse-Induced Phase-Shifting Effects at Midday in Hamsters. J. Neurochem. 2008, 106, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Caldelas, I.; Challet, E.; Saboureau, M.; Pevet, P. Light and Melatonin Inhibit in Vivo Serotonergic Phase Advances without Altering Serotonergic-Induced Decrease of per Expression in the Hamster Suprachiasmatic Nucleus. J. Mol. Neurosci. 2005, 25, 53–63. [Google Scholar] [CrossRef]
- Yesavage, J.A.; Noda, A.; Hernandez, B.; Friedman, L.; Cheng, J.J.; Tinklenberg, J.R.; Hallmayer, J.; O’hara, R.; David, R.; Robert, P.; et al. Circadian Clock Gene Polymorphisms and Sleep-Wake Disturbance in Alzheimer Disease. Am. J. Geriatr. Psychiatry 2011, 19, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, P.A.; Alvim-Soares, A.; Bicalho, M.A.C.; de Moraes, E.N.; Malloy-Diniz, L.; de Paula, J.J.; Romano-Silva, M.A.; Miranda, D.M. Lack of Association between Genetic Polymorphism of Circadian Genes (PER2, PER3, CLOCK and OX2R) with Late Onset Depression and Alzheimer’s Disease in a Sample of a Brazilian Population (Circadian Genes, Late-Onset Depression and Alzheimer’s Disease). Curr. Alzheimer Res. 2016, 13, 1397–1406. [Google Scholar] [CrossRef]
- Cermakian, N.; Lamont, E.W.; Boudreau, P.; Boivin, D.B. Circadian Clock Gene Expression in Brain Regions of Alzheimer ’s Disease Patients and Control Subjects. J. Biol. Rhythm. 2011, 26, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Bessi, V.; Mazzeo, S.; Bagnoli, S.; Padiglioni, S.; Carraro, M.; Piaceri, I.; Bracco, L.; Sorbi, S.; Nacmias, B. The Implication of BDNF Val66Met Polymorphism in Progression from Subjective Cognitive Decline to Mild Cognitive Impairment and Alzheimer’s Disease: A 9-Year Follow-up Study. Eur. Arch. Psychiatry Clin. Neurosci. 2020, 270, 471–482. [Google Scholar] [CrossRef]
- Stern, Y. Cognitive Reserve in Ageing and Alzheimer’s Disease. Lancet Neurol. 2012, 11, 1006–1012. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Variable | G Non-Carriers | G Carriers | p |
|---|---|---|---|
| N | 36 | 9 | |
| Age at baseline, median (IQR) | 60.80 (17.24) | 62.44 (9.81) | 0.758 |
| Disease duration, median (IQR) | 3.82 (2.92) | 3.33 (4.24) | 0.092 |
| Sex (no. women/no. men) | 25/11 | 8/1 | 0.241 |
| Education, median (IQR) | 12.00 (9.00) | 8.00 (7.00) | 0.016 |
| APOE ε4+, % (95% CI) | 30.55 (16.78–44.32) | 22.22 (9.79–34.65) | 0.625 |
| MMSE, median (IQR) | 27.00 (3.70) | 26.61 (4.70) | 0.251 |
| HDRS, median (IQR) | 4.00 (6.00) | 5.50 (4.00) | 0.303 |
| MAC-Q, median (IQR) | 25.00 (3.00) | 25.50 (4.00) | 0.326 |
| Long-term verbal memory, median (IQR) | 0.42 (0.54) | 0.25 (0.62) | 0.244 |
| Working memory, median (IQR) | −1.25 (1.61) | −1.03 (1.61) | 0.770 |
| Visual–spatial memory, median (IQR) | 0.36 (0.83) | 0.00 (0.18) | 0.163 |
| Visual–spatial ability, median (IQR) | 0.40 (±0.61) | 0.49 (±0.47) | 0.694 |
| Executive function, median (IQR) | 0.65 (±0.40) | 0.97 (±0.72) | 0.305 |
| Language, median (IQR) | 0.25 (0.94) | 0.62 (0.52) | 0.018 |
| B | Wald | p | OR | 95% C.I. | ||
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| 3.32 | 4.92 | 0.027 | 25.79 | 1.44 | 462.50 | |
| Education (years) | −0.345 | 4.05 | 0.044 | 0.71 | 0.50 | 0.99 |
| Age (years) | −0.05 | 0.48 | 0.49 | 0.95 | 0.82 | 1.10 |
| Disease duration (years) | −0.30 | 2.16 | 0.14 | 0.74 | 0.49 | 1.11 |
| Female sex | 0.35 | 0.06 | 0.80 | 1.41 | 0.10 | 20.22 |
| APOE ε4+ | 0.43 | 0.13 | 0.72 | 1.53 | 0.15 | 16.06 |
| Constant | 4.82 | 0.81 | 0.40 | 124.46 | ||
| Variable | G Non-Carriers | G Carriers | p |
|---|---|---|---|
| N | 36 | 9 | |
| Long-term verbal memory, median (IQR) | 0.07 (0.94) | 0.13 (0.87) | 0.547 |
| Working memory, median (IQR) | −1.25 (1.03) | −1.25 (0.79) | 0.833 |
| Visual-spatial memory, median (IQR) | 0.82 (1.19) | 0.22 (2.08) | 0.276 |
| Visual-spatial ability, median (IQR) | 0.50 (0.71) | 0.76 (0.99) | 0.323 |
| Executive function, median (IQR) | 0.51 (0.71) | 0.37 (1.91) | 0.838 |
| Language, median (IQR) | 0.20 (0.86) | 0.18 (0.46) | 0.850 |
| Follow-Up Time | Follow-Up Time × G Allele | |
|---|---|---|
| β (95% CI) | β (95% CI) | |
| Long-term verbal memory | 0.942 (0.889:0.999) | 1.016 (0.918:1.125) |
| Working memory | 1.013 (0.973:1.054) | 0.971 (0.922:1.023) |
| Visual–spatial memory | 1.034 (0.986:1.085) | 0.987 (0.856:1.138) |
| Visual–spatial ability | 1.030 (0.993:1.068) | 0.973 (0.874:1.083) |
| Executive function | 0.974 (0.944:1.006) | 0.335 (0.595:1.193) |
| Language | 0.995 (0.975:1.015) | 0.971 (0.923:1.021) |
| Cognitive Domains | Time | Time × G Allele |
|---|---|---|
| β (95% C.I) | β (95% C.I) | |
| Long-term verbal memory | 0.823 (0.785:0.862) *** | 1.049 (0.995:1.106) |
| Working memory | 1.017 (0.972:1.063) | 0.449 (0.108:1.866) |
| Visual–spatial memory | 0.987 (0.938:1.038) | 0.980 (0.788:1.006) |
| Visual–spatial ability | 0.977 (0.954:1.047) | 0.885 (0.827:0.946) *** |
| Executive function | 0.979 (0.954:1.005) | 0.921 (0.897:0.946) *** |
| Language | 0.941 (0.885:1.001) | 0.560 (0.475:0.660) *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzeo, S.; Bessi, V.; Bagnoli, S.; Giacomucci, G.; Balestrini, J.; Padiglioni, S.; Tomaiuolo, G.; Ingannato, A.; Ferrari, C.; Bracco, L.; et al. Dual Effect of PER2 C111G Polymorphism on Cognitive Functions across Progression from Subjective Cognitive Decline to Mild Cognitive Impairment. Diagnostics 2021, 11, 718. https://doi.org/10.3390/diagnostics11040718
Mazzeo S, Bessi V, Bagnoli S, Giacomucci G, Balestrini J, Padiglioni S, Tomaiuolo G, Ingannato A, Ferrari C, Bracco L, et al. Dual Effect of PER2 C111G Polymorphism on Cognitive Functions across Progression from Subjective Cognitive Decline to Mild Cognitive Impairment. Diagnostics. 2021; 11(4):718. https://doi.org/10.3390/diagnostics11040718
Chicago/Turabian StyleMazzeo, Salvatore, Valentina Bessi, Silvia Bagnoli, Giulia Giacomucci, Juri Balestrini, Sonia Padiglioni, Giulia Tomaiuolo, Assunta Ingannato, Camilla Ferrari, Laura Bracco, and et al. 2021. "Dual Effect of PER2 C111G Polymorphism on Cognitive Functions across Progression from Subjective Cognitive Decline to Mild Cognitive Impairment" Diagnostics 11, no. 4: 718. https://doi.org/10.3390/diagnostics11040718
APA StyleMazzeo, S., Bessi, V., Bagnoli, S., Giacomucci, G., Balestrini, J., Padiglioni, S., Tomaiuolo, G., Ingannato, A., Ferrari, C., Bracco, L., Sorbi, S., & Nacmias, B. (2021). Dual Effect of PER2 C111G Polymorphism on Cognitive Functions across Progression from Subjective Cognitive Decline to Mild Cognitive Impairment. Diagnostics, 11(4), 718. https://doi.org/10.3390/diagnostics11040718