
Mechanical Intermittent Compression Affects the Progression Rate of Malignant Melanoma Cells in a Cycle Period-Dependent Manner


Abstract
:1. Introduction
2. Materials and Methods
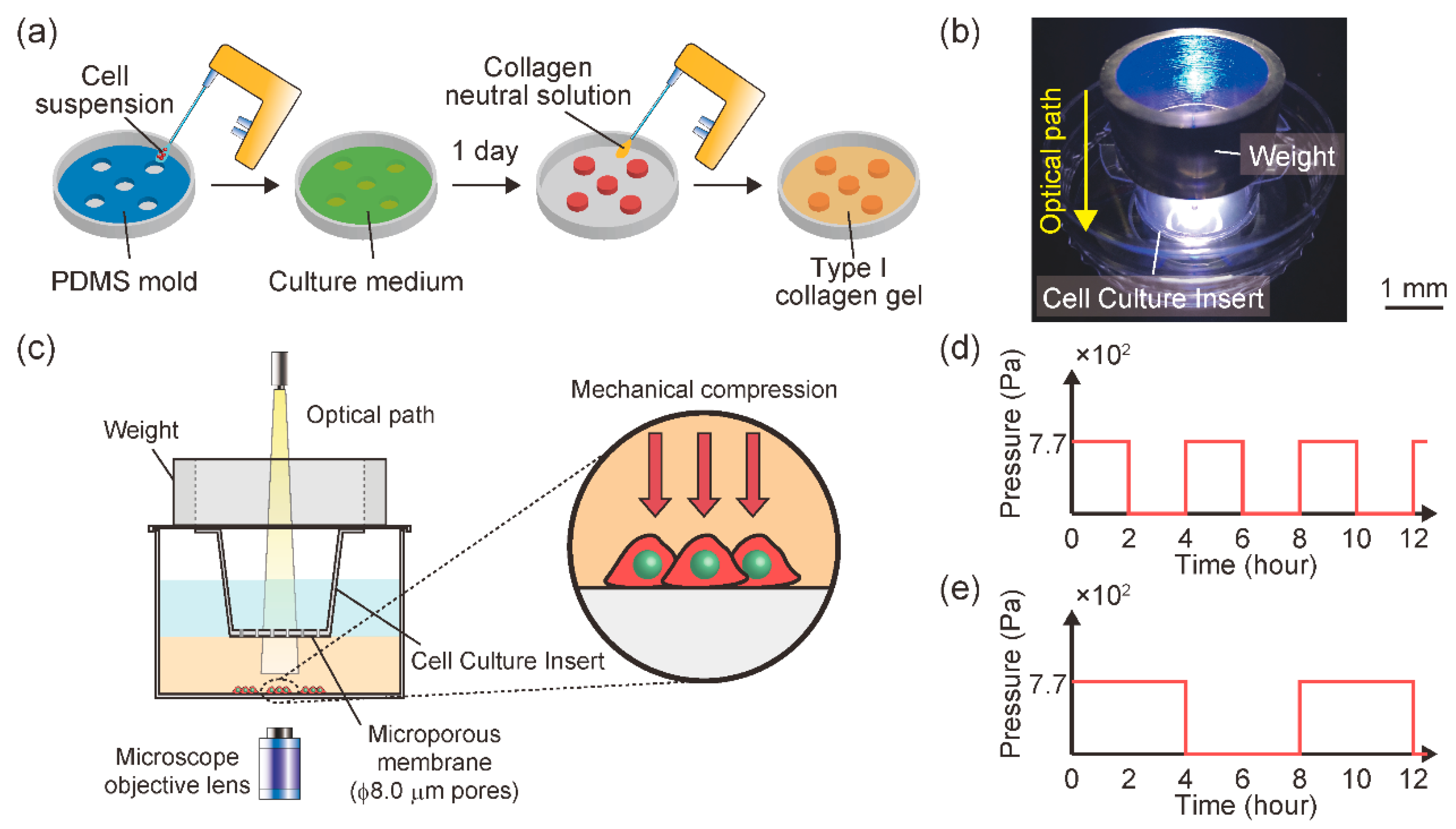
2.1. In Vitro Malignant Melanoma Model to Enable Mechanical Intermittent Compression and Temporal Observation of Cell Behavior
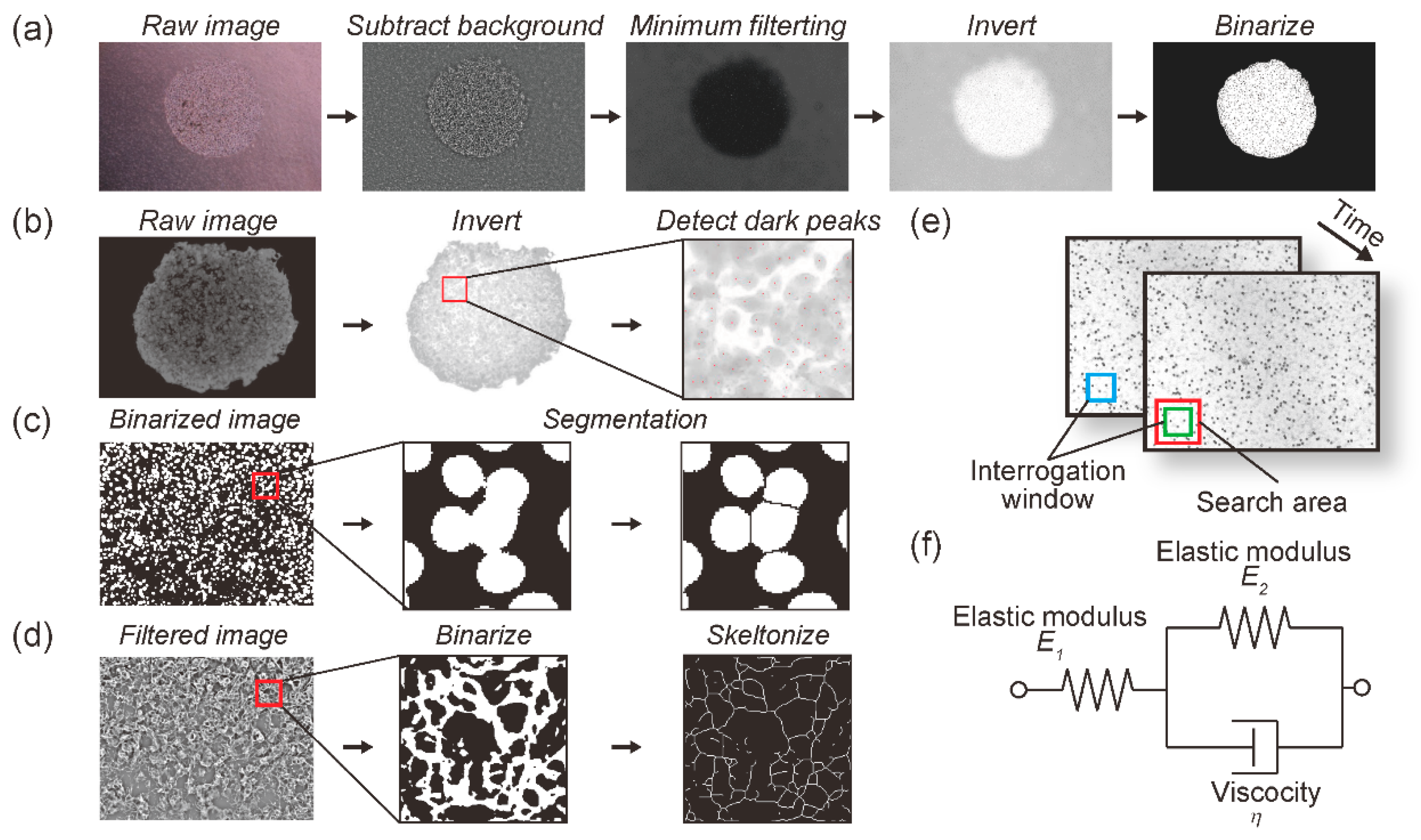
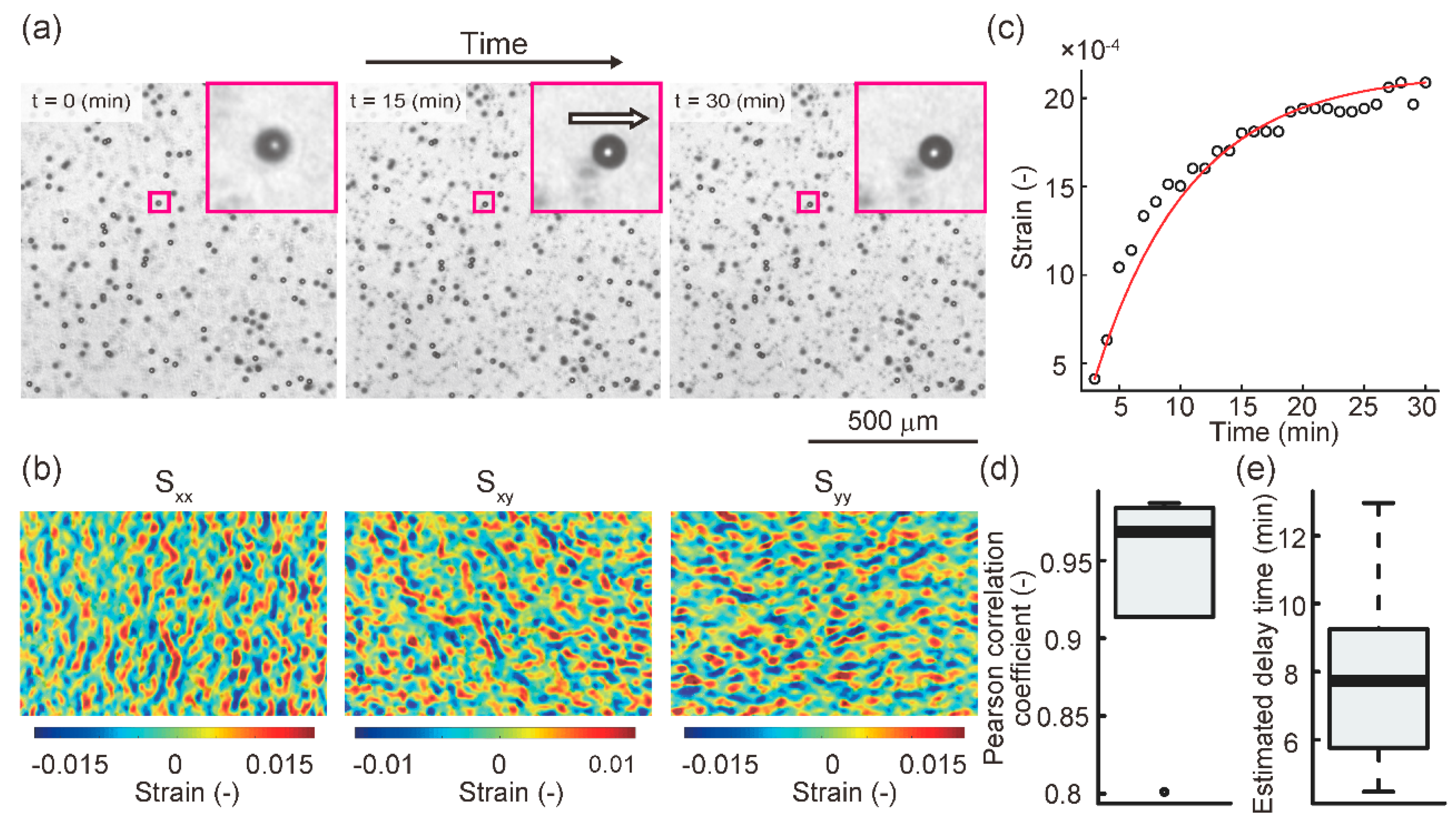
2.2. Creep Phenomenon of Collagen Gel in the Cell Culture Device during Application of Continuous Mechanical Compression
2.3. Quantification of Cell Progression
2.4. Cell Viability and Cell Proliferation Assay
2.5. Fluorescence Staining of F-Actin and Nuclei
2.6. Relative Quantification of mRNA Expression Levels
2.7. Statistical Analysis
3. Results
3.1. Establishment of a Cell Culture Device to Apply Mechanical Intermittent Compression with Temporal Observation
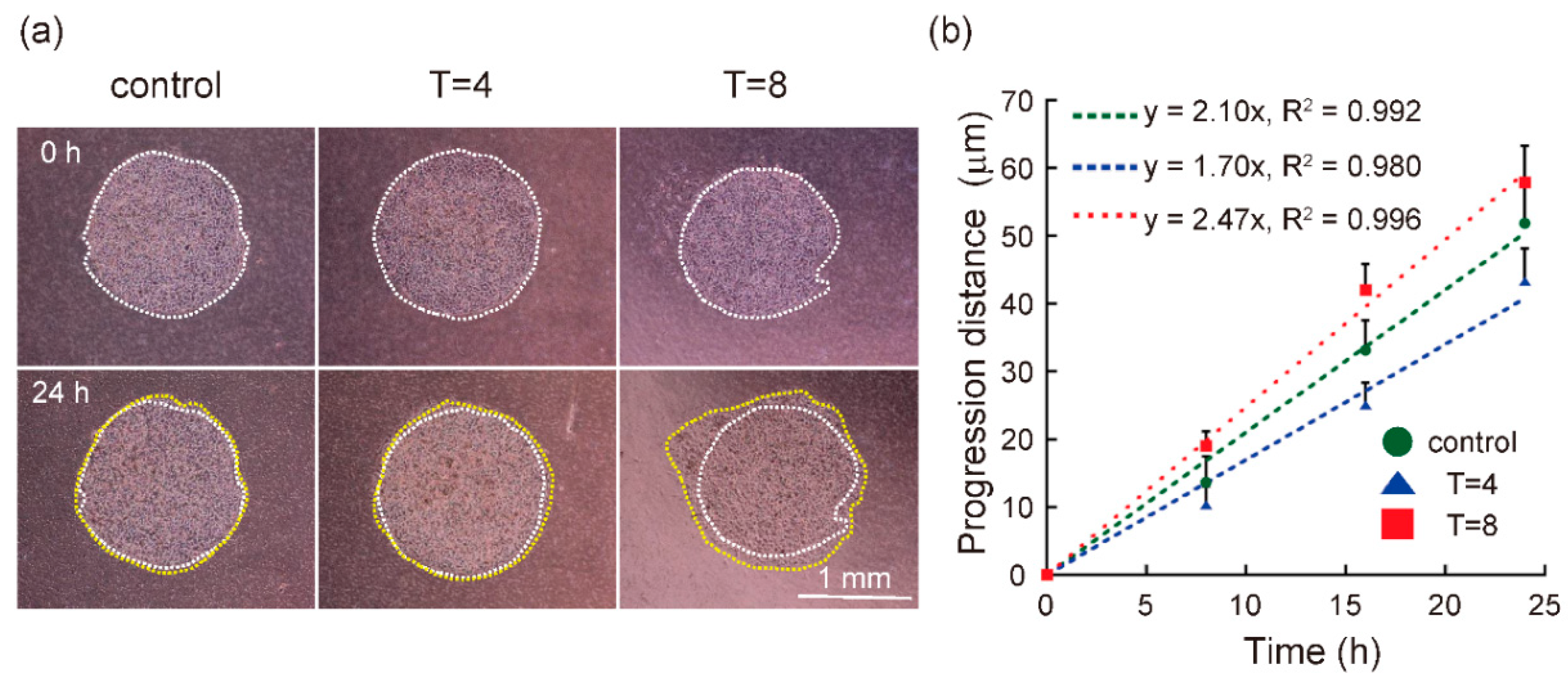
3.2. Progression Rate of Cells in Melanoma Model Was Regulated by Mechanical Intermittent Compression in a Cycle Period-Dependent Manner
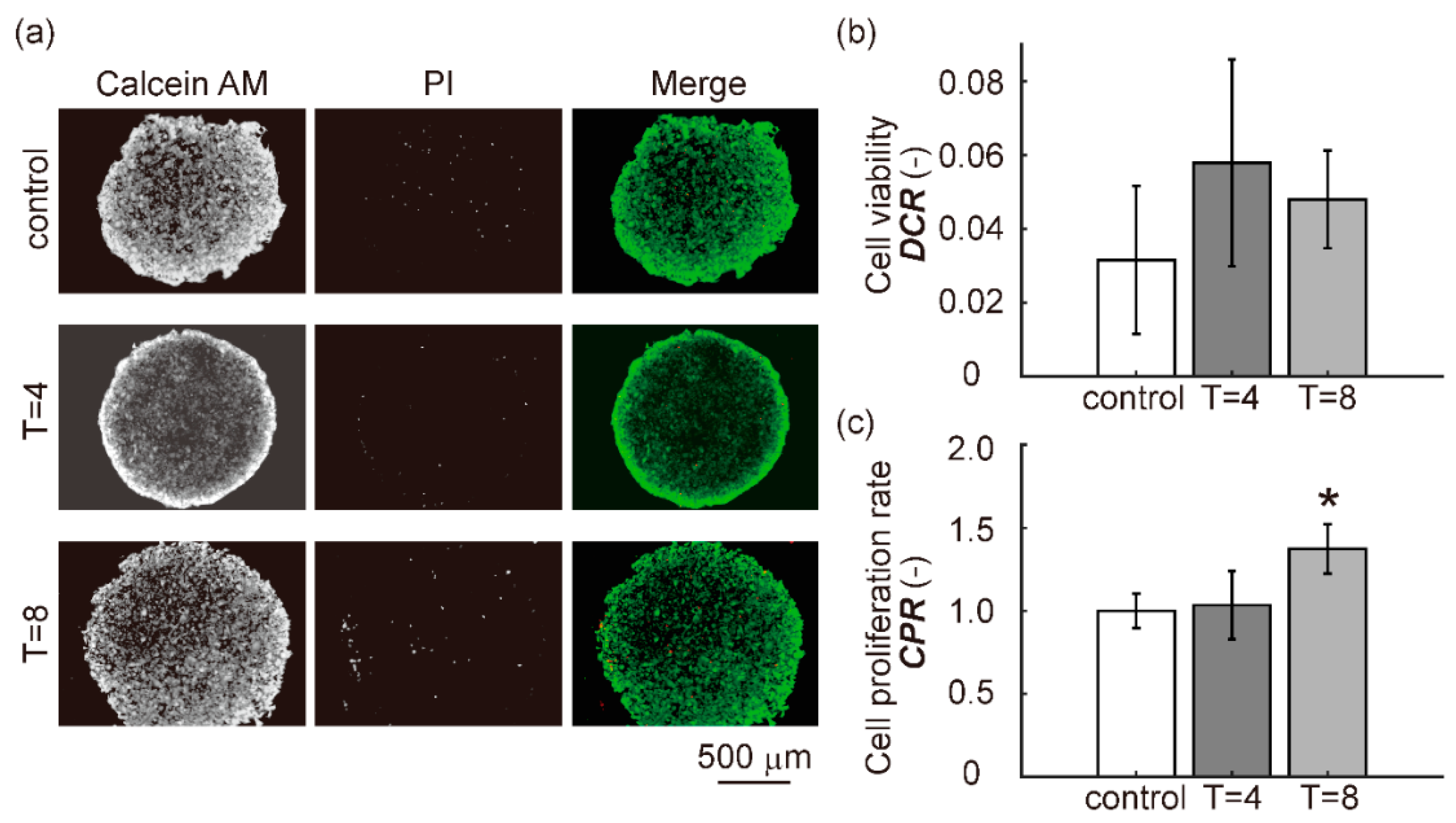
3.3. Cell Viability and Cell Proliferation Rate
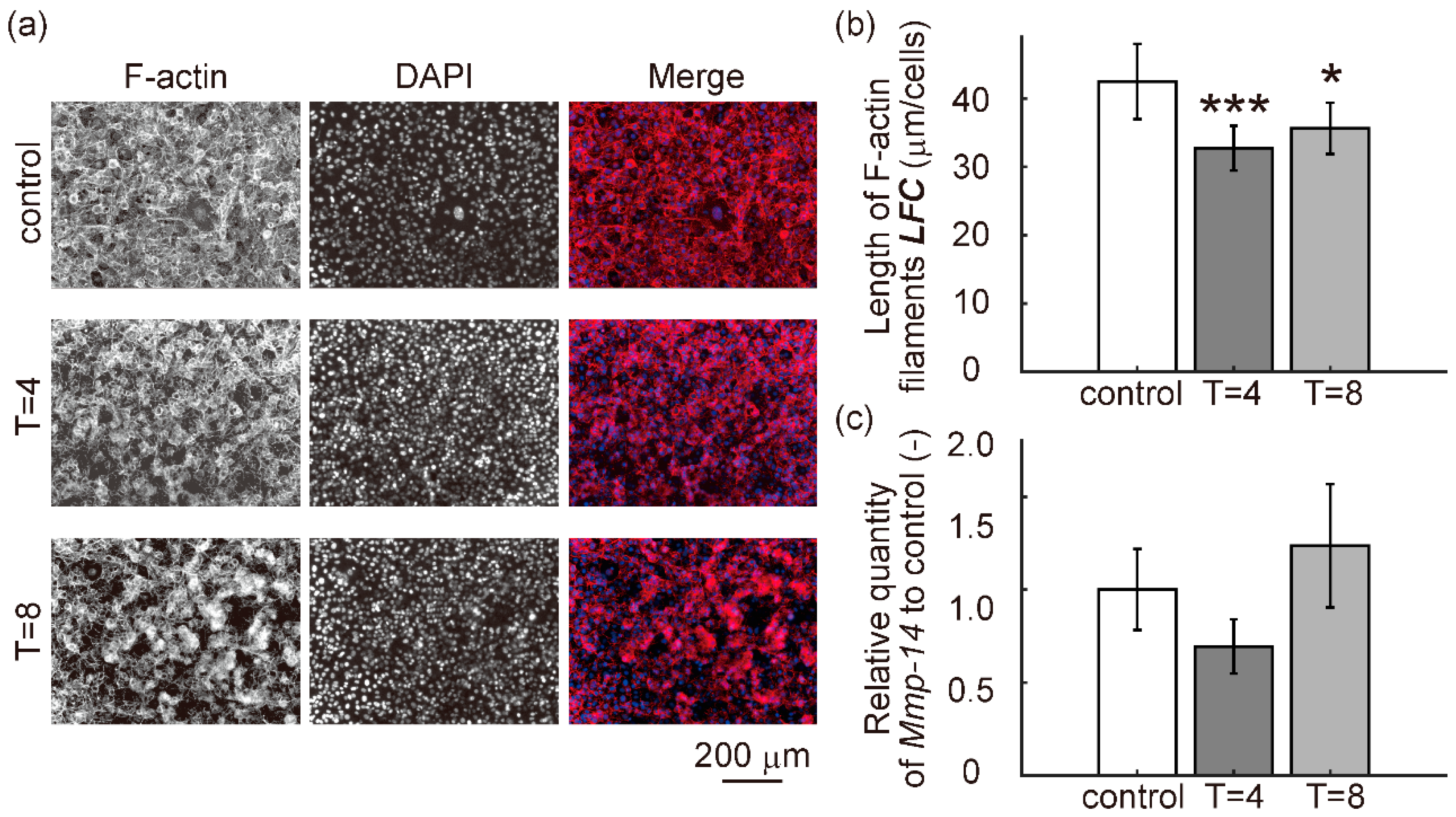
3.4. Cell Migration Capacity
3.5. Relative mRNA Expression Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kalampokas, E.; Kalampokas, T.; Damaskos, C. Primary Vaginal Melanoma, a Rare and Aggressive Entity. A Case Report and Review of the Literature. In Vivo 2017, 31, 133–140. [Google Scholar] [CrossRef] [PubMed]
- de Vries, E.; Bray, F.I.; Coebergh, J.W.W.; Parkin, D.M. Changing Epidemiology of Malignant Cutaneous Melanoma in Europe 1953–1997: Rising Trends in Incidence and Mortality but Recent Stabilizations in Western Europe and Decreases in Scandinavia. Int. J. Cancer 2003, 107, 119–126. [Google Scholar] [CrossRef]
- Stang, A.; Pukkala, E.; Sankila, R.; Söderman, B.; Hakulinen, T. Time Trend Analysis of the Skin Melanoma Incidence of Finland From 1953 Through 2003 Including 16,414 Cases. Int. J. Cancer 2006, 119, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Koh, D.; Wang, H.; Lee, J.; Chia, K.S.; Lee, H.P.; Goh, C.L. Basal Cell Carcinoma, Squamous Cell Carcinoma and Melanoma of the Skin: Analysis of the Singapore Cancer Registry Data 1968–1997. Br. J. Dermatol. 2003, 148, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Domingues, B.; Lopes, J.M.; Soares, P.; Pópulo, H. Melanoma Treatment in Review. Immuno Targets Ther. 2018, 7, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.W.C. Acral Melanoma: A Unique Disease in Asia. JAMA Dermatol. 2013, 149, 1272–1273. [Google Scholar] [CrossRef]
- Borkowska, A.M.; Szumera-Ciećkiewicz, A.; Spałek, M.J.; Teterycz, P.; Czarnecka, A.M.; Rutkowski, P.Ł. Clinicopathological Features and Prognostic Factors of Primary Acral Melanomas in Caucasians. J. Clin. Med. 2020, 9, 2996. [Google Scholar] [CrossRef]
- Guo, J.; Si, L.; Kong, Y.; Flaherty, K.T.; Xu, X.; Zhu, Y.; Corless, C.L.; Li, L.; Li, H.; Sheng, X.; et al. Phase II, Open-Label, Single-Arm Trial of Imatinib Mesylate in Patients With Metastatic Melanoma Harboring c-Kit Mutation or Amplification. J. Clin. Oncol. 2011, 29, 2904–2909. [Google Scholar] [CrossRef]
- Bai, X.; Mao, L.L.; Chi, Z.H.; Sheng, X.N.; Cui, C.L.; Kong, Y.; Dai, J.; Wang, X.; Li, S.M.; Tang, B.X.; et al. BRAF Inhibitors: Efficacious and Tolerable in BRAF-Mutant Acral and Mucosal Melanoma. Neoplasma 2017, 64, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Namikawa, K.; Yoshino, K.; Yoshikawa, S.; Uchi, H.; Goto, K.; Nakamura, Y.; Fukushima, S.; Kiniwa, Y.; Takenouchi, T.; et al. Anti-PD1 Checkpoint Inhibitor Therapy in Acral Melanoma: A Multicenter Study of 193 Japanese Patients. Ann. Oncol. 2020, 31, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.H.; Rapini, R.P. Acral Lentiginous Melanoma. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Bristow, I.R.; Acland, K. Acral Lentiginous Melanoma of the Foot and Ankle: A Case Series and Review of the Literature. J. Foot Ankle Res. 2008, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Albreski, D.; Sloan, S.B. Melanoma of the Feet: Misdiagnosed and Misunderstood. Clin. Dermatol. 2009, 27, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Bristow, I.R.; de Berker, D.A.; Acland, K.M.; Turner, R.J.; Bowling, J. Clinical Guidelines for the Recognition of Melanoma of the Foot and Nail Unit. J. Foot Ankle Res. 2010, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Stalkup, J.R.; Orengo, I.F.; Katta, R. Controversies in Acral Lentiginous Melanoma. Dermatol. Surg. 2002, 28, 1051–1059. [Google Scholar] [PubMed]
- Kwon, I.H.; Lee, J.H.; Cho, K.H. Acral Lentiginous Melanoma In Situ: A Study of Nine Cases. Am. J. Dermatopathol. 2004, 26, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Borkowska, A.; Szumera-Ciećkiewicz, A.; Spałek, M.; Teterycz, P.; Czarnecka, A.; Kowalik, A.; Rutkowski, P. Mutation profile of primary subungual melanomas in Caucasians. Oncotarget 2020, 11, 2404–2413. [Google Scholar] [CrossRef]
- Gandini, S.; Sera, F.; Cattaruzza, M.S.; Pasquini, P.; Picconi, O.; Boyle, P.; Melchi, C.F. Meta-Analysis of Risk Factors for Cutaneous Melanoma: II. Sun Exposure. Eur. J. Cancer 2005, 41, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Hodis, E.; Watson, I.R.; Kryukov, G.V.; Arold, S.T.; Imielinski, M.; Theurillat, J.P.; Nickerson, E.; Auclair, D.; Li, L.; Place, C.; et al. A Landscape of Driver Mutations in Melanoma. Cell 2012, 150, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.M.; Barrett, J.H.; Bishop, D.T.; Armstrong, B.K.; Bataille, V.; Bergman, W.; Berwick, M.; Bracci, P.M.; Elwood, J.M.; Ernstoff, M.S.; et al. Sun Exposure and Melanoma Risk at Different Latitudes: A Pooled Analysis of 5700 Cases and 7216 Controls. Int. J. Epidemiol. 2009, 38, 814–830. [Google Scholar] [CrossRef]
- Curtin, J.A.; Fridlyand, J.; Kageshita, T.; Patel, H.N.; Busam, K.J.; Kutzner, H.; Cho, K.H.; Aiba, S.; Bröcker, E.B.; LeBoit, P.E.; et al. Distinct Sets of Genetic Alterations in Melanoma. N. Engl. J. Med. 2005, 353, 2135–2147. [Google Scholar] [CrossRef]
- Wittekind, C.; Neid, M. Cancer Invasion and Metastasis. Oncology 2005, 69 (Suppl. 1), 14–16. [Google Scholar] [CrossRef]
- Krakhmal, N.V.; Zavyalova, M.V.; Denisov, E.V.; Vtorushin, S.V.; Perelmuter, V.M. Cancer Invasion: Patterns and Mechanisms. Acta Nat. 2015, 7, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Geiger, T.R.; Peeper, D.S. Metastasis Mechanisms. Biochim. Biophys. Acta 2009, 1796, 293–308. [Google Scholar] [CrossRef]
- Cheng, G.; Tse, J.; Jain, R.K.; Munn, L.L. Micro-Environmental Mechanical Stress Controls Tumor Spheroid Size and Morphology by Suppressing Proliferation and Inducing Apoptosis in Cancer Cells. PLoS ONE 2009, 4, e4632. [Google Scholar] [CrossRef] [Green Version]
- Tse, J.M.; Cheng, G.; Tyrrell, J.A.; Wilcox-Adelman, S.A.; Boucher, Y.; Jain, R.K.; Munn, L.L. Mechanical Compression Drives Cancer Cells Toward Invasive Phenotype. Proc. Natl. Acad. Sci. USA 2012, 109, 911–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stucke, S.; McFarland, D.; Goss, L.; Fonov, S.; McMillan, G.R.; Tucker, A.; Berme, N.; Cenk Guler, H.; Bigelow, C.; Davis, B.L. Spatial Relationships Between Shearing Stresses and Pressure on the Plantar Skin Surface During Gait. J. Biomech. 2012, 45, 619–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minagawa, A.; Omodaka, T.; Okuyama, R. Melanomas and Mechanical Stress Points on the Plantar Surface of the Foot. N. Engl. J. Med. 2016, 374, 2404–2406. [Google Scholar] [CrossRef] [Green Version]
- Morikura, T.; Miyata, S. Effect of Mechanical Compression on Invasion Process of Malignant Melanoma Using In Vitro Three-Dimensional Cell Culture Device. Micromachines 2019, 10, 666. [Google Scholar] [CrossRef] [Green Version]
- Chu, T.C.; Ranson, W.F.; Sutton, M.A. Applications of Digital-Image-Correlation Techniques to Experimental Mechanics. Exp. Mech. 1985, 25, 232–244. [Google Scholar] [CrossRef]
- Sutton, M.; Mingqi, C.; Peters, W.; Chao, Y.; McNeill, S. Application of an Optimized Digital Correlation Method to Planar Deformation Analysis. Image Vis. Comput. 1986, 4, 143–150. [Google Scholar] [CrossRef]
- Blaber, J.; Adair, B.; Antoniou, A. Ncorr: Open-Source 2D Digital Image Correlation MATLAB Software. Exp. Mech. 2015, 55, 1105–1122. [Google Scholar] [CrossRef]
- Ortiz, J.S.E.; Lagos, R.E. A Viscoelastic Model to Simulate Soft Tissue Materials. J. Phys. Conf. Ser. 2015, 633, 012099. [Google Scholar] [CrossRef] [Green Version]
- Jayabal, H.; Dingari, N.N.; Rai, B. A Linear Viscoelastic Model to Understand Skin Mechanical Behaviour and for Cosmetic Formulation Design. Int. J. Cosmet. Sci. 2019, 41, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.; Bedogni, B. The Membrane Tethered Matrix Metalloproteinase MT1-MMP at the Forefront of Melanoma Cell Invasion and Metastasis. Pharmacol. Res. 2016, 111, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Barber, R.D.; Harmer, D.W.; Coleman, R.A.; Clark, B.J. GAPDH as a Housekeeping Gene: Analysis of GAPDH mRNA Expression in a Panel of 72 Human Tissues. Physiol. Genom. 2005, 21, 389–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, T.D.; Borisy, G.G. Cellular Motility Driven by Assembly and Disassembly of Actin Filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Carlier, M.F.; Pantaloni, D. Control of Actin Assembly Dynamics in Cell Motility. J. Biol. Chem. 2007, 282, 23005–23009. [Google Scholar] [CrossRef] [Green Version]
- Jacquemet, G.; Hamidi, H.; Ivaska, J. Filopodia in Cell Adhesion, 3D Migration and Cancer Cell Invasion. Curr. Opin. Cell Biol. 2015, 36, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Woodrum, D.T.; Rich, S.A.; Pollard, T.D. Evidence for Biased Bidirectional Polymerization of Actin Filaments Using Heavy Meromyosin Prepared by an Improved Method. J. Cell Biol. 1975, 67, 231–237. [Google Scholar] [CrossRef]
- Pollard, T.D.; Cooper, J.A. Actin and Actin-Binding Proteins. A Critical Evaluation of Mechanisms and Functions. Annu. Rev. Biochem. 1986, 55, 987–1035. [Google Scholar] [CrossRef]
- Suetsugu, S. Shaping the Membrane at Submicron Scale by BAR Proteins and the Actin Cytoskeleton. Seikagaku 2014, 86, 637–649. [Google Scholar]
- Gardel, M.L.; Schneider, I.C.; Aratyn-Schaus, Y.; Waterman, C.M. Mechanical Integration of Actin and Adhesion Dynamics in Cell Migration. Annu. Rev. Cell Dev. Biol. 2010, 26, 315–333. [Google Scholar] [CrossRef] [Green Version]
- Peng, H. Bioimage informatics: A new area of engineering biology. Bioinformatics 2008, 24, 1827–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altadill, A.; Rodríguez, M.; González, L.O.; Junquera, S.; Corte, M.D.; González-Dieguez, M.L.; Linares, A.; Barbón, E.; Fresno-Forcelledo, M.; Rodrigo, L.; et al. Liver expression of matrix metalloproteases and their inhibitors in hepatocellular carcinoma. Dig. Liver Dis. 2009, 41, 740–748. [Google Scholar] [CrossRef]
- Hotary, K.B.; Allen, E.D.; Brooks, P.C.; Datta, N.S.; Long, M.W.; Weiss, S.J. Membrane type I matrix metalloproteinase usurps tumor growth control imposed by the three-dimensional extracellular matrix. Cell 2003, 114, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Koshikawa, N.; Minegishi, T.; Sharabi, A.; Quaranta, V.; Seiki, M. Membrane-type matrix metalloproteinase-1 (MT1-MMP) is a processing enzyme for human laminin gamma 2 chain. J. Biol. Chem. 2005, 280, 88–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiki, M. The cell surface: The stage for matrix metalloproteinase regulation of migration. Curr. Opin. Cell Biol. 2002, 14, 624–632. [Google Scholar] [CrossRef]
- Shaverdashvili, K.; Wong, P.; Ma, J.; Zhang, K.; Osman, I.; Bedogni, B. MT1-MMP modulates melanoma cell dissemination and metastasis through activation of MMP2 and RAC1. Pigment. Cell Melanoma Res. 2014, 27, 287–296. [Google Scholar] [CrossRef]
- Sato, H.; Takino, T.; Okada, Y.; Cao, J.; Shinagawa, A.; Yamamoto, E.; Seiki, M. A matrix metalloproteinase expressed on the surface of invasive tumour cells. Nature 1994, 370, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Kalraiya, R.D. Invasive potential of melanoma cells correlates with the expression of MT1-MMP and regulated by modulating its association with motility receptors via N-glycosylation on the receptors. Biomed. Res. Int. 2014, 2014, 804680. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Bank Accession Number | Sequence(5′-3′) | Tm (°C) | Product Size (bp) |
|---|---|---|---|---|
| Gapdh | NM_001289726.1 | Forward TGTGTCCGTCGTGGATCTGA Reverse TTGCTGTTGAAGTCGCAGGAG | 63.9 63.9 | 3939 |
| Mmp-14 | NM_008608.4 | Forward CCTCAAGTGGCAGCATAATGAGA Reverse TGGCCTCGAATGTGGCATAC | 63.7 64.3 | 83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morikura, T.; Miyata, S. Mechanical Intermittent Compression Affects the Progression Rate of Malignant Melanoma Cells in a Cycle Period-Dependent Manner. Diagnostics 2021, 11, 1112. https://doi.org/10.3390/diagnostics11061112
Morikura T, Miyata S. Mechanical Intermittent Compression Affects the Progression Rate of Malignant Melanoma Cells in a Cycle Period-Dependent Manner. Diagnostics. 2021; 11(6):1112. https://doi.org/10.3390/diagnostics11061112
Chicago/Turabian StyleMorikura, Takashi, and Shogo Miyata. 2021. "Mechanical Intermittent Compression Affects the Progression Rate of Malignant Melanoma Cells in a Cycle Period-Dependent Manner" Diagnostics 11, no. 6: 1112. https://doi.org/10.3390/diagnostics11061112
APA StyleMorikura, T., & Miyata, S. (2021). Mechanical Intermittent Compression Affects the Progression Rate of Malignant Melanoma Cells in a Cycle Period-Dependent Manner. Diagnostics, 11(6), 1112. https://doi.org/10.3390/diagnostics11061112