
Influence of Cerebral Vasodilation on Blood Reelin Levels in Growth Restricted Fetuses

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Patients and Methods
2.1. Selection of Patients and Evaluation of Fetal Growth
2.2. Sample Collection, Initial Processing, and Storage
2.3. Determination of Reelin in the Umbilical Vein through Western Blotting
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, C.C.; Santolaya-Forgas, J. Current concepts of fetal growth restriction: Part I. Causes, classification, and pathophysiology. Obstet. Gynecol. 1998, 92, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, C.; Marino, C.; Nosarti, C.; Vieno, A.; Visentin, S.; Simonelli, A. Association of Intrauterine Growth Restriction and Small for Gestational Age Status With Childhood Cognitive Outcomes. JAMA Pediatr. 2020, 174, 772. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, T.; Thilaganathan, B.; Hooper, R.; Khan, K.S.; Bhide, A. Neurodevelopmental delay in small babies at term: A systematic review. Ultrasound Obstet. Gynecol. 2012, 40, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Baschat, A.A. Neurodevelopment after Fetal Growth Restriction. Fetal Diagn. Ther. 2014, 36, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P. Adult consequences of fetal growth restriction. Clin. Obstet. Gynecol. 2006, 49, 270–283. [Google Scholar] [CrossRef]
- Rees, S.; Harding, R.; Walker, D. The biological basis of injury and neuroprotection in the fetal and neonatal brain. Int. J. Dev. Neurosci. 2011, 29, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Inversetti, A.; Fesslova, V.; Deprest, J.; Candiani, M.; Giorgione, V.; Cavoretto, P. Prenatal Growth in Fetuses with Isolated Cyanotic and Non-Cyanotic Congenital Heart Defects. Fetal Diagn. Ther. 2020, 47, 411–419, Epub 2018 Nov 9. [Google Scholar] [CrossRef] [PubMed]
- Golan, H.; Huleihel, M. The effect of prenatal hypoxia on brain development: Short- and long-term consequences demonstrated in rodent models. Dev. Sci. 2006, 9, 338–349. [Google Scholar] [CrossRef]
- Pugash, D.; Hendson, G.; Dunham, C.P.; Dewar, K.; Money, D.M.; Prayer, D. Sonographic assessment of normal and abnormal patterns of fetal cerebral lamination. Ultrasound Obstet. Gynecol. 2012, 40, 642–651. [Google Scholar] [CrossRef]
- Eixarch, E.; Muñoz-Moreno, E.; Bargalló, N.; Batalle, D.; Gratacos, E. Motor and cortico-striatal-thalamic connectivity alterations in intrauterine growth restriction. Am. J. Obstet. Gynecol. 2016, 214, 725.e1–725.e9. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Andrade, E.; Figueroa-Diesel, H.; Jansson, T.; Rangel-Nava, H.; Gratacos, E. Changes in regional fetal cerebral blood flow perfusion in relation to hemodynamic deterioration in severely growth-restricted fetuses. Ultrasound Obstet. Gynecol. 2008, 32, 71–76. [Google Scholar] [CrossRef]
- Cetin, I.; Barberis, B.; Brusati, V.; Brighina, E.; Mandia, L.; Arighi, A.; Rango, M. Lactate detection in the brain of growth-restricted fetuses with magnetic resonance spectroscopy. Am. J. Obstet. Gynecol. 2011, 205, 350.e1–350.e7. [Google Scholar] [CrossRef]
- Story, L.; Damodaram, M.S.; Allsop, J.M.; McGuinness, A.; Patel, A.; Wylezinska, M.; Rutherford, M.A. Brain metabolism in fetal intrauterine growth restriction: A proton magnetic resonance spectroscopy study. Am. J. Obstet. Gynecol. 2011, 205, 483.e1–483.e8. [Google Scholar] [CrossRef]
- Briana, D.D.; Malamitsi-Puchner, A. Perinatal biomarkers implying “Developmental Origins of Health and Disease” consequences in intrauterine growth restriction. Acta Paediatr. 2019, 109, 1317–1322. [Google Scholar] [CrossRef]
- Golan, M.H.; Mane, R.; Molczadzki, G.; Zuckerman, M.; Kaplan-Louson, V.; Huleihel, M.; Perez-Polo, J.R. Impaired migration signaling in the hippocampus following prenatal hypoxia. Neuropharmacology 2009, 57, 511–522. [Google Scholar] [CrossRef]
- Frotscher, M. Role for Reelin in stabilizing cortical architecture. Trends Neurosci. 2010, 33, 407–414. [Google Scholar] [CrossRef]
- Fatemi, S.H. Reeline Glycoprotein: Structura, Biology and Roles in Health and Disease, 2008th ed.; Springer: Verlag, NY, USA, 2008. [Google Scholar]
- Suárez-Solá, M.L.; González-Delgado, F.J.; Pueyo-Morlans, M.; Medina-Bolívar, O.C.; Hernández-Acosta, N.C.; González-Gómez, M.; Meyer, G. Neurons in the white matter of the adult human neocortex. Front Neuroanat. 2009, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Curristin, S.M.; Cao, A.; Stewart, W.B.; Zhang, H.; Madri, J.A.; Morrow, J.S.; Ment, L.R. Disrupted synaptic development in the hypoxic newborn brain. Proc. Natl. Acad. Sci. USA 2002, 99, 15729–15734. [Google Scholar] [CrossRef] [Green Version]
- Nisimov, H.; Orenbuch, A.; Pleasure, S.J.; Golan, H.M. Impaired Organization of GABAergic Neurons Following Prenatal Hypoxia. Neuroscience. 2018, 384, 300–313. [Google Scholar] [CrossRef]
- Ghiani, C.A.; Mattan, N.S.; Nobuta, H.; Malvar, J.S.; Boles, J.; Ross, M.G.; de Vellis, J. Early effects of lipopolysaccharide-induced inflammation on foetal brain development in rat. ASN Neuro 2011, 3, 233–245. [Google Scholar] [CrossRef]
- Rojas, J. Corticogénesis y neurodegeneración: Implicaciones de la vía de la reelina en la patogenia de la enfermedad de Alzheimer. Arch. Neurocienc. 2009, 14, 132–141. [Google Scholar]
- Jossin, Y. Reelin Functions, Mechanisms of Action and Signaling Pathways during Brain Development and Maturation. Biomolecules 2020, 10, 964. [Google Scholar] [CrossRef]
- Lees, C.C.; Stampalija, T.; Baschat, A.; Da Silva Costa, F.; Ferrazzi, E.; Figueras, F.; Unterscheider, J. ISUOG Practice Guidelines: Diagnosis and management of small-for-gestational-age fetus and fetal growth restriction. Ultrasound Obstet. Gynecol. 2020, 56, 298–312. [Google Scholar] [CrossRef]
- Ciobanu, A.; Wright, A.; Syngelaki, A.; Wright, D.; Akolekar, R.; Nicolaides, K.H. Fetal Medicine Foundation reference ranges for umbilical artery and middle cerebral artery pulsatility index and cerebroplacental ratio. Ultrasound Obstet. Gynecol. 2019, 53, 465–472, Epub 2019 Feb 13. [Google Scholar] [CrossRef]
- Gómez, O.; Figueras, F.; Fernández, S.; Bennasar, M.; Martínez, J.M.; Puerto, B.; Gratacós, E. Reference ranges for uterine artery mean pulsatility index at 11-41 weeks of gestation. Ultrasound Obstet. Gynecol. 2008, 32, 128–132. [Google Scholar] [CrossRef]
- Lugli, G.; Krueger, J.M.; Davis, J.M.; Persico, A.M.; Keller, F.; Smalheiser, N.R. Methodological factors influencing measurement and processing of plasma reelin in humans. BMC Biochem. 2003, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Moreno, T.; Galazo, M.J.; Porrero, C.; Martínez-Cerdeño, V.; Clascá, F. Extracellular matrix molecules and synaptic plasticity: Immunomapping of intracellular and secreted Reelin in the adult rat brain. Eur. J. Neurosci. 2006, 23, 401–422. [Google Scholar] [CrossRef]
- Alvarez-Dolado, M.; Ruiz, M.; del Río, J.A.; Alcántara, S.; Burgaya, F.; Sheldon, M.; Muñoz, A. Thyroid hormone regulates reelin and dab1 expression during brain development. J. Neurosci. 1999, 19, 6979–6993. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhai, X.; Liu, Y.; Li, J.; Shan, Z.; Teng, W. Treatment with Iodine in Pregnant Rats with Marginal Iodine Deficiency Improves Cell Migration in the Developing Brain of the Progeny. Mol. Neurobiol. 2015, 53, 2212–2221. [Google Scholar] [CrossRef]
- Skorput, A.G.J.; Yeh, H.H. Effects of ethanol exposure in utero on Cajal-Retzius cells in the developing cortex. Alcohol Clin. Exp. Res. 2015, 39, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Rideau Batista Novais, A.; Guiramand, J.; Cohen-Solal, C.; Crouzin, N.; de Jesus Ferreira, M.C.; Vignes, M.; Cambonie, G. N-acetyl-cysteine prevents pyramidal cell disarray and reelin-immunoreactive neuron deficiency in CA3 after prenatal immune challenge in rats. Pediatric Res. 2013, 73, 750–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatemi, S.H.; Emamian, E.S.; Kist, D.; Sidwell, R.W.; Nakajima, K.; Akhter, P.; Bailey, K. Defective corticogenesis and reduction in Reelin immunoreactivity in cortex and hippocampus of prenatally infected neonatal mice. Mol. Psychiatry 1999, 4, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brummelte, S.; Galea, L.A.M.; Devlin, A.M.; Oberlander, T.F. Antidepressant use during pregnancy and serotonin transporter genotype (SLC6A4) affect newborn serum reelin levels. Dev. Psychobiol. 2013, 55, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khialeeva, E.; Carpenter, E.M. Nonneuronal roles for the reelin signaling pathway. Dev. Dyn. 2016, 246, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Costas, E.; Fenton, E.Y.; Caruncho, H.J. Reelin expression in brain endothelial cells: An electron microscopy study. BMC Neurosci. 2015, 16, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ek, C.J.; Angelo, B.D.A.; Baburamani, A.A.; Lehner, C.; Leverin, A.-L.; Smith, P.L.; Mallard, C. Brain barrier properties and cerebral blood. J. Cereb. Blood Flow Metab. 2015, 35, 818–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatemi, S.H.; Kroll, J.L.; Stary, J.M. Altered levels of Reelin and its isoforms in schizophrenia and mood disorders. Neuroreport 2001, 12, 3209–3215. [Google Scholar] [CrossRef]
- Hornig, T.; Sturm, L.; Fiebich, B.; Tebartz van Elst, L. Increased Blood-Reelin-Levels in First Episode Schizophrenia. PLoS ONE 2015, 10, e0134671. [Google Scholar] [CrossRef]
- Botella-López, A.; Burgaya, F.; Gavín, R.; García-Ayllón, M.S.; Gómez-Tortosa, E.; Peña-Casanova, J.; Sáez-Valero, J. Reelin expression and glycosylation patterns are altered in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2006, 103, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Gonzalez, P.; Zhang, L. Fetal stress and programming of hypoxic/ischemic-sensitive phenotype in the neonatal brain: Mechanisms and possible interventions. Prog. Neurobiol. 2012, 98, 145–165. [Google Scholar] [CrossRef] [Green Version]
- Komitova, M.; Xenos, D.; Salmaso, N.; May, T.K.; Brand, T.; Schwartz, M.L.; Vaccarino, F.M. Hypoxia-Induced Developmental Delays of Inhibitory Interneurons Are Reversed by Environmental Enrichment in the Postnatal Mouse Forebrain. J. Neurosci. 2013, 33, 13375–13387. [Google Scholar] [CrossRef]
- Courtès, S.; Vernerey, J.; Pujadas, L.; Magalon, K.; Cremer, H.; Soriano, E.; Cayre, M. Reelin controls progenitor cell migration in the healthy and pathological adult mouse brain. PLoS ONE 2011, 6, e20430. [Google Scholar] [CrossRef] [Green Version]
- Ohishi, T.; Wang, L.; Ogawa, B.; Fujisawa, K.; Taniai, E.; Hayashi, H.; Shibutani, M. No effect of sustained systemic growth retardation on the distribution of Reelin-expressing interneurons in the neuron-producing hippocampal dentate gyrus in rats. Reprod. Toxicol. 2010, 30, 591–599. [Google Scholar] [CrossRef]
- Lane-Donovan, C.; Desai, C.; Pohlkamp, T.; Plautz, E.J.; Herz, J.; Stowe, A.M. Physiologic Reelin does not play a strong role in protection against acute stroke. J. Cereb. Blood Flow Metab. 2016, 36, 1295–1303. [Google Scholar] [CrossRef] [Green Version]
- Baschat, A.A.; Gembruch, U.; Harman, C.R. The sequence of changes in Doppler and biophysical parameters as severe fetal growth restriction worsens. Ultrasound Obstet. Gynecol. 2001, 18, 571–577. [Google Scholar] [CrossRef]
- Saunders, N.R.; Liddelow, S.A.; Dziegielewska, K.M. Barrier mechanisms in the developing brain. Front. Pharmacol. 2012, 3, 46. [Google Scholar] [CrossRef] [Green Version]
- Meher, S.; Hernandez-Andrade, E.; Basheer, S.N.; Lees, C. Impact of cerebral redistribution on neurodevelopmental outcome in small for gestational age or growth restricted babies: A systematic review. Ultrasound Obstet. Gynecol. 2015, 46, 398–404. [Google Scholar] [CrossRef]
- Sanz-Cortés, M.; Carbajo, R.J.; Crispi, F.; Figueras, F.; Pineda-Lucena, A.; Gratacos, E. Metabolomic Profile of Umbilical Cord Blood Plasma from Early and Late Intrauterine Growth Restricted (IUGR) Neonates with and without Signs of Brain Vasodilation. PLoS ONE 2013, 8, e80121. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GROUP 1 (Control) AGA Fetuses N:17 | GROUP 2 FGR Fetuses (Pathological MCA Normal UA) N:9 | GROUP 3 FGR Fetuses (Pathological UA) N:8 | p | |
|---|---|---|---|---|
| Maternal Age (years) m(SD) | 31.35 (4,5) | 31.38 (5.7) | 30.75 (5.6) | 0.91 |
| Gestational Age (weeks) m(SD) | 38.39 (2.2) | 36.04 (2.4) | 32.38 (1.8) | <0.001 |
| Fetal sex (female) n(%) | 7 (41) | 5 (56) | 2 (25) | 0.433 |
| Type of delivery (vaginal) n(%) | 10 (59) | 2 (22) | 0 | 0.003 |
| Birth weight (gr) M(IQR) | 3316 (350) | 2031 (698) | 1446 (620) | <0.001 |
| Percentile at birth M(IQR) | 63 (38) | 2 (5) | 2 (1) | <0.001 |
| pH UA (mean) M(IQR) | 7.26 (0.17) | 7.26 (0.13) | 7.27 (0.13) | 0.530 |
| Uterine artery PI > p95 n(%) | - | 3 (33) | 6 (67) | <0.001 |
| Rupture of membranes > than 12 h. n(%) | 4 (24) | 0 | 0 | 0.133 |
| Admission to the NICU n(%) | 1 (12) | 3 (33) | 6 (75) | 0.001 |
| Type 1 intraventricular hemorrhage n(%) | 0 | 0 | 2 (25) | 0.005 |
| Head circumference (cm) m(SD) | 34.4 (1) | 31.5 (3) | 28.13 (3) | <0.001 |
| Leukocytes (n º/uL) m(SD) | 14,725 (10,200) | 15,212 (7625) | 11,426 (2800) | 0.308 |
| Cortisol M(IQR) | 8.55 (5) | 19.70 (14.7) | 1.96 (2.7) | 0.035 |
| Reelin M(IQR) | 15,055 (3954) | 20,062(16,645) | 10,111(6956) | 0.083 |
| Total Reelin M (SD) | p | |
|---|---|---|
| Maternal Obtetric Characteristics | ||
| Age (years) | −251 (−928 426) | 0.456 |
| First pregnancy No (11) | 0.189 | |
| 13,318 (6459) | ||
| Yes (23) | 17,868(10,273) | |
| Disease during pregnancy (preeclampsia, diabetes, Hypertensión, hypothiroidism) No(26) | 0.254 | |
| 17,394 (9308) | ||
| Yes (8) | 10,968 (6167) | |
| Rupture of membranes > than 12 h. No (28) | 0.58 | |
| 17,102 (10,043) | ||
| Yes (4) | 14,147 (6311) | |
| SO4Mg treatment No (29) | 0.94 | |
| 16,147(8946) | ||
| Yes (3) | 16,619(17,911) | |
| Type of delivery Vaginal (19) | 17,288 (9077) | 0.43 |
| C-Section (15) | 14,532 (10,117) | |
| Neonatal Characteristics | ||
| Gestational age (34) | 0.18 | 0.309 |
| Gender Male (20) | 0.388 | |
| 15,111(7554) | ||
| Female (14) | 17,919(11,491) | |
| Birthweight (g) (34) | -0.02 | 0.911 |
| Lung maturation Yes (8) | 0.728 | |
| 15,645 | ||
| Arterial cord pH (34) | 0.111 | 0.531 |
| Neonatal reanimation (32) 0 (17) | 0.661 | |
| 16,846(10,625) | ||
| I (8) | 16,606(6002) | |
| II (3) | 9592(7782) | |
| III (5) | 16,639(11,965) | |
| NICU No (24) | 0.592 | |
| 16,776(9261) | ||
| Yes (10) | 14,880(9661) | |
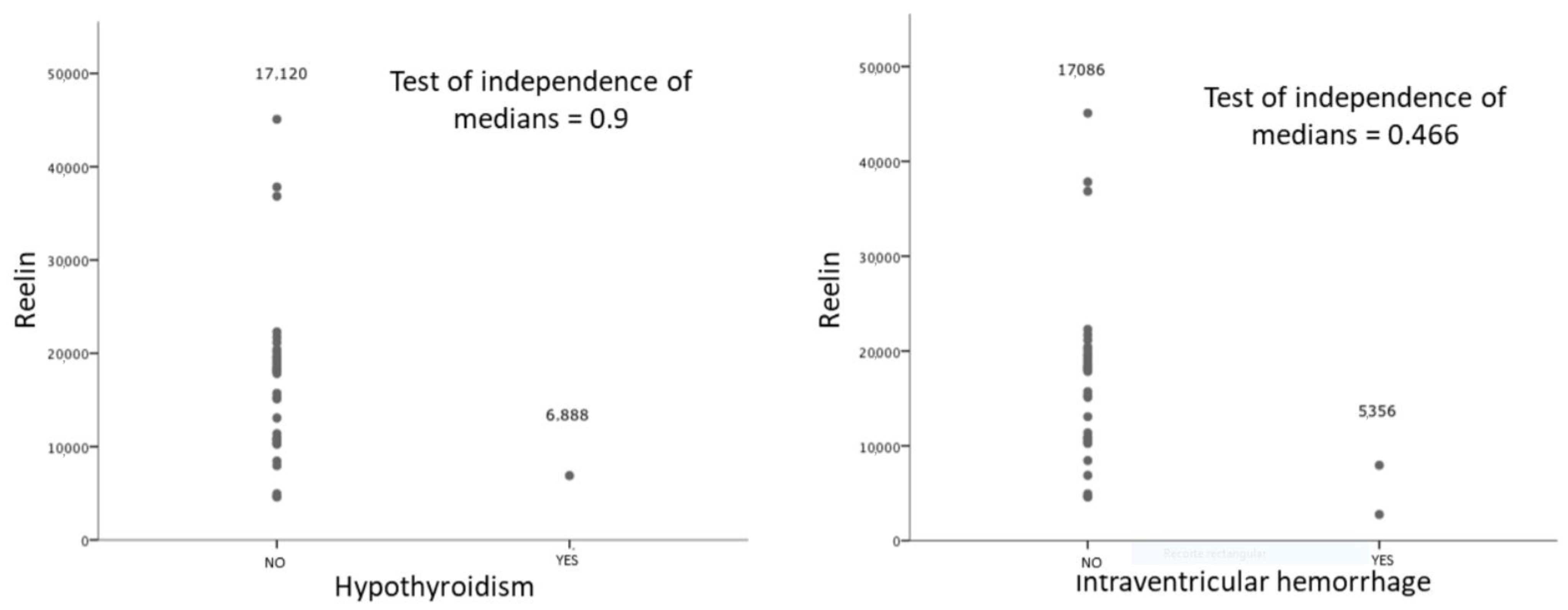
| Intraventricular Hemorrhage No (32) | 0.088 | |
| 16,894(9116) | ||
| Yes (2) | 5356(3675) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascual-Mancho, J.; Pintado-Recarte, P.; Romero-Román, C.; Morales-Camino, J.C.; Hernández-Martin, C.; Bujan, J.; Ortega, M.A.; De León-Luis, J. Influence of Cerebral Vasodilation on Blood Reelin Levels in Growth Restricted Fetuses. Diagnostics 2021, 11, 1036. https://doi.org/10.3390/diagnostics11061036
Pascual-Mancho J, Pintado-Recarte P, Romero-Román C, Morales-Camino JC, Hernández-Martin C, Bujan J, Ortega MA, De León-Luis J. Influence of Cerebral Vasodilation on Blood Reelin Levels in Growth Restricted Fetuses. Diagnostics. 2021; 11(6):1036. https://doi.org/10.3390/diagnostics11061036
Chicago/Turabian StylePascual-Mancho, Jara, Pilar Pintado-Recarte, Carlos Romero-Román, Jorge C. Morales-Camino, Concepción Hernández-Martin, Julia Bujan, Miguel A. Ortega, and Juan De León-Luis. 2021. "Influence of Cerebral Vasodilation on Blood Reelin Levels in Growth Restricted Fetuses" Diagnostics 11, no. 6: 1036. https://doi.org/10.3390/diagnostics11061036
APA StylePascual-Mancho, J., Pintado-Recarte, P., Romero-Román, C., Morales-Camino, J. C., Hernández-Martin, C., Bujan, J., Ortega, M. A., & De León-Luis, J. (2021). Influence of Cerebral Vasodilation on Blood Reelin Levels in Growth Restricted Fetuses. Diagnostics, 11(6), 1036. https://doi.org/10.3390/diagnostics11061036