
Non-Coding RNA Networks as Potential Novel Biomarker and Therapeutic Target for Sepsis and Sepsis-Related Multi-Organ Failure

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Methodology of Literature Search
3. Role of Biomarkers in Sepsis
- As a diagnostic tool, i.e., a biomarker able to confirm a disease;
- As a tool able to stage or to stratify disease severity;
- As a prognostic tool;
- An effective tool for prediction and monitoring of clinical response of an intervention [26].
3.1. C-Reactive Protein (CRP)
3.2. Procalcitonin (PCT)
3.3. Presepsin
4. Non-CodingRNA
4.1. CircRNAs
4.1.1. CircRNAs Biogenesis
4.1.2. CircRNAs Functions
4.2. Long Non-CodingRNAs (lncRNAs)
4.2.1. lncRNAs Biogenesis
4.2.2. lncRNAs Functions
4.3. MicroRNAs (miRNAs)
4.3.1. miRNAs Biogenesis
4.3.2. miRNAs Functions
5. Non-Coding RNA and Sepsis
6. Assessment of the Clinical and Prognostic Value of Non-Coding RNAs as Biomarkers in Sepsis
6.1. circRNAs
6.2. lncRNAs
6.3. miRNAs
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Taeb, A.M.; Hooper, M.H.; Marik, P.E. Sepsis: Current Definition, Pathophysiology, Diagnosis, and Management. Nutr. Clin. Pract. 2017, 32, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuttolomondo, A.; Colomba, C.; Di Bona, D.; Casuccio, A.; Di Raimondo, D.; Clemente, G.; Arnao, V.; Pecoraro, R.; Ragonese, P.; Aiello, A.; et al. HLA and killer cell immunoglobulin-like receptor (KIRs) genotyping in patients with acute viral encephalitis. Oncotarget 2018, 9, 17523–17532. [Google Scholar] [CrossRef] [Green Version]
- Tuttolomondo, A.; Di Raimondo, D.; Pecoraro, R.; Casuccio, A.; Di Bona, D.; Aiello, A.; Accardi, G.; Arnao, V.; Clemente, G.; Della Corte, V.; et al. HLA and killer cell immunoglobulin-like receptor (KIRs) genotyping in patients with acute ischemic stroke. J. Neuroinflamm. 2019, 16, 88. [Google Scholar] [CrossRef] [Green Version]
- Tuttolomondo, A.; Di Raimondo, D.; Pecoraro, R.; Arnao, V.; Pinto, A.; Licata, G. Inflammation in ischemic stroke subtypes. Curr. Pharm. Des. 2012, 18, 4289–4310. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Di Raimondo, D.; di Sciacca, R.; Pinto, A.; Licata, G. Inflammatory cytokines in acute ischemic stroke. Curr. Pharm. Des. 2008, 14, 3574–3589. [Google Scholar] [CrossRef]
- Maida, C.D.; Vasto, S.; Di Raimondo, D.; Casuccio, A.; Vassallo, V.; Daidone, M.; Del Cuore, A.; Pacinella, G.; Cirrincione, A.; Simonetta, I.; et al. Inflammatory activation and endothelial dysfunction markers in patients with permanent atrial fibrillation: A cross-sectional study. Aging 2020, 12, 8423–8433. [Google Scholar] [CrossRef]
- Dounousi, E.; Duni, A.; Naka, K.K.; Vartholomatos, G.; Zoccali, C. The Innate Immune System and Cardiovascular Disease in ESKD: Monocytes and Natural Killer Cells. Curr. Vasc. Pharmacol. 2021, 19, 63–76. [Google Scholar] [CrossRef]
- Pisetsky, D.S.; Gauley, J.; Ullal, A.J. HMGB1 and microparticles as mediators of the immune response to cell death. Antioxid. Redox Signal. 2011, 15, 2209–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Roberts, D.; Wood, K.E.; Light, B.; Parrillo, J.E.; Sharma, S.; Suppes, R.; Feinstein, D.; Zanotti, S.; Taiberg, L.; et al. Duration of hypotension before initiation of effective antimicrobial therapy is the critical determinant of survival in human septic shock. Crit. Care Med. 2006, 34, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.C.; Reinhart, K.; International Sepsis Forum. Biomarkers of sepsis. Crit. Care Med. 2009, 37, 2290–2298. [Google Scholar] [CrossRef] [PubMed]
- Bloos, F.; Reinhart, K. Rapid diagnosis of sepsis. Virulence 2014, 5, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Pierrakos, C.; Vincent, J.L. Sepsis biomarkers: A review. Crit. Care 2010, 14, R15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Flores-Concha, M.; Oñate, Á.A. Long Non-coding RNAs in the Regulation of the Immune Response and Trained Immunity. Front. Genet. 2020, 11, 718. [Google Scholar] [CrossRef]
- Liang, Z.Z.; Guo, C.; Zou, M.M.; Meng, P.; Zhang, T.T. circRNA-miRNA-mRNA regulatory network in human lung cancer: An update. Cancer Cell Int. 2020, 20, 173. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Simonetta, I.; Pinto, A. MicroRNA and receptor mediated signaling pathways as potential therapeutic targets in heart failure. Expert Opin. Ther. Targets 2016, 20, 1287–1300. [Google Scholar] [CrossRef]
- Sakshi, S.; Jayasuriya, R.; Ganesan, K.; Xu, B.; Ramkumar, K.M. Role of circRNA-miRNA-mRNA interaction network in diabetes and its associated complications. Mol. Ther. Nucleic Acids 2021, 26, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Bai, X.; Zeng, X.; Liu, J.; Liu, F.; Zhang, Z. circRNA-miRNA-mRNA in breast cancer. Clin. Chim. Acta Int. J. Clin. Chem. 2021, 523, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Sun, M.L.; Zhang, X.A.; Wang, X.Q. Crosstalk among circRNA/lncRNA, miRNA, and mRNA in Osteoarthritis. Front. Cell Dev. Biol. 2021, 9, 774370. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.L.; Marinov, G.K.; Liau, E.S.; Lam, Y.L.; Lim, Y.Y.; Ea, C.K. Inducible RasGEF1B circular RNA is a positive regulator of ICAM-1 in the TLR4/LPS pathway. RNA Biol. 2016, 13, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Agirre, X.; Meydan, C.; Jiang, Y.; Garate, L.; Doane, A.S.; Li, Z.; Verma, A.; Paiva, B.; Martín-Subero, J.I.; Elemento, O.; et al. Long non-coding RNAs discriminate the stages and gene regulatory states of human humoral immune response. Nat. Commun. 2019, 10, 821. [Google Scholar] [CrossRef] [Green Version]
- Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Black, S.; Kushner, I.; Samols, D. C-reactive Protein. J. Biol. Chem. 2004, 279, 48487–48490. [Google Scholar] [CrossRef] [Green Version]
- Volanakis, J.E. Acute-phase proteins in rheumatic disease. In Arthritis and Allied Conditions: A Textbook of Rheumatology, 13th ed.; Koopman, W.J., Ed.; Williams & Wilkins: Baltimore, MD, USA, 1997; pp. 505–514. [Google Scholar]
- Larsen, F.F.; Petersen, J.A. Novel biomarkers for sepsis: A narrative review. Eur. J. Intern. Med. 2017, 45, 46–50. [Google Scholar] [CrossRef]
- van Vugt, S.F.; Broekhuizen, B.D.; Lammens, C.; Zuithoff, N.P.; de Jong, P.A.; Coenen, S.; Ieven, M.; Butler, C.C.; Goossens, H.; Little, P.; et al. GRACE consortium. Use of serum C reactive protein and procalcitonin concentrations in addition to symptoms and signs to predict pneumonia in patients presenting to primary care with acute cough: Diagnostic study. BMJ 2013, 346, f2450. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; Gauvin, F.; Amre, D.K.; Saint-Louis, P.; Lacroix, J. Serum procalcitonin and C-reactive protein levels as markers of bacterial infection: A systematic review and meta-analysis. Clin. Infect. Dis. 2004, 39, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Hou, J.H.; Li, Q.; Chen, K.J.; Wang, S.N.; Wang, J.M. Biomarkers for diagnosis of sepsis in patients with systemic inflammatory response syndrome: A systematic review and meta-analysis. SpringerPlus 2016, 5, 2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.; Lu, Y.; Jiang, H.; Zhang, L. The diagnostic accuracy of procalcitonin and C-reactive protein for sepsis: A systematic review and meta-analysis. J. Cell. Biochem. 2018, 120, 5852–5859. [Google Scholar] [CrossRef] [PubMed]
- Brunkhorst, F.M.; Eberhard, O.K.; Brunkhorst, R. Discrimination of infectious and noninfectious causes of early acute respiratory distress syndrome by procalcitonin. Crit. Care Med. 1999, 27, 2172–2176. [Google Scholar] [CrossRef]
- Uzzan, B.; Cohen, R.; Nicolas, P.; Cucherat, M.; Perret, G.Y. Procalcitonin as a diagnostic test for sepsis in critically ill adults and after surgery or trauma: A systematic review and meta-analysis. Crit. Care Med. 2006, 34, 1996–2003. [Google Scholar] [CrossRef] [PubMed]
- Schmit, X.; Vincent, J.L. The time course of blood C-reactive protein concentrations in relation to the response to initial antimicrobial therapy in patients with sepsis. Infection 2008, 36, 213–219. [Google Scholar] [CrossRef]
- Assicot, M.; Gendrel, D.; Carsin, H.; Raymond, J.; Guilbaud, J.; Bohuon, C. High serum procalcitonin concentrations in patients with sepsis and infection. Lancet 1993, 341, 515–518. [Google Scholar] [CrossRef]
- Tang, B.M.; Eslick, G.D.; Craig, J.C.; McLean, A.S. Accuracy of procalcitonin for sepsis diagnosis in critically ill patients: Systematic review and meta-analysis. Lancet Infect. Dis. 2007, 7, 210–217. [Google Scholar] [CrossRef]
- Wacker, C.; Prkno, A.; Brunkhorst, F.M.; Schlattmann, P. Procalcitonin as a diagnostic marker for sepsis: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 426–435. [Google Scholar] [CrossRef]
- Memar, M.Y.; Baghi, H.B. Presepsin: A promising biomarker for the detection of bacterial infections. Biomed. Pharmacother. 2019, 111, 649–656. [Google Scholar] [CrossRef]
- Hung, S.K.; Lan, H.M.; Han, S.T.; Wu, C.C.; Chen, K.F. Current Evidence and Limitation of Biomarkers for Detecting Sepsis and Systemic Infection. Biomedicines 2020, 8, 494. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, D.; Liu, Y.N.; Wang, R.; Xie, L.X. The accuracy of presepsin (sCD14-ST) for the diagnosis of sepsis in adults: A meta-analysis. Crit. Care 2015, 19, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Jiang, L.; Ye, L.; Gao, Y.; Tang, L.; Zhang, M. The accuracy of presepsin for the diagnosis of sepsis from SIRS: A systematic review and meta-analysis. Ann. Intensive Care 2015, 5, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hu, Z.D.; Song, J.; Shao, J. Diagnostic Value of Presepsin for Sepsis: A Systematic Review and Meta-Analysis. Medicine 2015, 94, e2158. [Google Scholar] [CrossRef]
- Zeng, Z.; Li, Y.; Pan, Y.; Lan, X.; Song, F.; Sun, J.; Zhou, K.; Liu, X.; Ren, X.; Wang, F.; et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat. Commun. 2018, 9, 5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhu, L.; Bai, M.; Liu, Y.; Zhan, Y.; Deng, T.; Yang, H.; Sun, W.; Wang, X.; Zhu, K.; et al. Exosomal circRNA derived from gastric tumor promotes white adipose browning by targeting the miR-133/PRDM16 pathway. Int. J. Cancer 2019, 144, 2501–2515. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver miR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Shen, T.; Han, M.; Wei, G.; Ni, T. An intriguing RNA species—Perspectives of circularized RNA. Protein Cell 2015, 6, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [Green Version]
- Enuka, Y.; Lauriola, M.; Feldman, M.E.; Sas-Chen, A.; Ulitsky, I.; Yarden, Y. Circular RNAs are long-lived and display only minimal early alterations in response to a growth factor. Nucleic Acids Res. 2016, 44, 1370–1383. [Google Scholar] [CrossRef]
- Harland, R.; Misher, L. Stability of RNA in developing Xenopus embryos and identification of a destabilizing sequence in TFIIIA messenger RNA. Development 1988, 102, 837–852. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kearse, M.G.; Huang, C. The nuclear export of circular RNAs is primarily defined by their length. RNA Biol. 2019, 16, 404–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.Y.; Zhu, Y.R.; Dai, D.J.; Wang, X.; Jin, H.C. Epigenetic regulation of alternative splicing. Am. J. Cancer Res. 2018, 8, 2346–2358. [Google Scholar]
- Chen, L.L.; Yang, L. Regulation of circRNA biogenesis. RNA Biol. 2015, 12, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, D.; Pu, W.; Wang, J.; Peng, Y. Circular RNAs in Cancer: Biogenesis, Function, and Clinical Significance. Trends Cancer 2020, 6, 319–336. [Google Scholar] [CrossRef]
- Zhang, X.O.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Z.; Zhang, J.; Chen, L.L.; Yang, L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Lei, M.; Zheng, G.; Ning, Q.; Zheng, J.; Dong, D. Translation and functional roles of circular RNAs in human cancer. Mol. Cancer 2020, 19, 30. [Google Scholar] [CrossRef]
- Huang, C.; Liang, D.; Tatomer, D.C.; Wilusz, J.E. A length-dependent evolutionarily conserved pathway controls nuclear export of circular RNAs. Genes Dev. 2018, 32, 639–644. [Google Scholar] [CrossRef]
- Piwecka, M.; Glažar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357, eaam8526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef] [PubMed]
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and Characterizing circRNA-Protein Interaction. Theranostics 2017, 7, 4183–4191. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Lv, X. Revealing new landscape of cardiovascular disease through circular RNA-miRNA-mRNA axis. Genomics 2020, 112, 1680–1685. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Tan, W.; Wang, C. The emerging roles of exosomal circRNAs in diseases. Clin. Transl. Oncol. 2021, 23, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar] [CrossRef] [Green Version]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, M.; Tanemura, M.; Akamatsu, H. Long noncoding RNAs regulate malignant phenotypes in colorectal cancer. Biotarget 2018, 2, 4. [Google Scholar] [CrossRef]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The reference human genome annotation for The ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.; Tang, Q.; Sharma, S.; Yu, F.; Escobar, T.M.; Muljo, S.A.; Zhu, J.; Zhao, K. Expression and regulation of intergenic long noncoding RNAs during T cell development and differentiation. Nat. Immunol. 2013, 14, 1190–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, J.; Chang, H. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Wu, H.; Yang, L.; Chen, L.L. The Diversity of Long Noncoding RNAs and Their Generation. Trends Genet. 2017, 33, 540–552. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Chousterman, B.G.; Swirski, F.K.; Weber, G.F. Cytokine storm and sepsis disease pathogenesis. Semin. Immunopathol. 2017, 39, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Grondman, I.; Pirvu, A.; Riza, A.; Ioana, M.; Netea, M.G. Biomarkers of inflammation and the etiology of sepsis. Biochem. Soc. Trans. 2020, 48, 537–546. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, L.; Zhang, H.; Chen, P. Up-regulation of TUG1 can regulate miR-494/PDK4 axis to inhibit LPS-induced acute lung injury caused by sepsis. Am. J. Transl. Res. 2021, 13, 12375–12385. [Google Scholar]
- Qiu, N.; Xu, X.; He, Y. LncRNA TUG1 alleviates sepsis-induced acute lung injury by targeting miR-34b-5p/GAB1. BMC Pulm. Med. 2020, 20, 49. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Fan, G.; Li, Y.; Zhou, Q. TUG1 Represses Apoptosis, Autophagy, and Inflammatory Response by Regulating miR-27a-3p/SLIT2 in Lipopolysaccharide-Treated Vascular Endothelial Cells. J. Surg. Res. 2020, 256, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Chen, W.; Li, W.; Yu, W.; Li, S.; Rao, T.; Ruan, Y.; Zhou, X.; Liu, C.; Qi, Y.; et al. LncRNA TUG1 attenuates ischaemia-reperfusion-induced apoptosis of renal tubular epithelial cells by sponging miR-144-3p via targeting Nrf2. J. Cell. Mol. Med. 2021, 25, 9767–9783. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Niu, Y.; Li, H.; Pan, G. Downregulation of lncRNA TUG1 attenuates inflammation and apoptosis of renal tubular epithelial cell induced by ischemia-reperfusion by sponging miR-449b-5p via targeting HMGB1 and MMP2. Inflammation 2020, 43, 1362–1374. [Google Scholar] [CrossRef]
- Su, Q.; Liu, Y.; Lv, X.W.; Ye, Z.L.; Sun, Y.H.; Kong, B.H.; Qin, Z.B. Inhibition of lncRNA TUG1 upregulates miR-142-3p to ameliorate myocardial injury during ischemia and reperfusion via targeting HMGB1- and Rac1-induced autophagy. J. Mol. Cell. Cardiol. 2019, 133, 12–25. [Google Scholar] [CrossRef]
- Song, T.; Wang, P.; Xin, L. LncRNA TUG1 Contributes to Hypoxia-Induced Myocardial Cell Injury through Downregulating miR-29a-3p in AC16 Cells. J. Cardiovasc. Pharmacol. 2020, 76, 533–539. [Google Scholar] [CrossRef]
- Cai, X.; Wang, S.; Hong, L.; Yu, S.; Li, B.; Zeng, H.; Yang, X.; Zhang, P.; Shao, L. Long Noncoding RNA Taurine-Upregulated Gene 1 Knockdown Protects Cardiomyocytes Against Hypoxia/Reoxygenation-induced Injury through Regulating miR-532-5p/Sox8 Axis. J. Cardiovasc. Pharmacol. 2020, 76, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhao, S.; Li, C.; Liu, C. LncRNA TUG1 serves an important role in hypoxia-induced myocardial cell injury by regulating the miR-145-5p-Binp3 axis. Mol. Med. Rep. 2018, 17, 2422–2430. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Y.; Mo, S.; Wei, W.; Ye, Z.; Su, Q. LncRNA TUG1 competitively binds to miR-340 to accelerate myocardial ischemia-reperfusion injury. FASEB J. 2021, 35, e21163. [Google Scholar] [CrossRef]
- Wei, S.; Liu, Q. Long noncoding RNA MALAT1 modulates sepsis-induced cardiac inflammation through the miR-150-5p/NF-κB axis. Int. J. Clin. Exp. Pathol. 2019, 12, 3311–3319. [Google Scholar] [PubMed]
- Gao, F.; Chen, R.; Xi, Y.; Zhao, Q.; Gao, H. Long noncoding RNA MALAT1 regulates sepsis in patients with burns by modulating miR-214 with TLR5. Mol. Med. Rep. 2019, 19, 3756–3766. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.M.; Liu, G.Q.; Xian, H.B.; Si, J.L.; Qi, S.X.; Yu, Y.P. LncRNA NEAT1 alleviates sepsis-induced myocardial injury by regulating the TLR2/NF-κB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4898–4907. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xue, J.; Qin, L.; Zhang, J.; Liu, J.; Yu, J. LncRNA NEAT1 Promotes Inflammatory Response in Sepsis via the miR-31-5p/POU2F1 Axis. Inflammation 2021, 44, 1518–1528. [Google Scholar] [CrossRef]
- Liu, B.; Ren, H.; Chen, J. LncRNA NEAT1 correlates with Th1 and Th17 and could serve as an assistant biomarker in sepsis. Biomark. Med. 2021, 15, 1177–1186. [Google Scholar] [CrossRef]
- Iro, M.A.; Soundara Pandi, S.P. Clinical application of non-coding RNAs in sepsis. Curr. Opin. Infect. Dis. 2020, 33, 530–539. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Taheri, M.; Arefian, N. Regulatory Role of Non-Coding RNAs on Immune Responses during Sepsis. Front. Immunol. 2021, 12, 798713. [Google Scholar] [CrossRef]
- Wei, B.; Yu, L. Circular RNA PRKCI and microRNA-545 relate to sepsis risk, disease severity and 28-day mortality. Scand. J. Clin. Lab. Investig. 2020, 80, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Liu, J.; Di, X.; Cong, S.; Zhao, M.; Wang, K. Exosomal hsa_circRNA_104484 and hsa_circRNA_104670 may serve as potential novel biomarkers and therapeutic targets for sepsis. Sci. Rep. 2021, 11, 14141. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Li, S.; Wang, J.; Zhao, Z.; Feng, Z. Circular RNA circFADS2 is overexpressed in sepsis and suppresses LPS-induced lung cell apoptosis by inhibiting the maturation of miR-15a-5p. BMC Immunol. 2021, 22, 29. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Wu, J.; Yang, S. Circulating lncRNA ITSN1-2 is upregulated, and its high expression correlates with increased disease severity, elevated inflammation, and poor survival in sepsis patients. J. Clin. Lab. Anal. 2019, 33, e22836. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shao, B. Circulating long noncoding RNA ZNFX1 antisense RNA negatively correlates with disease risk, severity, inflammatory markers, and predicts poor prognosis in sepsis patients. Medicine 2019, 98, e14558. [Google Scholar] [CrossRef]
- Gui, F.; Peng, H.; Liu, Y. Elevated circulating lnc-ANRIL/miR-125a axis level predicts higher risk, more severe disease condition, and worse prognosis of sepsis. J. Clin. Lab. Anal. 2019, 33, e22917. [Google Scholar] [CrossRef]
- Geng, F.; Liu, W.; Yu, L. Potential role of circulating long noncoding RNA MALAT1 in predicting disease risk, severity, and patients’ survival in sepsis. J. Clin. Lab. Anal. 2019, 33, e22968. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fu, X.; Yu, B.; Ai, F. Long non-coding RNA THRIL predicts increased acute respiratory distress syndrome risk and positively correlates with disease severity, inflammation, and mortality in sepsis patients. J. Clin. Lab. Anal. 2020, 33, e22882. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Z.; Liu, B.; Chen, L.; Wang, M. LncRNA GAS5 upregulates miR-214 through methylation to participate in cell apoptosis of sepsis. Arch. Physiol. Biochem. 2020, 1–6. [Google Scholar] [CrossRef]
- Wu, X.; Chen, D.; Yu, L. The value of circulating long non-coding RNA maternally expressed gene 3 as a predictor of higher acute respiratory distress syndrome risk and 28-day mortality in sepsis patients. J. Clin. Lab. Anal. 2020, 34, e23488. [Google Scholar] [CrossRef]
- Chen, J.; He, Y.; Zhou, L.; Deng, Y.; Si, L. Long non-coding RNA MALAT1 serves as an independent predictive biomarker for the diagnosis, severity and prognosis of patients with sepsis. Mol. Med. Rep. 2020, 21, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Geng, F.; Yu, L. Long non-coding RNA MALAT1/microRNA 125a axis presents excellent value in discriminating sepsis patients and exhibits positive association with general disease severity, organ injury, inflammation level, and mortality in sepsis patients. J. Clin. Lab. Anal. 2020, 34, e23222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Yang, L.; Liu, Z.; Wang, Y.; Yang, J. Long noncoding RNA NEAT 1 and its target microRNA-125a in sepsis: Correlation with acute respiratory distress syndrome risk, biochemical indexes, disease severity, and 28-day mortality. J. Clin. Lab. Anal. 2020, 34, e23509. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Feng, Q.; Wu, Y.; Feng, L.; Yuan, H.; Hou, L.; Wei, P.; Wang, C.; Wang, J. Association of circulating long non-coding RNA HULC expression with disease risk, inflammatory cytokines, biochemical index levels, severity-assessed scores, and mortality of sepsis. J. Clin. Lab. Anal. 2021, 35, e23656. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Peng, H.; Gui, F. Long noncoding plasmacytoma variant translocation 1 facilitates the surveillance of acute respiratory distress syndrome and mortality prediction in sepsis. Biomark. Med. 2021, 15, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.F.; Yu, M.L.; Yu, G.; Bian, J.J.; Deng, X.M.; Wan, X.J.; Zhu, K.M. Serum miR-146a and miR-223 as potential new biomarkers for sepsis. Biochem. Biophys. Res. Commun. 2010, 394, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, P.; Chen, W.; Feng, D.; Jia, Y.; Xie, L.X. Evidence for serum miR-15a and miR-16 levels as biomarkers that distinguish sepsis from systemic inflammatory response syndrome in human subjects. Clin. Chem. Lab. Med. 2012, 50, 1423–1428. [Google Scholar] [CrossRef]
- Ma, Y.; Vilanova, D.; Atalar, K.; Delfour, O.; Edgeworth, J.; Ostermann, M.; Hernandez-Fuentes, M.; Razafimahatratra, S.; Michot, B.; Persing, D.H.; et al. Genome-wide sequencing of cellular microRNAs identifies a combinatorial expression signature diagnostic of sepsis. PLoS ONE 2013, 8, e75918. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, H.C.; Chen, C.; Zeng, J.; Wang, Q.; Zheng, L.; Yu, H.D. Differential expression of plasma miR-146a in sepsis patients compared with non-sepsis-SIRS patients. Exp. Ther. Med. 2013, 5, 1101–1104. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, C.; He, Y.; Li, Q.; Wang, G.; Wen, P.; Yang, W.; Yang, Y. Relationship between expression of microRNA and inflammatory cytokines plasma level in pediatric patients with sepsis. Zhonghua Er Ke Za Zhi 2014, 52, 28–33. [Google Scholar]
- Yao, L.; Liu, Z.; Zhu, J.; Li, B.; Chai, C.; Tian, Y. Clinical evaluation of circulating microRNA-25 level change in sepsis and its potential relationship with oxidative stress. Int. J. Clin. Exp. Pathol. 2015, 8, 7675–7684. [Google Scholar] [PubMed]
- Liu, J.; Shi, K.; Chen, M.; Xu, L.; Hong, J.; Hu, B.; Yang, X.; Sun, R. Elevated miR-155 expression induces immunosuppression via CD39+ regulatory T-cells in sepsis patient. Int. J. Infect. Dis. 2015, 40, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Dai, Q.C.; Shen, H.L.; Zhang, X.W. Diagnostic value of elevated serum miRNA-143 levels in sepsis. J. Int. Med. Res. 2016, 44, 875–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; He, J.; Xiao, Z. Value of Micro RNA-499 in prediction of myocardial damage in sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2015, 27, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, J.; Yu, L.; Long, D. Plasma miRNA-223 correlates with risk, inflammatory markers as well as prognosis in sepsis patients. Medicine 2018, 97, e11352. [Google Scholar] [CrossRef]
- Lan, C.; Shi, X.; Guo, N.; Pei, H.; Zhang, H. Value of serum miR-155-5p and miR-133a-3p expression for the diagnosis and prognosis evaluation of sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2016, 28, 694–698. [Google Scholar] [CrossRef]
- Sun, B.; Luan, C.; Guo, L.; Zhang, B.; Liu, Y. Low expression of microRNA-328 can predict sepsis and alleviate sepsis-induced cardiac dysfunction and inflammatory response. Braz. J. Med. Biol. Res. 2020, 53, e9501. [Google Scholar] [CrossRef]
- Guo, H.; Tang, L.; Xu, J.; Lin, C.; Ling, X.; Lu, C.; Liu, Z. MicroRNA-495 serves as a diagnostic biomarker in patients with sepsis and regulates sepsis-induced inflammation and cardiac dysfunction. Eur. J. Med. Res. 2019, 24, 37. [Google Scholar] [CrossRef]
- Zhang, W.; Jia, J.; Liu, Z.; Si, D.; Ma, L.; Zhang, G. Circulating microRNAs as biomarkers for Sepsis secondary to pneumonia diagnosed via Sepsis 3.0. BMC Pulm. Med. 2019, 19, 93. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Liu, X.; Ni, H. Clinical significance of miR-19b-3p in patients with sepsis and its regulatory role in the LPS-induced inflammatory response. Eur. J. Med. Res. 2020, 25, 9. [Google Scholar] [CrossRef]
- Lin, R.; Hu, H.; Li, L.; Chen, G.; Luo, L.; Rao, P. The potential of microRNA-126 in predicting disease risk, mortality of sepsis, and its correlation with inflammation and sepsis severity. J. Clin. Lab. Anal. 2020, 34, e23408. [Google Scholar] [CrossRef] [PubMed]
- Rahmel, T.; Schäfer, S.T.; Frey, U.H.; Adamzik, M.; Peters, J. Increased circulating microRNA-122 is a biomarker for discrimination and risk stratification in patients defined by sepsis-3 criteria. PLoS ONE 2018, 13, e0197637. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Qiu, G.; Ge, M.; Meng, J.; Zhang, G.; Wang, J.; Huang, R.; Shu, Q.; Xu, J. miR-10a in Peripheral Blood Mononuclear Cells Is a Biomarker for Sepsis and Has Anti-Inflammatory Function. Mediat. Inflamm. 2020, 2020, 4370983. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X. MiR-125b but not miR-125a is upregulated and exhibits a trend to correlate with enhanced disease severity, inflammation, and increased mortality in sepsis patients. J. Clin. Lab. Anal. 2020, 34, e23094. [Google Scholar] [CrossRef] [Green Version]
- Karam, R.A.; Zidan, H.E.; Karam, N.A.; Abdel Rahman, D.M.; El-Seifi, O.S. Diagnostic and prognostic significance of serum miRNA-146-a expression in Egyptian children with sepsis in a pediatric intensive care unit. J. Gene Med. 2019, 21, e3128. [Google Scholar] [CrossRef]
- Liu, G.; Liu, W.; Guo, J. Clinical significance of miR-181a in patients with neonatal sepsis and its regulatory role in the lipopolysaccharide-induced inflammatory response. Exp. Ther. Med. 2020, 19, 1977–1983. [Google Scholar] [CrossRef] [Green Version]
- Salim, R.F.; Sobeih, A.A.; Abd El Kareem, H.M. Evaluation of the clinical value of circulating miR-101, miR-187 and miR-21 in neonatal sepsis diagnosis and prognosis. Egypt. J. Med. Hum. Genet. 2020, 21, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.F.; Li, Y.; Wang, Y.Q.; Li, H.J.; Dou, L. MicroRNA-494-3p alleviates inflammatory response in sepsis by targeting TLR6. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2971–2977. [Google Scholar] [CrossRef]
- Abou El-Khier, N.T.; Zaki, M.E.; Alkasaby, N.M. Study of MicroRNA-122 as a Diagnostic Biomarker of Sepsis. Egypt. J. Immunol. 2019, 26, 105–116. [Google Scholar]
- Na, L.; Ding, H.; Xing, E.; Zhang, Y.; Gao, J.; Liu, B.; Yu, J.; Zhao, Y. The predictive value of microRNA-21 for sepsis risk and its correlation with disease severity, systemic inflammation, and 28-day mortality in sepsis patients. J. Clin. Lab. Anal. 2020, 34, e23103. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, Q.; Zhang, Y.; Zhou, S.; Chen, H. Decreased microRNA 103 and microRNA 107 predict increased risks of acute respiratory distress syndrome and 28-day mortality in sepsis patients. Medicine 2020, 99, e20729. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yu, L.; Zhang, R.; Zhu, L.; Shen, W. Correlation of microRNA-146a/b with disease risk, biochemical indices, inflammatory cytokines, overall disease severity, and prognosis of sepsis. Medicine 2020, 99, e19754. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Li, S.; Cui, J.; Wang, L.; Ma, X.; Li, Y. Plasma miR-125a and miR-125b in sepsis: Correlation with disease risk, inflammation, severity, and prognosis. J. Clin. Lab. Anal. 2020, 34, e23036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Ding, Y.; Song, S.; Li, M.; Wang, T.; Guo, F. Expression patterns and prognostic value of miR-210, miR-494, and miR-205 in middle-aged and old patients with sepsis-induced acute kidney injury. Bosn. J. Basic Med. Sci. 2019, 19, 249–256. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, D.; Gao, J.; Xiang, X.; Hu, X.; Li, S.; Wu, W.; Cai, J.; Tang, C.; Zhang, D.; et al. Discovery and validation of miR-452 as an effective biomarker for acute kidney injury in sepsis. Theranostics 2020, 10, 11963–11975. [Google Scholar] [CrossRef]
- Li, S.; Zhao, D.; Cui, J.; Wang, L.; Ma, X.; Li, Y. Correlation of microRNA-125a/b with acute respiratory distress syndrome risk and prognosis in sepsis patients. J. Clin. Lab. Anal. 2020, 34, e23098. [Google Scholar] [CrossRef]
- Zhang, B.; Yu, L.; Sheng, Y. Clinical value and role of microRNA-29c-3p in sepsis-induced inflammation and cardiac dysfunction. J. Clin. Lab. Anal. 2021, 26, 90. [Google Scholar] [CrossRef]


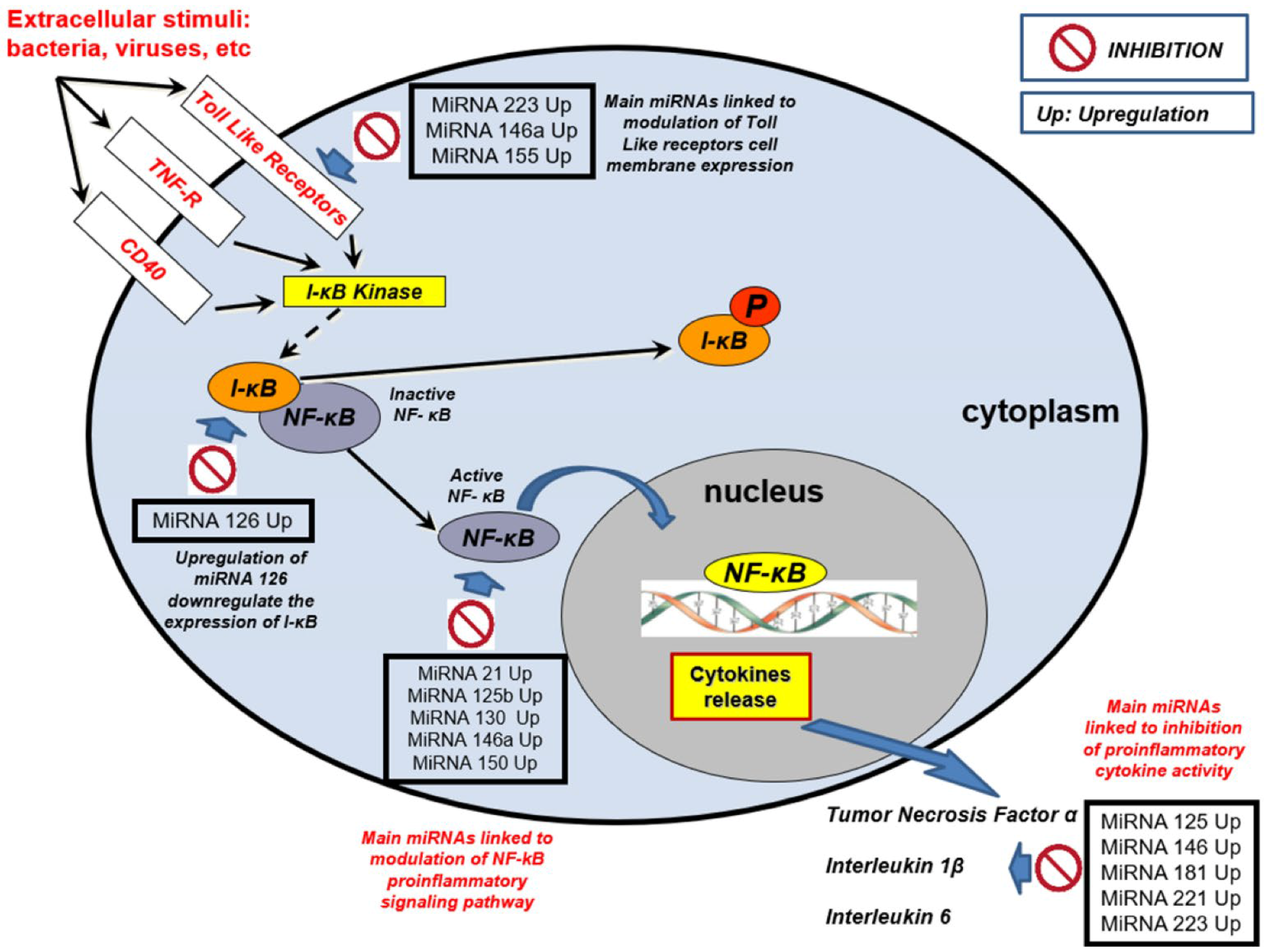{kind=link}
| Ref. | Year | lncRNA | Novel/Validation | Pattern of Expression | Sample Size | Diagnostic Power | Other Results | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sepsis Cases | Control | N° | 1. Sensitivity (%) | 2. Specificity (%) | AUC | ||||||
| [105] | 2019 | lncRNA ITSN1-2 | Validation | Upregulated | 309 | HC | 300 | 59.5 | 86.3 | 0.777 | Positive correlation with APACHE II, CRP, TNF-α, IL-6, il-8; Negative correlation with IL-10 |
| [106] | 2019 | lncRNA ZFAS1 | Novel | Downregulated | 202 | HC | 200 | NR | NR | 0.814 | Negative correlation with APACHE II, CRP, TNF-α, IL-6; Positive correlation with IL-10; predicts survivor from non-survivor |
| [107] | 2019 | lncRNA ANRIL | Novel | Upregulated | 126 | HC | 126 | NR | NR | 0.800 | Positive correlation with CRP, PCT, APACHE II, SOFA, TNF-α, IL-8 |
| [108] | 2019 | lncRNA MALAT1 | Novel | Upregulated | 190 | HC | 190 | NR | NR | 0.823 | Positive correlation with PCT, Scr, WBC, CRP, SOFA and APACHE II; predict 28-day mortality |
| [109] | 2020 | lncRNA THRIL | Novel | Upregulated | 32 ARDS +sepsis | nonARDS-sepsis | 77 | NR | NR | 0.706 | Positive correlation with CRP, PCT, TNF-α, IL-1β |
| [110] | 2020 | lncRNA GAS5 | Novel | Downregulated | 60 | HC | 60 | NR | NR | NR | Positive correlation with miRNA-214 |
| [111] | 2020 | lncRNA MEG3 | Validation | Upregulated | 112 | HC | 100 | 77.7 | 94 | 0.893 | Predictive role for ARDS-sepsis |
| [112] | 2020 | lncRNA MALAT1 | Validation | Upregulated | 120 | HC | 60 | NR | NR | 0.910 | Positive correlation with PCT, Lactate levels, SOFA and APACHE II |
| [113] | 2020 | lncRNA MALAT1 | Validation | Upregulated | 196 | HC | 196 | 91.3 | 78.6 | 0.931 | Negative correlation with miR125a and albumin; positive correlation with APACHE II, SOFA, Scr, CRP, IL-6, IL-8, IL-1β, TNF-α |
| [114] | 2020 | lncRNA NEAT1 | Validation | Upregulated | 102 | HC | 100 | NR | NR | 0.992 | Negative correlation with miR-125a |
| [115] | 2021 | lncRNA HULC | Novel | Upregulated | 174 | HC | 100 | 78.7 | 97 | 0.939 | Positive correlation with TNF-α, IL-6, IL-17, ICAM1, and VCAM1 APACHE II, SOFA Score, |
| [116] | 2021 | lncRNA PVT1 | Validation | Upregulated | 109 | HC | 100 | NR | NR | NR | Predictive role for ARDS and 28-day mortality, positive correlation with disease severity; |
| Ref. | Year | miRNA | Novel/Validation | Pattern of Expression | Sample Size | Diagnostic Power | Other Results | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sepsis Cases | Control | Number | Sensitivity (%) | Specificity (%) | AUC | ||||||
| [117] | 2009 | miR-146a | Novel | Downregulated | 50 | SIRS + HCs | 30 + 20 | NR | NR | 0.804 | N/A |
| [117] | 2009 | miR-223 | Novel | Downregulated | 50 | SIRS + HCs | 30 + 20 | NR | NR | 0.858 | N/A |
| [118] | 2012 | miR-15a | Novel | Downregulated | 166 | SIRS | 32 | 68.3 | 94.4 | 0.858 | N/A |
| [119] | 2013 | miR-150 | Novel | Upregulated | 23 | SIRS | 22 | 72.7 | 85.7 | 0.830 | N/A |
| [119] | 2013 | miR-4772-5p-iso | Novel | Downregulated | 23 | SIRS | 22 | 68.2 | 71.4 | 0.760 | N/A |
| [120] | 2013 | miR-146a | Validation | Downregulated | 14 | SIRS | 14 | 60 | 87.5 | 0.813 | N/A |
| [121] | 2013 | miR-146a | Validation | Upregulated | 40 | SIRS | 20 | 77.5 | 77 | 0.815 | Positive correlation with miR-223, IL-10, TNF-α |
| [121] | 2013 | miR-123 | Novel | Upregulated | 40 | SIRS | 20 | 77.5 | 55 | 0.678 | Positive correlation with miR-146a, IL-10, TNF-α |
| [122] | 2014 | miR-25 | Novel | Downregulated | 70 | SIRS | 30 | NR | NR | 0.806 | Negative correlation with SOFA, PCT, CRP. Predictive role in 28-day mortality risk (AUC: 0.756) |
| [123] | 2014 | miR-155 | Novel | Upregulated | 60 | HCs | 30 | NR | NR | NR | Positive correlation with SOFA; predictive role in 28-days mortality risk (AUC: 763) |
| [124] | 2014 | miR-143 | Novel | Upregulated | 103 | SIRS | 95 | 78.6 | 91.6 | 0.910 | Positive correlation with SOFA, APACHE II |
| [125] | 2015 | miR-499 | Novel | Upregulated | 112 | HCs | 20 | 86.7 | 90.8 | 0.838 | N/A |
| [126] | 2016 | miR-223 | Validation | Upregulated | 187 | HCs | 186 | 56.6 | 86.6 | 0.754 | Positive correlation with CRP, TNF-α, IL-1β, IL-6, IL-8 and negatively with IL-10; predicts survivor from non-survivor |
| [127] | 2016 | miR-155-5p | Validation | Upregulated | 105 | HCs | 35 | 85.3 | 80.6 | 0.855 | N/A |
| [127] | 2016 | miR-133a-3p | Novel | Upregulated | 105 | HCs | 35 | 97.9 | 54.8 | 0.769 | N/A |
| [128] | 2017 | miR-328 | Novel | Upregulated | 110 | HCs | 89 | 87.6 | 86.4 | 0.926 | Positive correlation with Scr, WBC, CRP, PTC, APACHE II, SOFA, |
| [129] | 2017 | miR-495 | Novel | Downregulated | 105 | HCs | 100 | 89.5 | 83 | 0.915 | Distinguishes sepsis from sepsis shock (Sen: 85.3%; Spec: 87.3; AUC 0.885); Negative correlation with Scr, WBC, CRP, PCT, APACHE II, SOFA |
| [130] | 2017 | miR-7110-5p | Novel | Upregulated | 44 | Non sepsis pneumonia + HC | 96 | 84.2 | 90.5 | 0.883 | N/A |
| [130] | 2017 | miR-223-3p | Validation | Upregulated | 44 | Non sepsis pneumonia + HCs | 96 | 82.9 | 100 | 0.964 | N/A |
| [131] | 2017 | miR-19b-3p | Novel | Downregulated | 103 | HCs | 98 | 85.4 | 85.7 | 0.921 | Independent prognostic factor for 28-days survival; Negative correlation with IL-6, TNF-α |
| [132] | 2018 | miR-126 | Novel | Upregulated | 208 | HCs | 210 | NR | NR | 0.726 | Positive correlation with APACHE II, ICU stay, MCD, Scr, CRP, TNF-α, IL-6, IL-8 and negative with IL-10 |
| [133] | 2018 | miR-122 | Validation | Upregulated | 108 | Non sepsis infection | 20 | 58.3 | 95 | 0.760 | Independent prognostic factor for 30-days mortality (HR: 4.3) |
| [134] | 2018 | miR-10a | Novel | Downregulated | 62 | HCs | 20 | NR | NR | 0.804 | Negative correlation with APACHE II, SOFA, CRP, PCT; predictive role in 28-days mortality risk (AUC: 0.795) |
| [135] | 2018 | miR-125b | Novel | Upregulated | 120 | HCs | 120 | 49.2 | 80 | 0.658 | Positive correlation with APACHE II, SOFA, Scr, CRP, PCT, TNF-α, IL-6; Independent factor for mortality risk. In this study miR-125a upregulation was not associated with enhanced disease severity, inflammation, and increased mortality in sepsis patients |
| [136] | 2018 | miR-146a | Validation | Downregulated | 55 | HCs | 60 | 86.6 | 56.6 | 0.803 | Negative correlation with CRP, PCT, IL-6, TNF-α |
| [137] | 2018 | miR-181a | Novel | Downregulated | 102 | Local infection | 50 | 83.3 | 84 | 0.893 | N/A |
| [138] | 2018 | miR-101 | Novel | Upregulated | 50 | SIRS | 30 | 84 | 84 | 0.908 | N/A |
| [138] | 2018 | miR-187 | Novel | Upregulated | 50 | SIRS | 30 | 72 | 76 | 0.789 | N/A |
| [138] | 2018 | miR-21 | Novel | Upregulated | 50 | SIRS | 30 | 64 | 66 | 0.711 | N/A |
| [139] | 2019 | miR-494-3p | Novel | Downregulated | NR | HCs | NR | NR | NR | 0.837 | N/A |
| [140] | 2019 | miR-122 | Novel | Upregulated | 25 | LWI | 25 | 100 | 100 | 1.000 | Higher AUC than CRP and WBC; 56% of accuracy as a prognostic biomarker |
| [141] | 2019 | miR-21 | Validation | Downregulated | 219 | HCs | 219 | NR | NR | 0.801 | Negative correlation with APACHE II, SOFA, Scr, CRP, TNF-α, IL-1β, IL-6, IL-17; |
| [142] | 2019 | miR-103 | Novel | Downregulated | 196 | HCs | 196 | NR | NR | NR | Negative correlation with APACHE II, SOFA, Scr, CRP, TNF, IL-1β, IL-6, IL-8 positive with albumin; predicted high ARDS risk (AUC: 0.727) and increased 28-days mortality risk (AUC: 0.704) |
| [142] | 2019 | miR-107 | Novel | Downregulated | 196 | HC | 196 | NR | NR | NR | Negative correlation with APACHE II, SOFA, Scr, CRP, TNF, IL-1β, IL-6, IL-8 positive with albumin; predicted high ARDS risk (AUC: 0.694) and increased 28-days mortality risk (AUC: 0.649) |
| [143] | 2019 | miR-146a | Validation | Upregulated | 180 | HCs | 180 | NR | NR | 0.774 | Positive correlation with APACHE II, SOFA, Scr, CRP, TNF-α, IL-1β, IL-6, IL17 and negative with albumin |
| [143] | 2019 | miR-146b | Novel | Upregulated | 180 | HCs | 180 | NR | NR | 0.897 | Good predictive value in 28-days mortality risk (AUC: 0.703);Positive correlation with APACHE II, SOFA, Scr, CRP, TNF-α, IL-1β, IL-6, IL17 and negative with albumin |
| [144] | 2019 | miR-125a | Novel | Upregulated | 150 | HCs | 150 | NR | NR | 0.749 | Positive correlation with APACHE II, SOFA. Not correlates with level of inflammation, disease severity, and 28-day mortality risk in sepsis patients |
| [144] | 2019 | miR-125b | Validation | Upregulated | 150 | HCs | 150 | NR | NR | 0.839 | Positive correlation with APACHE II, SOFA, CRP, TNF-α, IL-6, IL-17, IL-23; predictive role in 28-days mortality risk (AUC: 0.699) |
| [145] | 2019 | miR-210 | Novel | Upregulated | 125 | HCs | 110 | 81 | 80.9 | 0.852 | Positive correlation with BUN, Scr, CysC |
| [145] | 2019 | miR-494 | Validation | Upregulated | 125 | HCs | 110 | 80.9 | 72.1 | 0.847 | Positive correlation with BUN, Scr, CysC |
| [145] | 2019 | miR-205 | Novel | Upregulated | 125 | HCs | 110 | 78.6 | 90.5 | 0.860 | Negative correlation with BUN, Scr, CysC |
| [146] | 2020 | miR-452 | Novel | Upregulated | 97 | HCs | 89 | NR | NR | NR | High efficacy in distinguishing AKI in sepsis patients |
| [147] | 2020 | miR-125a | Validation | Upregulated | 41 | noARDS-sepsis | 109 | NR | NR | 0.650 | Positive correlation with Scr, APACHE II, SOFA |
| [147] | 2020 | miR-125b | Validation | Upregulated | 41 | noARDS-sepsis | 109 | NR | NR | 0.739 | Positive correlation with with Scr, APACHE II, SOFA |
| [148] | 2021 | miR-29c-3p | Novel | Upregulated | 86 | HCs | 85 | 80.2 | 81.1 | 0.872 | Positive correlation with APACHE II, SOFA, CRP, PCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Raimondo, D.; Pirera, E.; Rizzo, G.; Simonetta, I.; Musiari, G.; Tuttolomondo, A. Non-Coding RNA Networks as Potential Novel Biomarker and Therapeutic Target for Sepsis and Sepsis-Related Multi-Organ Failure. Diagnostics 2022, 12, 1355. https://doi.org/10.3390/diagnostics12061355
Di Raimondo D, Pirera E, Rizzo G, Simonetta I, Musiari G, Tuttolomondo A. Non-Coding RNA Networks as Potential Novel Biomarker and Therapeutic Target for Sepsis and Sepsis-Related Multi-Organ Failure. Diagnostics. 2022; 12(6):1355. https://doi.org/10.3390/diagnostics12061355
Chicago/Turabian StyleDi Raimondo, Domenico, Edoardo Pirera, Giuliana Rizzo, Irene Simonetta, Gaia Musiari, and Antonino Tuttolomondo. 2022. "Non-Coding RNA Networks as Potential Novel Biomarker and Therapeutic Target for Sepsis and Sepsis-Related Multi-Organ Failure" Diagnostics 12, no. 6: 1355. https://doi.org/10.3390/diagnostics12061355
APA StyleDi Raimondo, D., Pirera, E., Rizzo, G., Simonetta, I., Musiari, G., & Tuttolomondo, A. (2022). Non-Coding RNA Networks as Potential Novel Biomarker and Therapeutic Target for Sepsis and Sepsis-Related Multi-Organ Failure. Diagnostics, 12(6), 1355. https://doi.org/10.3390/diagnostics12061355