
The Potential Role of Growth Differentiation Factor 15 in COVID-19: A Corollary Subjective Effect or Not?

 , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
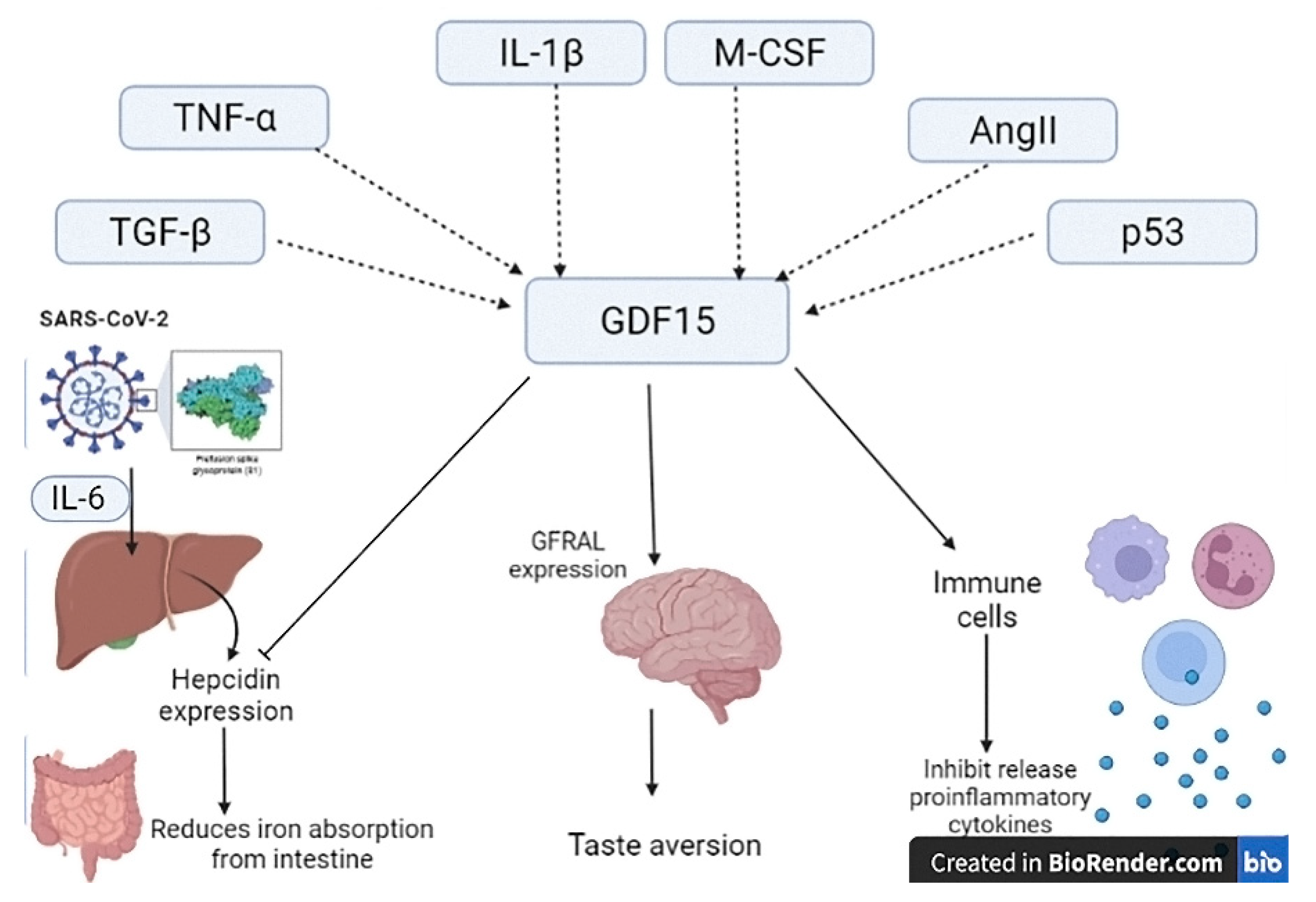
2. Growth Differentiation Factor 15
3. Growth Differentiation Factor 15 and Viral Infections
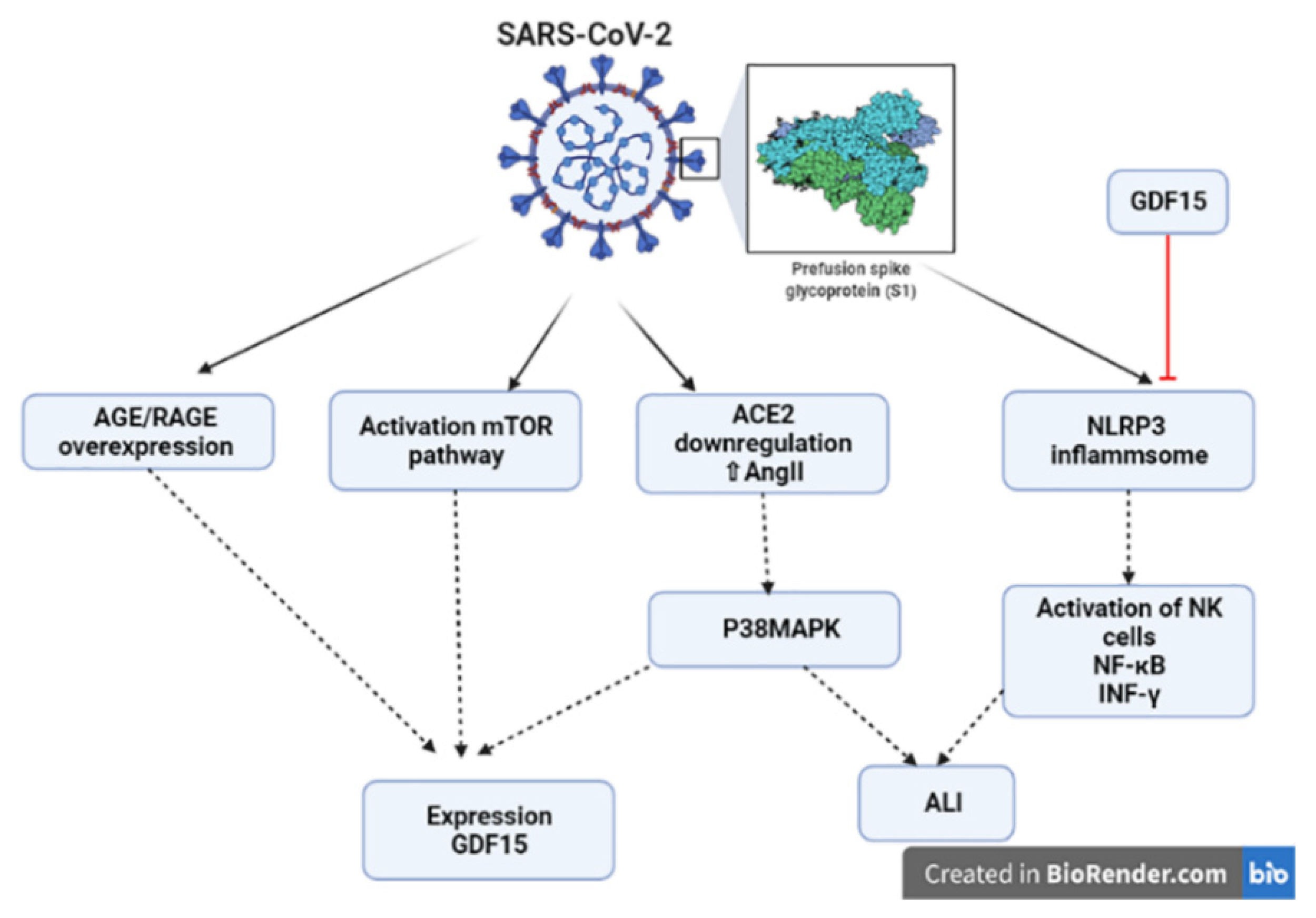
4. Growth Differentiation Factor 15 and COVID-19
5. Growth Differentiation Factor 15 and Inflammatory Pathways in COVID-19
6. GDF15 in Comparison with Other COVID-19 Biomarkers
7. Modulation Release of GDF15
8. Mitochondrial Dysfunction and GDF15 in COVID-19
9. Thrombosis and GDF15 in COVID-19
10. Gastrointestinal Injury and GDF15 in COVID-19
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Fageyinbo, M.S.; Batiha, G.E.-S. Vinpocetine is the forthcoming adjuvant agent in the management of COVID-19. Future Sci. OA 2022, 8, FSO797. [Google Scholar] [CrossRef] [PubMed]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Alzahrani, K.J.; Alexiou, A.; Batiha, G.E.-S. Niclosamide for COVID-19: Bridging the gap. Mol. Biol. Rep. 2021, 48, 8195–8202. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.; Moubarak, M.; Shaheen, H.M.; Zakariya, A.M.; Usman, I.M.; Rauf, A.; Adhikari, A.; Dey, A.; Alexiou, A.; Hetta, H.F. Favipiravir in SARS-CoV-2 infection: Is it Worthwhile? Comb. Chem. High Throughput Screen. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.; Muecksch, F.; Weisblum, Y.; Da Silva, J.; Bednarski, E.; Cho, A.; Wang, Z.; Gaebler, C.; Caskey, M.; Nussenzweig, M.C. Plasma neutralization of the SARS-CoV-2 Omicron variant. N. Engl. J. Med. 2022, 386, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Alorabi, M.; Mohammed, D.S.; Mostafa-Hedeab, G.; El-Sherbeni, S.A.; Negm, W.A.; Mohammed, A.I.A.; Al-Kuraishy, H.M.; Nasreldin, N.; Alotaibi, S.S.; Lawal, B. Combination treatment of omega-3 fatty acids and vitamin C exhibited promising therapeutic effect against oxidative impairment of the liver in methotrexate-intoxicated mice. Biomed. Res. Int. 2022, 2022, 4122166. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Qusti, S.; Alshammari, E.M.; Gyebi, G.A.; Batiha, G.E.-S. COVID-19-induced dysautonomia: A menace of sympathetic storm. ASN Neuro 2021, 13, 17590914211057635. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Alkazmi, L.; Habotta, O.A.; Batiha, G.E.-S. High-mobility group box 1 (HMGB1) in COVID-19: Extrapolation of dangerous liaisons. Inflammopharmacology 2022, 30, 811–820. [Google Scholar] [CrossRef]
- Papadopoulou, G.; Manoloudi, E.; Repousi, N.; Skoura, L.; Hurst, T.; Karamitros, T. Molecular and clinical prognostic biomarkers of COVID-19 severity and persistence. Pathogens 2022, 11, 311. [Google Scholar] [CrossRef]
- de Guadiana Romualdo, L.G.; Mulero, M.D.R.; Olivo, M.H.; Rojas, C.R.; Arenas, V.R.; Morales, M.G.; Abellán, A.B.; Conesa-Zamora, P.; García-García, J.; Hernández, A.C. Circulating levels of GDF-15 and calprotectin for prediction of in-hospital mortality in COVID-19 patients: A case series. J. Infect. 2021, 82, e40–e42. [Google Scholar] [CrossRef]
- Wollert, K.C.; Kempf, T.; Wallentin, L. Growth differentiation factor 15 as a biomarker in cardiovascular disease. Clin. Chem. 2017, 63, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Bootcov, M.R.; Bauskin, A.R.; Valenzuela, S.M.; Moore, A.G.; Bansal, M.; He, X.Y.; Zhang, H.P.; Donnellan, M.; Mahler, S.; Pryor, K. MIC-1, a novel macrophage inhibitory cytokine, is a divergent member of the TGF-β superfamily. Proc. Natl. Acad. Sci. USA 1997, 94, 11514–11519. [Google Scholar] [CrossRef] [PubMed]
- Corre, J.; Hébraud, B.; Bourin, P. Concise review: Growth differentiation factor 15 in pathology: A clinical role? Stem Cells Transl. Med. 2013, 2, 946–952. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef]
- Wu, Z.; Bennett, D.; Brosnan, J.; Calle, R.; Collins, S.; Esquejo, R.; Joaquim, S.; Joyce, A.; Kim, H.; LaCarubba, B. 2O growth differentiation factor 15 (GDF-15) neutralization reverses cancer cachexia, restores physical performance and mitigates emesis associated with platinum-based chemotherapy. Ann. Oncol. 2020, 31, S245. [Google Scholar] [CrossRef]
- Chang, J.Y.; Hong, H.J.; Kang, S.G.; Kim, J.T.; Zhang, B.Y.; Shong, M. The role of growth differentiation factor 15 in energy metabolism. Diabetes Metab. J. 2020, 44, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.C.; Trask, A.J. Growth differentiation factor-15 and its role in diabetes and cardiovascular disease. Cytokine Growth Factor Rev. 2021, 57, 11–18. [Google Scholar] [CrossRef]
- L’homme, L.; Sermikli, B.P.; Staels, B.; Piette, J.; Legrand-Poels, S.; Dombrowicz, D. Saturated fatty acids promote GDF15 expression in human macrophages through the PERK/eIF2/CHOP signaling pathway. Nutrients 2020, 12, 3771. [Google Scholar] [CrossRef]
- Berezin, A.E. Diabetes mellitus related biomarker: The predictive role of growth-differentiation factor-15. Diabetes Metab. Syndr. Clin. Res. Rev. 2016, 10, S154–S157. [Google Scholar] [CrossRef] [PubMed]
- George, M.; Jena, A.; Srivatsan, V.; Muthukumar, R.; Dhandapani, V. GDF 15-a novel biomarker in the offing for heart failure. Curr. Cardiol. Rev. 2016, 12, 37–46. [Google Scholar] [CrossRef]
- Tanrıkulu, O.; Sarıyıldız, M.A.; Batmaz, İ.; Yazmalar, L.; Polat, N.; Kaplan, İ.; Çevik, R. Serum GDF-15 level in rheumatoid arthritis: Relationship with disease activity and subclinical atherosclerosis. Acta Reumatol. Port. 2017, 1, 66–72. [Google Scholar]
- Coll, A.P.; Chen, M.; Taskar, P.; Rimmington, D.; Patel, S.; Tadross, J.A.; Cimino, I.; Yang, M.; Welsh, P.; Virtue, S. GDF15 mediates the effects of metformin on body weight and energy balance. Nature 2020, 578, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Insights into mechanisms of GDF15 and receptor GFRAL: Therapeutic targets. Trends Endocrinol. Metab. 2020, 31, 939–951. [Google Scholar] [CrossRef]
- Wu, Q.; Jiang, D.; Schaefer, N.R.; Harmacek, L.; O’Connor, B.P.; Eling, T.E.; Eickelberg, O.; Chu, H.W. Overproduction of growth differentiation factor 15 promotes human rhinovirus infection and virus-induced inflammation in the lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L514–L527. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, P.; Librán-Pérez, M.; Figueras, A.; Novoa, B. Conserved function of zebrafish (Danio rerio) Gdf15 as a sepsis tolerance mediator. Dev. Comp. Immunol. 2020, 109, 103698. [Google Scholar] [CrossRef]
- Kriechbaum, S.D.; Wiedenroth, C.B.; Peters, K.; Barde, M.A.; Ajnwojner, R.; Wolter, J.-S.; Haas, M.; Roller, F.C.; Guth, S.; Rieth, A.J. Galectin-3, GDF-15, and sST2 for the assessment of disease severity and therapy response in patients suffering from inoperable chronic thromboembolic pulmonary hypertension. Biomarkers 2020, 25, 578–586. [Google Scholar] [CrossRef]
- Lignelli, E.; Palumbo, F.; Myti, D.; Morty, R.E. Recent advances in our understanding of the mechanisms of lung alveolarization and bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L832–L887. [Google Scholar] [CrossRef]
- Si, Y.; Liu, X.; Cheng, M.; Wang, M.; Gong, Q.; Yang, Y.; Wang, T.; Yang, W. Growth differentiation factor 15 is induced by hepatitis C virus infection and regulates hepatocellular carcinoma-related genes. PLoS ONE 2011, 6, e19967. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dai, W.; Zhu, C.; Liu, H.; Li, Y.; Zhang, P. Circulating levels of growth differentiation factor 15 and sex hormones in male patients with HBV-associated hepatocellular carcinoma. Biomed. Pharmacother. 2020, 121, 109574. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Sjøberg, K.A.; Carl, C.S.; Jeppesen, J.F.; Wojtaszewski, J.F.; Kiens, B.; Richter, E.A. Exercise increases circulating GDF15 in humans. Mol. Metab. 2018, 9, 187–191. [Google Scholar] [CrossRef]
- Zhao, B.; Li, H.; Cao, S.; Zhong, W.; Li, B.; Jia, W.; Ning, Z. Negative regulators of inflammation response to the dynamic expression of cytokines in DF-1 and MDCK cells infected by avian influenza viruses. Inflammation 2022, 45, 573–589. [Google Scholar] [CrossRef]
- Luan, H.H.; Wang, A.; Hilliard, B.K.; Carvalho, F.; Rosen, C.E.; Ahasic, A.M.; Herzog, E.L.; Kang, I.; Pisani, M.A.; Yu, S. GDF15 is an inflammation-induced central mediator of tissue tolerance. Cell 2019, 178, 1231–1244.e1211. [Google Scholar] [CrossRef]
- Ahmed, D.S.; Isnard, S.; Berini, C.; Lin, J.; Routy, J.-P.; Royston, L. Coping with stress: The mitokine GDF-15 as a biomarker of COVID-19 severity. Front. Immunol. 2022, 13, 820350. [Google Scholar] [CrossRef]
- Teng, X.; Zhang, J.; Shi, Y.; Liu, Y.; Yang, Y.; He, J.; Luo, S.; Huang, Y.; Liu, Y.; Liu, D. Comprehensive profiling of inflammatory factors revealed that growth differentiation factor-15 is an indicator of disease severity in COVID-19 patients. Front. Immunol. 2021, 12, 662465. [Google Scholar] [CrossRef]
- Sindhu, R.K.; Kaur, P.; Kaur, P.; Singh, H.; Batiha, G.E.-S.; Verma, I. Exploring multifunctional antioxidants as potential agents for management of neurological disorders. Environ. Sci. Pollut. Res. 2022, 29, 24458–24477. [Google Scholar] [CrossRef]
- Mostafa-Hedeab, G.; Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Jeandet, P.; Saad, H.M.; Batiha, G.E.-S. A raising dawn of pentoxifylline in management of inflammatory disorders in COVID-19. Inflammopharmacology 2022, 30, 799–809. [Google Scholar] [CrossRef]
- Lippi, G.; Henry, B.M. Prognostic value of growth differentiation factor 15 in COVID-19. Scand. J. Clin. Lab. Investig. 2022, 82, 170–172. [Google Scholar] [CrossRef]
- Notz, Q.; Schmalzing, M.; Wedekink, F.; Schlesinger, T.; Gernert, M.; Herrmann, J.; Sorger, L.; Weismann, D.; Schmid, B.; Sitter, M. Pro-and anti-inflammatory responses in severe COVID-19-induced acute respiratory distress syndrome—An observational pilot study. Front. Immunol. 2020, 11, 581338. [Google Scholar] [CrossRef]
- Gisby, J.; Clarke, C.L.; Medjeral-Thomas, N.; Malik, T.H.; Papadaki, A.; Mortimer, P.M.; Buang, N.B.; Lewis, S.; Pereira, M.; Toulza, F. Longitudinal proteomic profiling of dialysis patients with COVID-19 reveals markers of severity and predictors of death. Elife 2021, 10, e64827. [Google Scholar] [CrossRef]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. GDF15: An emerging modulator of immunity and a strategy in COVID-19 in association with iron metabolism. Trends Endocrinol. Metab. 2021, 32, 875–889. [Google Scholar] [CrossRef]
- Chung, H.K.; Kim, J.T.; Kim, H.-W.; Kwon, M.; Kim, S.Y.; Shong, M.; Kim, K.S.; Yi, H.-S. GDF15 deficiency exacerbates chronic alcohol-and carbon tetrachloride-induced liver injury. Sci. Rep. 2017, 7, 17238. [Google Scholar] [CrossRef]
- Song, H.; Chen, Q.; Xie, S.; Huang, J.; Kang, G. GDF-15 prevents lipopolysaccharide-mediated acute lung injury via upregulating SIRT1. Biochem. Biophys. Res. Commun. 2020, 526, 439–446. [Google Scholar] [CrossRef]
- Banaszkiewicz, M.; Małyszko, J.; Vesole, D.H.; Woziwodzka, K.; Jurczyszyn, A.; Żórawski, M.; Krzanowski, M.; Małyszko, J.; Batko, K.; Kuźniewski, M. New biomarkers of ferric management in multiple myeloma and kidney disease-associated anemia. J. Clin. Med. 2019, 8, 1828. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Al-Gareeb, A.I.; Qusti, S.; Alshammari, E.M.; Kaushik, D.; Verma, R.; Al-Kuraishy, H.M. Deciphering the immunoboosting potential of macro and micronutrients in COVID support therapy. Environ. Sci. Pollut. Res. 2022, 29, 43516–43531. [Google Scholar] [CrossRef]
- Nai, A.; Lorè, N.I.; Pagani, A.; De Lorenzo, R.; Di Modica, S.; Saliu, F.; Cirillo, D.M.; Rovere-Querini, P.; Manfredi, A.A.; Silvestri, L. Hepcidin levels predict COVID-19 severity and mortality in a cohort of hospitalized Italian patients. Am. J. Hematol. 2021, 96, E32–E35. [Google Scholar] [CrossRef]
- Mostafa-Hedeab, G.; Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Welson, N.N.; Batiha, G.E.-S.; Conte-Junior, C.A. Selinexor and COVID-19: The neglected warden. Front. Pharmacol. 2022, 13, 884228. [Google Scholar] [CrossRef]
- Ehsani, S. Distant sequence similarity between hepcidin and the novel coronavirus spike glycoprotein: A potential hint at the possibility of local iron dysregulation in COVID-19. arXiv 2020, arXiv:2003.12191. [Google Scholar]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Welson, N.N.; Batiha, G.E.-S. Trimetazidine and COVID-19-induced acute cardiac injury: A missed key. Int. J. Clin. Pharm. 2022, 44, 832–833. [Google Scholar] [CrossRef]
- Zeng, J.; Xie, X.; Feng, X.-L.; Xu, L.; Han, J.-B.; Yu, D.; Zou, Q.-C.; Liu, Q.; Li, X.; Ma, G. Specific inhibition of the NLRP3 inflammasome suppresses immune overactivation and alleviates COVID-19 like pathology in mice. EBioMedicine 2022, 75, 103803. [Google Scholar] [CrossRef]
- Lertpatipanpong, P.; Lee, J.; Kim, I.; Eling, T.; Oh, S.Y.; Seong, J.K.; Baek, S.J. The anti-diabetic effects of NAG-1/GDF15 on HFD/STZ-induced mice. Sci. Rep. 2021, 11, 15027. [Google Scholar] [CrossRef]
- Roberti, A.; Chaffey, L.E.; Greaves, D.R. NF-κB signaling and inflammation—Drug repurposing to treat inflammatory disorders? Biology 2022, 11, 372. [Google Scholar] [CrossRef]
- Ratnam, N.M.; Peterson, J.M.; Talbert, E.E.; Ladner, K.J.; Rajasekera, P.V.; Schmidt, C.R.; Dillhoff, M.E.; Swanson, B.J.; Haverick, E.; Kladney, R.D. NF-κB regulates GDF-15 to suppress macrophage surveillance during early tumor development. J. Clin. Investig. 2017, 127, 3796–3809. [Google Scholar] [CrossRef] [PubMed]
- Asiedu, S.O.; Kwofie, S.K.; Broni, E.; Wilson, M.D. Computational identification of potential anti-inflammatory natural compounds targeting the p38 mitogen-activated protein kinase (MAPK): Implications for COVID-19-induced cytokine storm. Biomolecules 2021, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Shirai, T.; Morishita, S.; Uchida, S.; Saeki-Miura, K.; Makishima, F. p38 mitogen-activated protein kinase functionally contributes to chondrogenesis induced by growth/differentiation factor-5 in ATDC5 cells. Exp. Cell Res. 1999, 250, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Ciullo, A.; Li, C.; Garcia, G.; Peck, K.; Miyamoto, K.; Arumugaswami, V.; Marbán, E. Engineered extracellular vesicles antagonize SARS-CoV-2 infection by inhibiting mTOR signaling. Biomater. Biosyst. 2022, 6, 100042. [Google Scholar] [CrossRef]
- Li, F.; Li, J.; Wang, P.-H.; Yang, N.; Huang, J.; Ou, J.; Xu, T.; Zhao, X.; Liu, T.; Huang, X. SARS-CoV-2 spike promotes inflammation and apoptosis through autophagy by ROS-suppressed PI3K/AKT/mTOR signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166260. [Google Scholar] [CrossRef]
- Wischhusen, J.; Melero, I.; Fridman, W.H. Growth/differentiation factor-15 (GDF-15): From biomarker to novel targetable immune checkpoint. Front. Immunol. 2020, 11, 951. [Google Scholar] [CrossRef]
- Sellegounder, D.; Zafari, P.; Rajabinejad, M.; Taghadosi, M.; Kapahi, P. Advanced glycation end products (AGEs) and its receptor, RAGE, modulate age-dependent COVID-19 morbidity and mortality. A review and hypothesis. Int. Immunopharmacol. 2021, 98, 107806. [Google Scholar] [CrossRef]
- Mouanness, M.; Merhi, Z. Impact of dietary advanced glycation end products on female reproduction: Review of potential mechanistic pathways. Nutrients 2022, 14, 966. [Google Scholar] [CrossRef]
- Papadopoulou-Marketou, N.; Chrousos, G.P.; Kanaka-Gantenbein, C. Diabetic nephropathy in type 1 diabetes: A review of early natural history, pathogenesis, and diagnosis. Diabetes Metab. Res. Rev. 2017, 33, e2841. [Google Scholar] [CrossRef]
- Cheema, P.S.; Nandi, D.; Nag, A. Exploring the therapeutic potential of forkhead box O for outfoxing COVID-19. Open Biol. 2021, 11, 210069. [Google Scholar] [CrossRef]
- Tsiambas, E.; Chrysovergis, A.; Papanikolaou, V.; Mastronikolis, N.; Ragos, V.; Kavantzas, N.; Lazaris, A.C.; Patsouris, E.; Riziotis, C.; Paschopoulos, M. Chromosome X riddle in SARS-CoV-2 (COVID-19)-related lung pathology. Pathol. Oncol. Res. 2020, 26, 2839–2841. [Google Scholar] [CrossRef] [PubMed]
- Kianmehr, A.; Faraoni, I.; Kucuk, O.; Mahrooz, A. Epigenetic alterations and genetic variations of angiotensin-converting enzyme 2 (ACE2) as a functional receptor for SARS-CoV-2: Potential clinical implications. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1587–1598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Moszczynski, L.A.; Liu, Q.; Jiang, J.; Zhao, D.; Quan, D.; Mele, T.; McAlister, V.; Jevnikar, A.; Baek, S.J. Over-expression of growth differentiation factor 15 (GDF15) preventing cold ischemia reperfusion (I/R) injury in heart transplantation through Foxo3a signaling. Oncotarget 2017, 8, 36531. [Google Scholar] [CrossRef] [PubMed]
- Albini, A.; Di Guardo, G.; Noonan, D.M.; Lombardo, M. The SARS-CoV-2 receptor, ACE-2, is expressed on many different cell types: Implications for ACE-inhibitor-and angiotensin II receptor blocker-based cardiovascular therapies. Intern. Emerg. Med. 2020, 15, 759–766. [Google Scholar] [CrossRef]
- Serebrovska, Z.O.; Chong, E.Y.; Serebrovska, T.V.; Tumanovska, L.V.; Xi, L. Hypoxia, HIF-1α, and COVID-19: From pathogenic factors to potential therapeutic targets. Acta Pharmacol. Sin. 2020, 41, 1539–1546. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, Y.; Guo, T.; Liu, F.; Xu, Y.; Cai, S. Hypoxia induces growth differentiation factor 15 to promote the metastasis of colorectal cancer via PERK-eIF2α signaling. Biomed. Res. Int. 2020, 2020, 5958272. [Google Scholar] [CrossRef]
- Giron, L.B.; Dweep, H.; Yin, X.; Wang, H.; Damra, M.; Goldman, A.R.; Gorman, N.; Palmer, C.S.; Tang, H.-Y.; Shaikh, M.W. Severe COVID-19 is fueled by disrupted gut barrier integrity. arXiv 2020, arXiv:20231209. [Google Scholar]
- Myhre, P.L.; Prebensen, C.; Strand, H.; Røysland, R.; Jonassen, C.M.; Rangberg, A.; Sørensen, V.; Søvik, S.; Røsjø, H.; Svensson, M. Growth differentiation factor 15 provides prognostic information superior to established cardiovascular and inflammatory biomarkers in unselected patients hospitalized with COVID-19. Circulation 2020, 142, 2128–2137. [Google Scholar] [CrossRef]
- Day, E.A.; Ford, R.J.; Smith, B.K.; Mohammadi-Shemirani, P.; Morrow, M.R.; Gutgesell, R.M.; Lu, R.; Raphenya, A.R.; Kabiri, M.; McArthur, A.G. Metformin-induced increases in GDF15 are important for suppressing appetite and promoting weight loss. Nat. Metab. 2019, 1, 1202–1208. [Google Scholar] [CrossRef]
- Scheen, A. Metformin and COVID-19: From cellular mechanisms to reduced mortality. Diabetes Metab. 2020, 46, 423–426. [Google Scholar] [CrossRef]
- Da Silveira, M.P.; da Silva Fagundes, K.K.; Bizuti, M.R.; Starck, É.; Rossi, R.C.; de Resende E Silva, D.T. Physical exercise as a tool to help the immune system against COVID-19: An integrative review of the current literature. Clin. Exp. Med. 2021, 21, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, A.; Vecchié, A.; Dagna, L.; Tangianu, F.; Abbate, A.; Dentali, F. Colchicine for COVID-19: Targeting NLRP3 inflammasome to blunt hyperinflammation. Inflamm. Res. 2022, 71, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Qusty, N.; Cruz-Martins, N.; Batiha, G.E.-S. Sequential doxycycline and colchicine combination therapy in COVID-19: The salutary effects. Pulm. Pharmacol. Ther. 2021, 67, 102008. [Google Scholar] [CrossRef]
- Al, H.M.; Hussien, N.R.; Al, M.S.; Al, A.I. Narrative review colchicine in the management of COVID-19: With or lieu of evidence. Narrative 2021, 71, 12. [Google Scholar]
- Yasmin, F.; Najeeb, H.; Moeed, A.; Hassan, W.; Khatri, M.; Asghar, M.S.; Naveed, A.K.; Ullah, W.; Surani, S. Safety and efficacy of colchicine in COVID-19 patients: A systematic review and meta-analysis of randomized control trials. PLoS ONE 2022, 17, e0266245. [Google Scholar] [CrossRef]
- Toro-Huamanchumo, C.J.; Benites-Meza, J.K.; Mamani-García, C.S.; Bustamante-Paytan, D.; Gracia-Ramos, A.E.; Diaz-Vélez, C.; Barboza, J.J. Efficacy of colchicine in the treatment of COVID-19 patients: A systematic review and meta-analysis. J. Clin. Med. 2022, 11, 2615. [Google Scholar] [CrossRef]
- Fujita, Y.; Ito, M.; Ohsawa, I. Mitochondrial stress and GDF15 in the pathophysiology of sepsis. Arch. Biochem. Biophys. 2020, 696, 108668. [Google Scholar] [CrossRef]
- Johann, K.; Kleinert, M.; Klaus, S. The role of GDF15 as a myomitokine. Cells 2021, 10, 2990. [Google Scholar] [CrossRef]
- Montero, R.; Yubero, D.; Villarroya, J.; Henares, D.; Jou, C.; Rodríguez, M.A.; Ramos, F.; Nascimento, A.; Ortez, C.I.; Campistol, J.; et al. GDF-15 is elevated in children with mitochondrial diseases and is induced by mitochondrial dysfunction. PLoS ONE 2016, 11, e0148709. [Google Scholar] [CrossRef]
- de Las Heras, N.; Martín Giménez, V.M.; Ferder, L.; Manucha, W.; Lahera, V. Implications of oxidative stress and potential role of mitochondrial dysfunction in COVID-19: Therapeutic effects of vitamin D. Antioxidants 2020, 9, 897. [Google Scholar] [CrossRef]
- Díaz-Resendiz, K.J.; Covantes-Rosales, C.E.; Benítez-Trinidad, A.B.; Navidad-Murrieta, M.S.; Razura-Carmona, F.F.; Carrillo-Cruz, C.D.; Frias-Delgadillo, E.J.; Pérez-Díaz, D.A.; Díaz-Benavides, M.V.; Zambrano-Soria, M.; et al. Effect of fucoidan on the mitochondrial membrane potential (ΔΨm) of leukocytes from patients with active COVID-19 and subjects that recovered from SARS-CoV-2 infection. Mar. Drugs 2022, 20, 99. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz-Enríquez, J.; Rojas-Morales, E.; Ruíz-García, M.G.; Tobón-Velasco, J.C.; Jiménez-Ortega, J.C. SARS-CoV-2 induces mitochondrial dysfunction and cell death by oxidative stress/inflammation in leukocytes of COVID-19 patients. Free Radic. Res. 2021, 55, 982–995. [Google Scholar] [CrossRef]
- Skorupski, W.J.; Grygier, M.; Lesiak, M.; Kałużna-Oleksy, M. Coronary stent thrombosis in COVID-19 patients: A systematic review of cases reported worldwide. Viruses 2022, 14, 260. [Google Scholar] [CrossRef] [PubMed]
- de Gregorio, C.; Colarusso, L.; Calcaterra, G.; Bassareo, P.P.; Ieni, A.; Mazzeo, A.T.; Ferrazzo, G.; Noto, A.; Koniari, I.; Mehta, J.L.; et al. Cerebral venous sinus thrombosis following COVID-19 vaccination: Analysis of 552 worldwide cases. Vaccines 2022, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.S.; Huisman, M.V.; Klok, F.A. Diagnostic management of acute pulmonary embolism in COVID-19 and other special patient populations. Diagnostics 2022, 12, 1350. [Google Scholar] [CrossRef]
- Mosleh, W.; Chen, K.; Pfau, S.E.; Vashist, A. Endotheliitis and endothelial dysfunction in patients with COVID-19: Its role in thrombosis and adverse outcomes. J. Clin. Med. 2020, 9, 1862. [Google Scholar] [CrossRef]
- Chi, G.; Lee, J.J.; Jamil, A.; Gunnam, V.; Najafi, H.; Memar Montazerin, S.; Shojaei, F.; Marszalek, J. Venous thromboembolism among hospitalized patients with COVID-19 undergoing thromboprophylaxis: A systematic review and meta-analysis. J. Clin. Med. 2020, 9, 2489. [Google Scholar] [CrossRef]
- Lankeit, M.; Kempf, T.; Dellas, C.; Cuny, M.; Tapken, H.; Peter, T.; Olschewski, M.; Konstantinides, S.; Wollert, K.C. Growth differentiation factor-15 for prognostic assessment of patients with acute pulmonary embolism. Am. J. Respir. Crit. Care Med. 2008, 177, 1018–1025. [Google Scholar] [CrossRef]
- Hu, X.F.; Zhan, R.; Xu, S.; Wang, J.; Wu, J.; Liu, X.; Li, Y.; Chen, L. Growth differentiation factor 15 is associated with left atrial/left atrial appendage thrombus in patients with nonvalvular atrial fibrillation. Clin. Cardiol. 2018, 41, 34–38. [Google Scholar] [CrossRef]
- Kaski, J.C. Inflammation and thrombosis in atrial fibrillation. Rev. Esp. Cardiol. 2011, 64, 551–553. [Google Scholar] [CrossRef]
- Rossaint, J.; Vestweber, D.; Zarbock, A. GDF-15 prevents platelet integrin activation and thrombus formation. J. Thromb. Haemost. 2013, 11, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Yadav, K.C.; Bhattarai, S.; Shiwakoti, L.D.; Paudel, S.; Subedi, M.; Pant, B.R.; Paudel, M.; Dhugana, S.; Bhattarai, S.; Tiwari, T. Sensorial and chemical analysis of biscuits prepared by incorporating Moringa flower powder and leaf powder. Int. J. Food Prop. 2022, 25, 894–906. [Google Scholar]
- Suryavanshi, S.V.; Zaiachuk, M.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids alleviate the LPS-induced cytokine storm via attenuating NLRP3 inflammasome signaling and TYK2-mediated STAT3 signaling pathways in vitro. Cells 2022, 11, 1391. [Google Scholar] [CrossRef] [PubMed]
- Zimmers, T.A.; Jin, X.; Hsiao, E.C.; Perez, E.A.; Pierce, R.H.; Chavin, K.D.; Koniaris, L.G. Growth differentiation factor-15: Induction in liver injury through p53 and tumor necrosis factor-independent mechanisms1. J. Surg. Res. 2006, 130, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Koniaris, L.G. Induction of MIC-1/growth differentiation factor-15 following bile duct injury. J. Gastrointest. Surg. 2003, 7, 901–905. [Google Scholar] [CrossRef]
- Lee, E.S.; Kim, S.H.; Kim, H.J.; Kim, K.H.; Lee, B.S.; Ku, B.J. Growth differentiation factor 15 predicts chronic liver disease severity. Gut Liver 2017, 11, 276. [Google Scholar] [CrossRef]
- Zhong, P.; Xu, J.; Yang, D.; Shen, Y.; Wang, L.; Feng, Y.; Du, C.; Song, Y.; Wu, C.; Hu, X. COVID-19-associated gastrointestinal and liver injury: Clinical features and potential mechanisms. Signal Transduct. Target Ther. 2020, 5, 256. [Google Scholar] [CrossRef]
- Huang, W.; Li, C.; Wang, Z.; Wang, H.; Zhou, N.; Jiang, J.; Ni, L.; Zhang, X.A.; Wang, D.-W. Decreased serum albumin level indicates poor prognosis of COVID-19 patients: Hepatic injury analysis from 2623 hospitalized cases. Sci. China Life Sci. 2020, 63, 1678–1687. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ref. | Study Type | Findings |
|---|---|---|
| de Guadiana et al. [9] | Prospective study | Higher GDF15 serum level was associated with higher mortality |
| Ahmed et al. [32] | Review study | Higher GDF15 serum level was regarded as a prognostic biomarker and correlated with COVID-19 severity. |
| Teng et al. [33] | Retrospective study | Higher GDF15 serum is an indicator of the COVID-19 severity. |
| Lippi and Henry [36] | Pooled analysis study | The GDF15 serum was significantly correlated with most of COVID-19 regardless of its severity. |
| Notz et al. [37] | Observational pilot study | The GDF15 is implicated in the pathogenesis and severity of COVID-19. |
| Gisby et al. [38] | Longitudinal proteomic study | The GDF15 serum level is correlated with COVID-19 severity |
| Rochette et al. [39] | Review study | The GDF15 can attenuate abnormal immune responses and prevent the associated inflammation in COVID-19. |
| Giron et al. [67] | A case–control study | The GDF15 serum level was increased together with increasing levels of galectin-9 and C3a in severely affected COVID-19 patients. |
| Myhre et al. [68] | A prospective observational study | The GDF15 offers a prognostic biomarker superior to other inflammatory biomarkers in unselected hospitalized COVID-19 patients. |
| Notz et al. [37] | Observational pilot study | In severely affected COVID-19 patients with ARDS at ICU, the anti-inflammatory IL-10 and GDF15 were increased, positively and negatively correlated with pro-inflammatory IL-6 and lymphopenia, respectively. |
| Huang et al. [98] | A retrospective study | The High GDF15 serum level is correlated with COVID-19-induced acute hepatic injury |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babalghith, A.O.; Al-kuraishy, H.M.; Al-Gareeb, A.I.; De Waard, M.; Sabatier, J.-M.; Saad, H.M.; Batiha, G.E.-S. The Potential Role of Growth Differentiation Factor 15 in COVID-19: A Corollary Subjective Effect or Not? Diagnostics 2022, 12, 2051. https://doi.org/10.3390/diagnostics12092051
Babalghith AO, Al-kuraishy HM, Al-Gareeb AI, De Waard M, Sabatier J-M, Saad HM, Batiha GE-S. The Potential Role of Growth Differentiation Factor 15 in COVID-19: A Corollary Subjective Effect or Not? Diagnostics. 2022; 12(9):2051. https://doi.org/10.3390/diagnostics12092051
Chicago/Turabian StyleBabalghith, Ahmad O., Hayder M. Al-kuraishy, Ali I. Al-Gareeb, Michel De Waard, Jean-Marc Sabatier, Hebatallah M. Saad, and Gaber El-Saber Batiha. 2022. "The Potential Role of Growth Differentiation Factor 15 in COVID-19: A Corollary Subjective Effect or Not?" Diagnostics 12, no. 9: 2051. https://doi.org/10.3390/diagnostics12092051
APA StyleBabalghith, A. O., Al-kuraishy, H. M., Al-Gareeb, A. I., De Waard, M., Sabatier, J. -M., Saad, H. M., & Batiha, G. E. -S. (2022). The Potential Role of Growth Differentiation Factor 15 in COVID-19: A Corollary Subjective Effect or Not? Diagnostics, 12(9), 2051. https://doi.org/10.3390/diagnostics12092051