
Genomic Breakpoints’ Characterization of a Large CHEK2 Duplication in an Italian Family with Hereditary Breast Cancer

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Case Presentation
3. Results
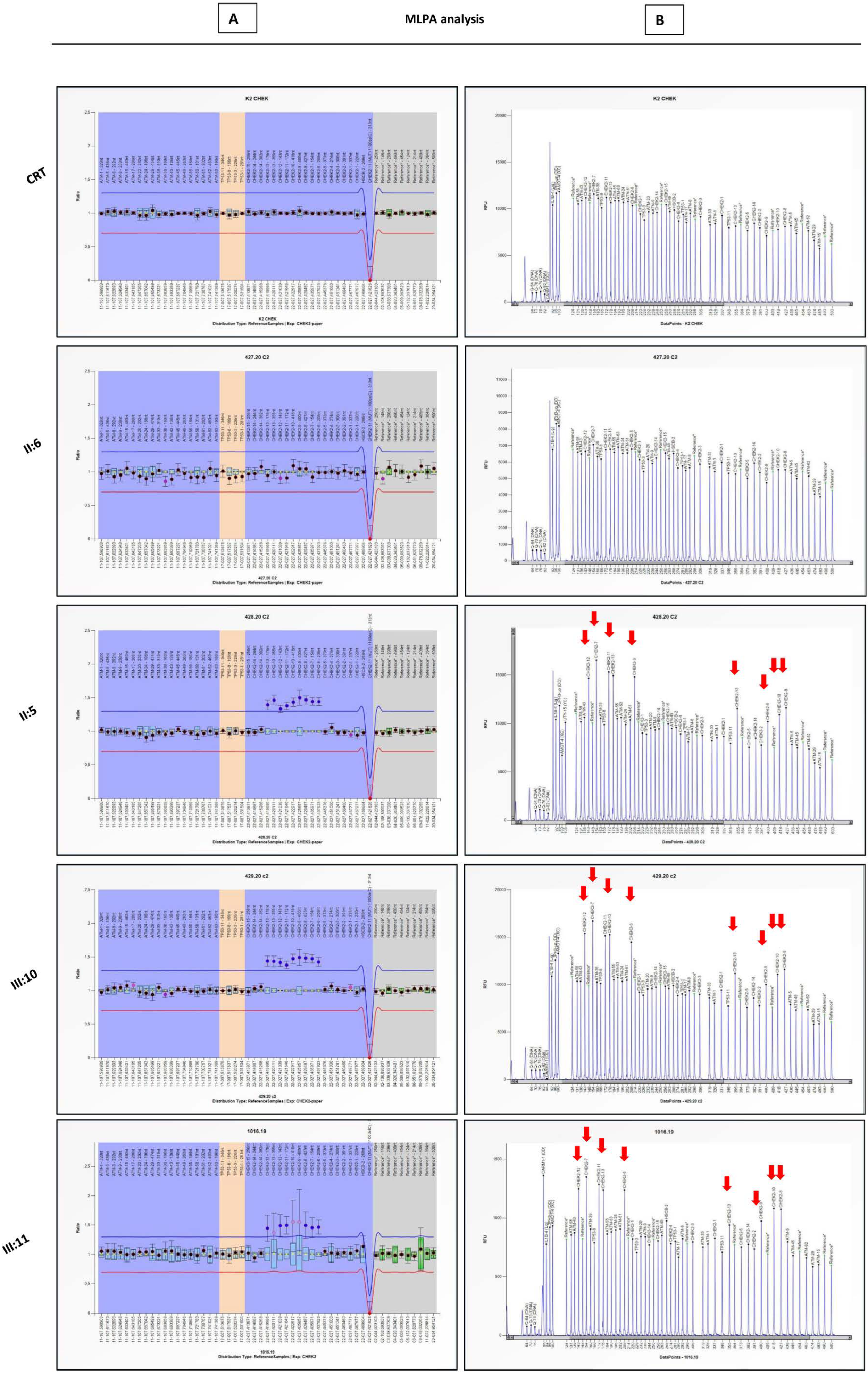
3.1. NGS and MLPA Results for BRCA1/2 Genes
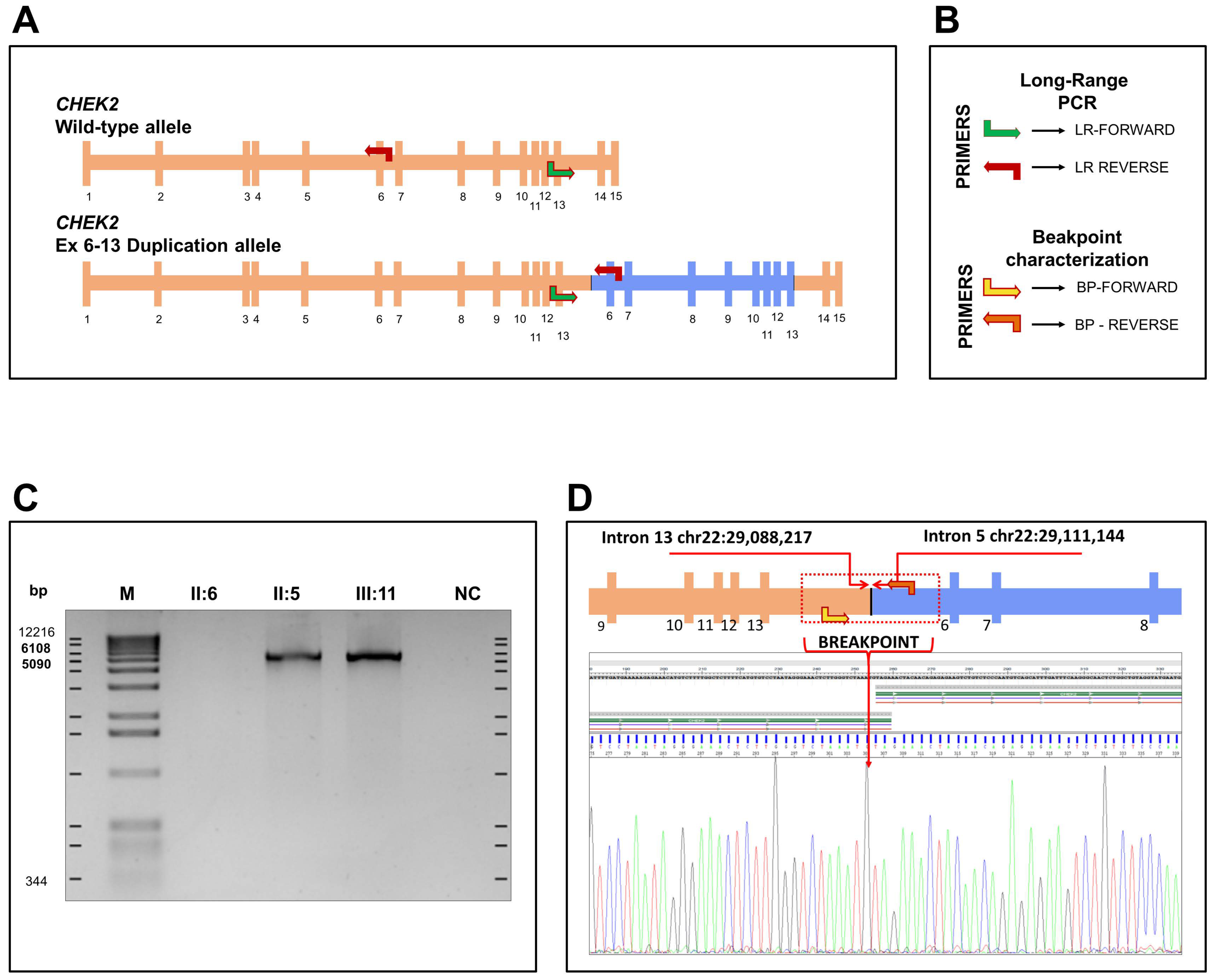
3.2. Large Rearrangement Characterization on CHEK2 Gene
3.3. Characterization and Identification of Breakpoint Duplication
3.4. RNA Analysis
4. Materials and Methods
4.1. Mutational Analysis of BRCA1 and BRCA2 Genes
4.1.1. NGS Analysis
4.1.2. MLPA Analysis
4.2. Duplication Analysis of CHEK2 Gene
4.3. Long-Range PCR and Breakpoint Analysis
4.4. RNA Extraction and qPCR
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pujol, P.; Barberis, M.; Beer, P.; Friedman, E.; Piulats, J.M.; Capoluongo, E.D.; Garcia Foncillas, J.; Ray-Coquard, I.; Penault-Llorca, F.; Foulkes, W.D.; et al. Clinical practice guidelines for BRCA1 and BRCA2 genetic testing. Eur. J. Cancer 2021, 146, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Germani, A.; Petrucci, S.; De Marchis, L.; Libi, F.; Savio, C.; Amanti, C.; Bonifacino, A.; Campanella, B.; Capalbo, C.; Lombardi, A.; et al. Beyond BRCA1 and BRCA2: Deleterious Variants in DNA Repair Pathway Genes in Italian Families with Breast/Ovarian and Pancreatic Cancers. J. Clin. Med. 2020, 9, 3003. [Google Scholar] [CrossRef] [PubMed]
- Stolarova, L.; Kleiblova, P.; Janatova, M.; Soukupova, J.; Zemankova, P.; Macurek, L.; Kleibl, Z. CHEK2 Germline Variants in Cancer Predisposition: Stalemate Rather than Checkmate. Cells 2020, 9, 2675. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Chehab, N.H.; Pavletich, N.P. Structure and activation mechanism of the CHK2 DNA damage checkpoint kinase. Mol. Cell 2020, 35, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, E.G.; Stettner, A.R.; Miller, S.A.; Solomon, S.R.; Marshall, M.L.; Roberts, M.E.; Susswein, L.R.; Arvai, K.J.; Klein, R.T.; Murphy, P.D.; et al. Differences in cancer prevalence among CHEK2 carriers identified via multi-gene panel testing. Cancer Genet. 2020, 246, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Tedaldi, G.; Danesi, R.; Zampiga, V.; Tebaldi, M.; Bedei, L.; Zoli, W.; Amadori, D.; Falcini, F.; Calistri, D. First evidence of a large CHEK2 duplication involved in cancer predisposition in an Italian family with hereditary breast cancer. BMC Cancer 2014, 14, 478. [Google Scholar] [CrossRef] [Green Version]
- Leedom, T.P.; LaDuca, H.; McFarland, R.; Li, S.; Dolinsky, J.S.; Chao, E.C. Breast cancer risk is similar for CHEK2 founder and non-founder mutation carriers. Cancer Genet. 2016, 209, 403–407. [Google Scholar] [CrossRef] [Green Version]
- Cybulski, C.; Wokołorczyk, D.; Huzarski, T.; Byrski, T.; Gronwald, J.; Górski, B.; Debniak, T.; Masojć, B.; Jakubowska, A.; Gliniewicz, B.; et al. A large germline deletion in the Chek2 kinase gene is associated with an increased risk of prostate cancer. J. Med. Genet. 2006, 43, 863–866. [Google Scholar] [CrossRef]
- Cybulski, C.; Wokołorczyk, D.; Huzarski, T.; Byrski, T.; Gronwald, J.; Górski, B.; Debniak, T.; Masojć, B.; Jakubowska, A.; van de Wetering, T.; et al. A deletion in CHEK2 of 5,395 bp predisposes to breast cancer in Poland. Breast Cancer Res. Treat. 2007, 102, 119–122. [Google Scholar] [CrossRef]
- Cybulski, C.; Górski, B.; Huzarski, T.; Masojć, B.; Mierzejewski, M.; Debniak, T.; Teodorczyk, U.; Byrski, T.; Gronwald, J.; Matyjasik, J.; et al. CHEK2 is a multiorgan cancer susceptibility gene. Am. J. Hum. Genet. 2014, 75, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2014, 17, 405–424. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Parra, G.; Del Valle, J.; Rofes, P.; Gausachs, M.; Stradella, A.; Moreno-Cabrera, J.M.; Velasco, A.; Tornero, E.; Menéndez, M.; Muñoz, X.; et al. Comprehensive analysis and ACMG-based classification of CHEK2 variants in hereditary cancer patients. Hum. Mut. 2020, 41, 2128–2142. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.E.; Chong, H.; Mu, W.; Conner, B.R.; Hsuan, V.; Willett, S.; Lam, S.; Tsai, P.; Pesaran, T.; Chamberlin, A.C.; et al. DNA breakpoint assay reveals a majority of gross duplications occur in tandem reducing VUS classifications in breast cancer predisposition genes. Genet. Med. 2020, 21, 683–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jekimovs, C.R.; Chen, X.; Arnold, J.; Gatei, M.; Richard, D.J.; Spurdle, A.B.; Khanna, K.K.; Chenevix-Trench, G. Low frequency of CHEK2 1100delC allele in Australian multiple-case breast cancer families: Functional analysis in heterozygous individuals. Br. J. Cancer 2005, 92, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiblová, P.; Stolařová, L.; Křížová, K.; Lhota, F.; Hojný, J.; Zemánková, P.; Havránek, O.; Vočka, M.; Černá, M.; Lhotová, K.; et al. Germline CHEK2 Gene Mutations in Hereditary Breast Cancer Predisposition—Mutation Types and Their Biological and Clinical Relevance. Dědičné mutace v genu CHEK2 jako příčina dispozice k nádorům prsu—Typy mutací, jejich biologická a klinická relevance. Klin. Onkol. 2020, 32 (Suppl. S2), 36–50. [Google Scholar] [CrossRef]
- Daly, M.B.; Pilarski, R.; Yurgelun, M.B.; Berry, M.P.; Buys, S.S.; Dickson, P.; Domchek, S.M.; Elkhanany, A.; Friedman, S.; Garber, J.E.; et al. NCCN Guidelines Insights: Genetic/Familial High-Risk Assessment: Breast, Ovarian, and Pancreatic, Version 1.2020. J. Natl. Compr. Cancer Netw. 2020, 18, 380–391. [Google Scholar] [CrossRef]
- Ow, G.S.; Ivshina, A.V.; Fuentes, G.; Kuznetsov, V.A. Identification of two poorly prognosed ovarian carcinoma subtypes associated with CHEK2 germ-line mutation and non-CHEK2 somatic mutation gene signatures. Cell Cycle 1999, 13, 2262–2280. [Google Scholar] [CrossRef] [Green Version]
- Bell, D.W.; Varley, J.M.; Szydlo, T.E.; Kang, D.H.; Wahrer, D.C.; Shannon, K.E.; Lubratovich, M.; Verselis, S.J.; Isselbacher, K.J.; Fraumeni, J.F.; et al. Heterozygous germ line hCHK2 mutations in Li-Fraumeni syndrome. Science 1999, 286, 2528–2531. [Google Scholar] [CrossRef]
- Sodha, N.; Houlston, R.S.; Bullock, S.; Yuille, M.A.; Chu, C.; Turner, G.; Eeles, R.A. Increasing evidence that germline mutations in CHEK2 do not cause Li-Fraumeni syndrome. Hum. Mut. 1999, 20, 460–462. [Google Scholar] [CrossRef]
- Evans, D.G.; Birch, J.M.; Narod, S.A. Is CHEK2 a cause of the Li-Fraumeni syndrome? J. Med. Genet. 1999, 45, 63–64. [Google Scholar] [CrossRef]
- De Silva, D.L.; Winship, I. Is CHEK2 a moderate-risk breast cancer gene or the younger sister of Li-Fraumeni? BMJ Case Rep. 2020, 13, e236435. [Google Scholar] [CrossRef] [PubMed]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.R.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Jang, W.; et al. ClinVar: Improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 2018, 46, D1062–D1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cybulski, C.; Wokołorczyk, D.; Jakubowska, A.; Huzarski, T.; Byrski, T.; Gronwald, J.; Masojć, B.; Deebniak, T.; Górski, B.; Blecharz, P.; et al. Risk of breast cancer in women with a CHEK2 mutation with and without a family history of breast cancer. J. Clin. Oncol. 2011, 29, 3747–3752. [Google Scholar] [CrossRef] [PubMed]
- Bąk, A.; Janiszewska, H.; Junkiert-Czarnecka, A.; Heise, M.; Pilarska-Deltow, M.; Laskowski, R.; Pasińska, M.; Haus, O. A risk of breast cancer in women—Carriers of constitutional CHEK2 gene mutations, originating from the North-Central Poland. Hered. Cancer Clin. Pract. 2014, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Lykke-Andersen, S.; Jensen, T.H. Nonsense-mediated mRNA decay: An intricate machinery that shapes transcriptomes. Mol. Cell Biol. 2015, 16, 665–677. [Google Scholar] [CrossRef] [Green Version]
- Van Luttikhuizen, J.L.; Bublitz, J.; Schubert, S.; Schmidt, G.; Hofmann, W.; Morlot, S.; Buurman, R.; Auber, B.; Schlegelberger, B.; Steinemann, D. From a variant of unknown significance to pathogenic: Reclassification of a large novel duplication in BRCA2 by high-throughput sequencing. Mol. Genet. Genom. Med. 2020, 8, e1045. [Google Scholar] [CrossRef] [Green Version]
- Szymanska-Pasternak, J.; Szymanska, A.; Medrek, K.; Imyanitov, E.N.; Cybulski, C.; Gorski, B.; Magnowski, P.; Dziuba, I.; Gugala, K.; Debniak, B.; et al. CHEK2 variants predispose to benign, borderline and low-grade invasive ovarian tumors. Gynecol. Oncol. 2006, 102, 429–431. [Google Scholar] [CrossRef]
- Baysal, B.E.; DeLoia, J.A.; Willett-Brozick, J.E.; Goodman, M.T.; Brady, M.F.; Modugno, F.; Lynch, H.T.; Conley, Y.P.; Watson, P.; Gallion, H.H. Analysis of CHEK2 gene for ovarian cancer susceptibility. Gynecol. Oncol. 2004, 95, 62–69. [Google Scholar] [CrossRef]
- Trost, B.; Walker, S.; Wang, Z.; Thiruvahindrapuram, B.; MacDonald, J.R.; Sung, W.; Pereira, S.L.; Whitney, J.; Chan, A.; Pellecchia, G.; et al. A Comprehensive Workflow for Read Depth-Based Identification of Copy-Number Variation from Whole-Genome Sequence Data. Am. J. Hum. Genet. 2018, 102, 142–155. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germani, A.; Guadagnolo, D.; Salvati, V.; Micolonghi, C.; Mancini, R.; Mastromoro, G.; Sadeghi, S.; Petrucci, S.; Pizzuti, A.; Piane, M. Genomic Breakpoints’ Characterization of a Large CHEK2 Duplication in an Italian Family with Hereditary Breast Cancer. Diagnostics 2022, 12, 1520. https://doi.org/10.3390/diagnostics12071520
Germani A, Guadagnolo D, Salvati V, Micolonghi C, Mancini R, Mastromoro G, Sadeghi S, Petrucci S, Pizzuti A, Piane M. Genomic Breakpoints’ Characterization of a Large CHEK2 Duplication in an Italian Family with Hereditary Breast Cancer. Diagnostics. 2022; 12(7):1520. https://doi.org/10.3390/diagnostics12071520
Chicago/Turabian StyleGermani, Aldo, Daniele Guadagnolo, Valentina Salvati, Caterina Micolonghi, Rita Mancini, Gioia Mastromoro, Soha Sadeghi, Simona Petrucci, Antonio Pizzuti, and Maria Piane. 2022. "Genomic Breakpoints’ Characterization of a Large CHEK2 Duplication in an Italian Family with Hereditary Breast Cancer" Diagnostics 12, no. 7: 1520. https://doi.org/10.3390/diagnostics12071520
APA StyleGermani, A., Guadagnolo, D., Salvati, V., Micolonghi, C., Mancini, R., Mastromoro, G., Sadeghi, S., Petrucci, S., Pizzuti, A., & Piane, M. (2022). Genomic Breakpoints’ Characterization of a Large CHEK2 Duplication in an Italian Family with Hereditary Breast Cancer. Diagnostics, 12(7), 1520. https://doi.org/10.3390/diagnostics12071520