
Use of Envelope Domain III Protein for the Detection of IgG Type Antibodies Specific to Zika Virus by Indirect ELISA
 ,
,  ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Descriptions
2.3. Study Design
2.4. Recombinant EDIII from ZIKV
2.5. Cells and Viruses
2.6. Antigens and Antibodies
2.7. Serologic Tests
- Inhouse Indirect IgG ELISA
- Plaque reduction neutralization test (PRNT)
- (a)
- PRNT testing for ZIKV, WNV, and YFV
- (b)
- FRNT testing for DENV
- (c)
- Recombinant Domain III indirect IgG ELISA
2.8. Statistical Analysis
3. Results
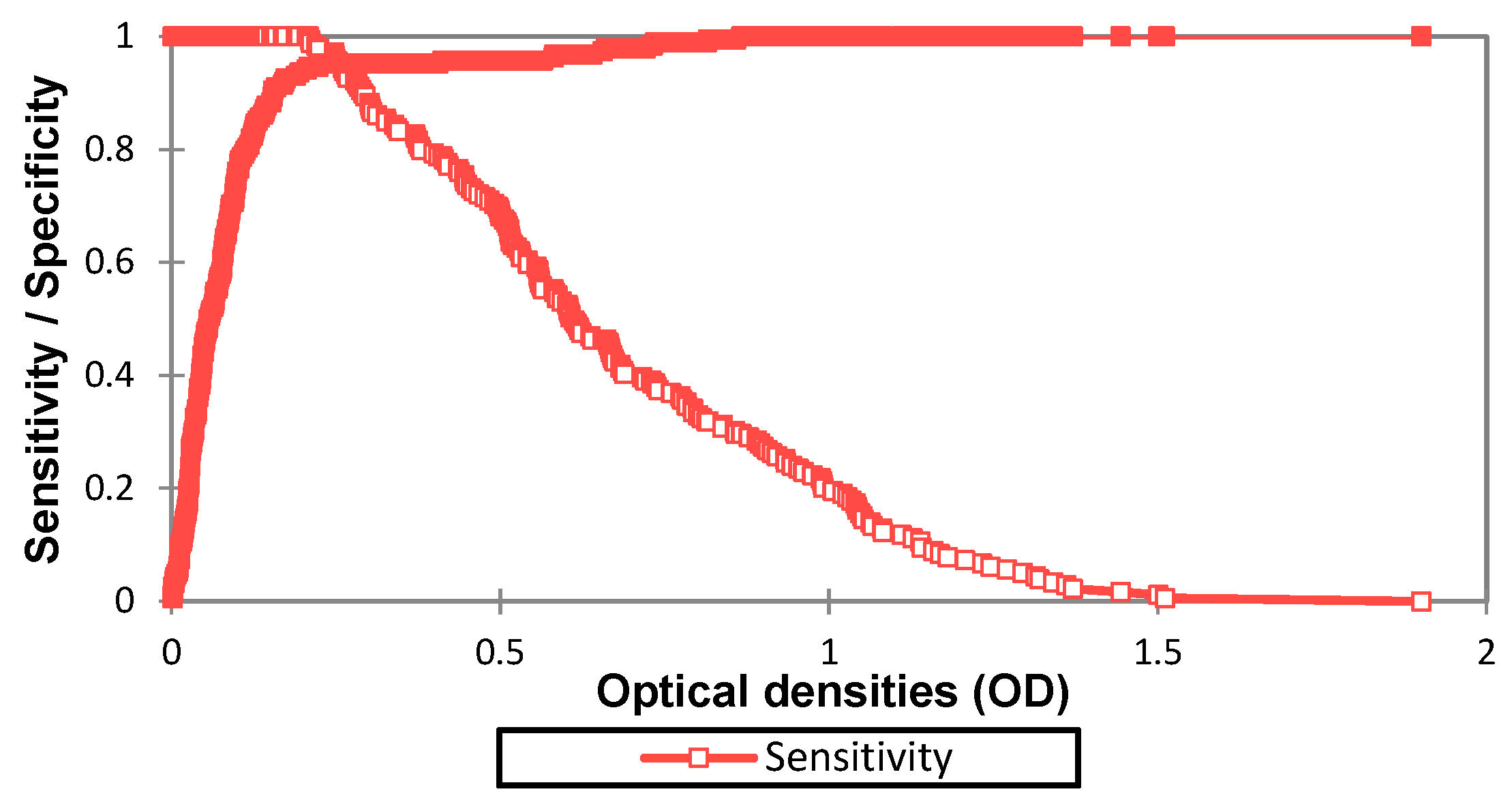
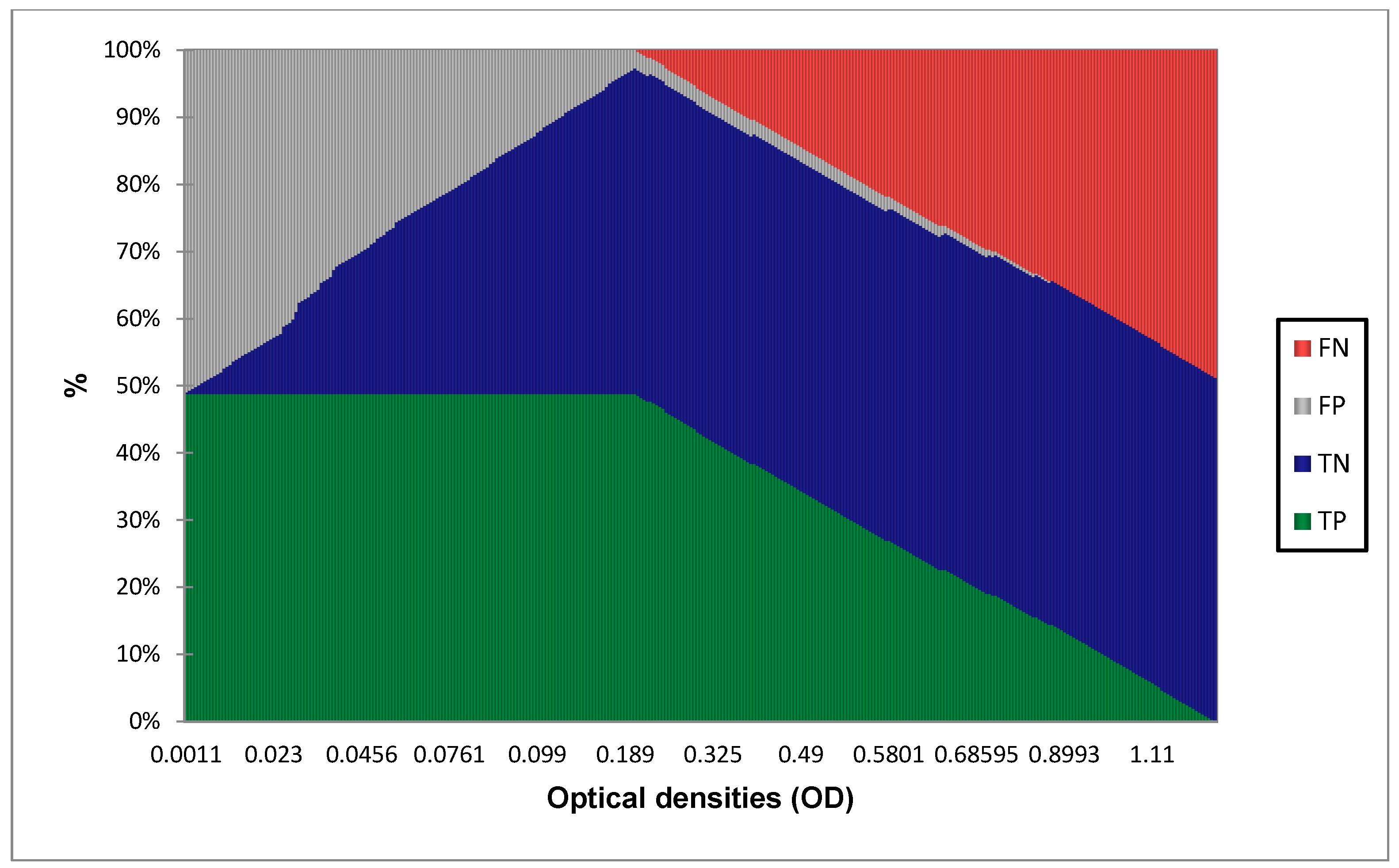
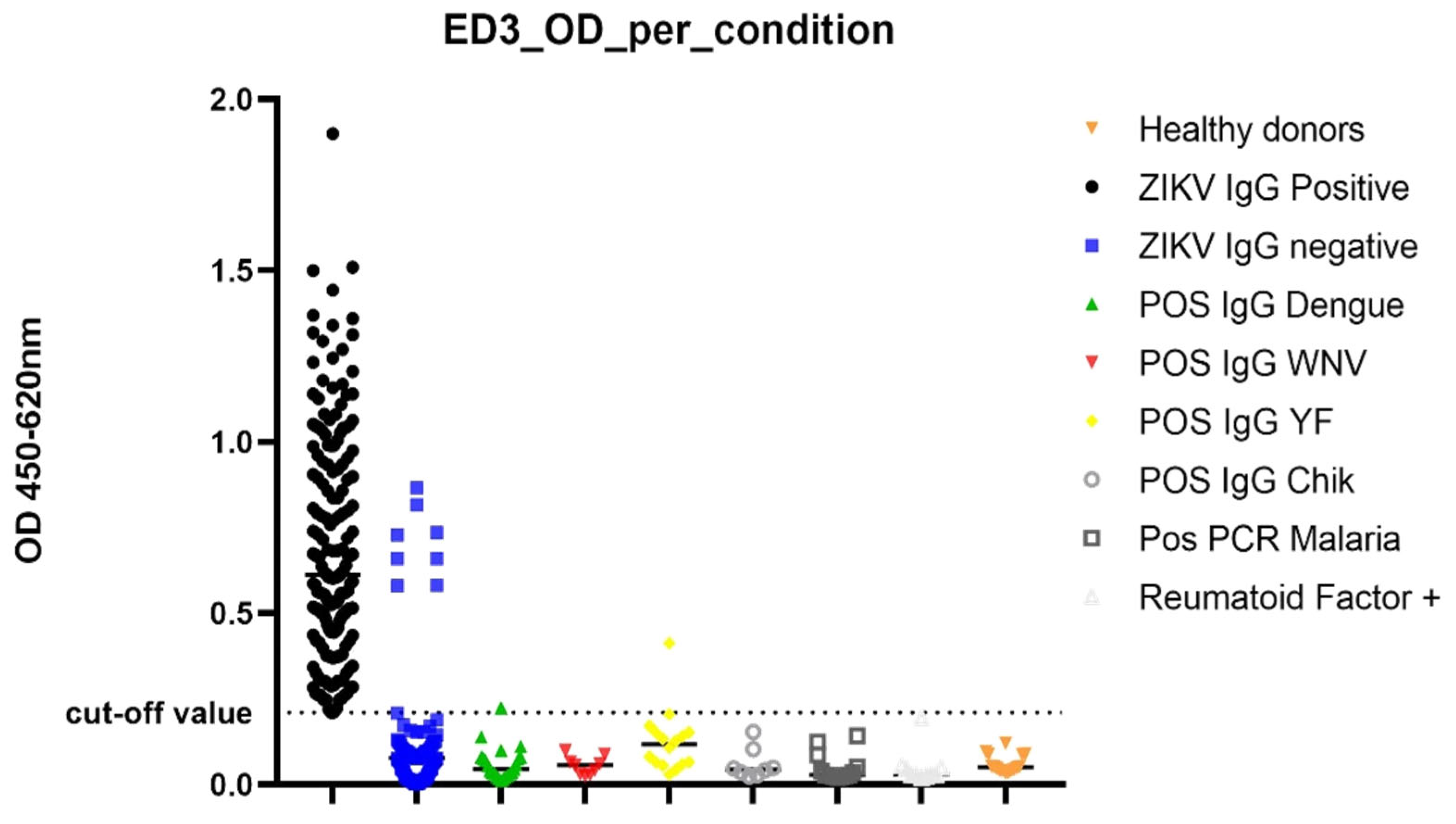
3.1. Determination of Diagnostic Value of the IgG ELISA EDIII
3.2. Diagnostic Performance Calculation with 0.208 as Cut-Off Value
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and Serologic Properties of Zika Virus Associated with an Epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- De Magalhães-Barbosa, M.C.; Prata-Barbosa, A.; Robaina, J.R.; Raymundo, C.E.; Lima-Setta, F.; da Cunha, A.J.L.A. Trends of the Microcephaly and Zika Virus Outbreak in Brazil, January-July 2016. Travel Med. Infect. Dis. 2016, 14, 458–463. [Google Scholar] [CrossRef]
- Araujo, L.M.; Ferreira, M.L.B.; Nascimento, O.J. Guillain-Barré Syndrome Associated with the Zika Virus Outbreak in Brazil. Arq. Neuro-Psiquiatr. 2016, 74, 253–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckerle, I.; Briciu, V.T.; Ergönül, Ö.; Lupşe, M.; Papa, A.; Radulescu, A.; Tsiodras, S.; Tsitou, C.; Drosten, C.; Nussenblatt, V.R. Emerging Souvenirs—Clinical Presentation of the Returning Traveller with Imported Arbovirus Infections in Europe. Clin. Microbiol. Infect. 2018, 24, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, M.L.; St. George, K. Laboratory Diagnosis of Zika Virus Infection. Arch. Pathol. Lab. Med. 2016, 141, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Musso, D.; Rouault, E.; Teissier, A.; Lanteri, M.C.; Zisou, K.; Broult, J.; Grange, E.; Nhan, T.; Aubry, M. Molecular Detection of Zika Virus in Blood and RNA Load Determination during the French Polynesian Outbreak. J. Med. Virol. 2017, 89, 1505–1510. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.-H.; Narayan, R.J. Analytical Methods for Detection of Zika Virus. MRS Commun. 2017, 7, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.; Stevenson, M. Zika Virus Diagnosis: Challenges and Solutions. Clin. Microbiol. Infect. 2019, 25, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Morales, I.; Rosenberger, K.D.; Magalhaes, T.; Morais, C.N.L.; Braga, C.; Marques, E.T.A.; Calvet, G.A.; Damasceno, L.; Brasil, P.; Bispo de Filippis, A.M.; et al. Diagnostic Performance of Anti-Zika Virus IgM, IgAM and IgG ELISAs during Co-Circulation of Zika, Dengue, and Chikungunya Viruses in Brazil and Venezuela. PLoS Negl. Trop. Dis. 2021, 15, e0009336. [Google Scholar] [CrossRef]
- Lee, W.T.; Wong, S.J.; Kulas, K.E.; Dupuis, A.P.; Payne, A.F.; Kramer, L.D.; Dean, A.B.; St. George, K.; White, J.L.; Sommer, J.N.; et al. Development of Zika Virus Serological Testing Strategies in New York State. J. Clin. Microbiol. 2018, 56, e01591-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Laboratory Testing for Zika Virus Infection: Interim Guidance. Available online: http://apps.who.int/iris/bitstream/handle/10665/359857/WHO-ZIKV-DENV-LAB-2022.1-eng.pdf (accessed on 10 August 2022).
- Medialdea-Carrera, R.; Levy, F.; Castanha, P.; Carvalho de Sequeira, P.; Brasil, P.; Lewis-Ximenez, L.L.; Turtle, L.; Solomon, T.; Bispo de Filippis, A.M.; Brown, D.W. A Systematic Evaluation of IgM and IgG Antibody Assay Accuracy in Diagnosing Acute Zika Virus Infection in Brazil: Lessons Relevant to Emerging Infections. J. Clin. Microbiol. 2021, 59, e02893-20. [Google Scholar] [CrossRef] [PubMed]
- Rockstroh, A.; Barzon, L.; Pacenti, M.; Palù, G.; Niedrig, M.; Ulbert, S. Recombinant Envelope-Proteins with Mutations in the Conserved Fusion Loop Allow Specific Serological Diagnosis of Dengue-Infections. PLoS Negl. Trop. Dis. 2015, 9, e0004218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobson-Peters, J. Approaches for the Development of Rapid Serological Assays for Surveillance and Diagnosis of Infections Caused by Zoonotic Flaviviruses of the Japanese Encephalitis Virus Serocomplex. J. Biomed. Biotechnol. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chávez, J.H.; Silva, J.R.; Amarilla, A.A.; Moraes Figueiredo, L.T. Domain III Peptides from Flavivirus Envelope Protein Are Useful Antigens for Serologic Diagnosis and Targets for Immunization. Biologicals 2010, 38, 613–618. [Google Scholar] [CrossRef]
- Mathengtheng, L.; Burt, F.J. Use of Envelope Domain III Protein for Detection and Differentiation of Flaviviruses in the Free State Province, South Africa. Vector Borne Zoonotic Dis. 2014, 14, 261–271. [Google Scholar] [CrossRef]
- Nguyen, N.M.; Duong, B.T.; Azam, M.; Phuong, T.T.; Park, H.; Thuy, P.T.B.; Yeo, S.-J. Diagnostic Performance of Dengue Virus Envelope Domain III in Acute Dengue Infection. Int. J. Mol. Sci. 2019, 20, 3464. [Google Scholar] [CrossRef]
- Chua, A.J.; Vituret, C.; Tan, M.L.; Gonzalez, G.; Boulanger, P.; Ng, M.-L.; Hong, S.-S. A Novel Platform for Virus-like Particle-Display of Flaviviral Envelope Domain III: Induction of Dengue and West Nile Virus Neutralizing Antibodies. Virol. J. 2013, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Ehmen, C.; Medialdea-Carrera, R.; Brown, D.; de Filippis, A.M.B.; de Sequeira, P.C.; Nogueira, R.M.R.; Brasil, P.; Calvet, G.A.; Blessmann, J.; Mallmann, A.-M.; et al. Accurate Detection of Zika Virus IgG Using a Novel Immune Complex Binding ELISA. Trop. Med. Int. Health 2021, 26, 89–101. [Google Scholar] [CrossRef]
- Faye, O.; de Lourdes Monteiro, M.; Vrancken, B.; Prot, M.; Lequime, S.; Diarra, M.; Ndiaye, O.; Valdez, T.; Tavarez, S.; Ramos, J.; et al. Genomic Epidemiology of 2015–2016 Zika Virus Outbreak in Cape Verde. Emerg. Infect. Dis. 2020, 26, 1084–1090. [Google Scholar] [CrossRef]
- Beck, C.; Desprès, P.; Paulous, S.; Vanhomwegen, J.; Lowenski, S.; Nowotny, N.; Durand, B.; Garnier, A.; Blaise-Boisseau, S.; Guitton, E. A High-Performance Multiplex Immunoassay for Serodiagnosis of Flavivirus-Associated Neurological Diseases in Horses. Biomed. Res. Int. 2015, 2015, 678084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wispelaere, M.; Frenkiel, M.-P.; Desprès, P. A Japanese Encephalitis Virus Genotype 5 Molecular Clone Is Highly Neuropathogenic in a Mouse Model: Impact of the Structural Protein Region on Virulence. J. Virol. 2015, 89, 5862–5875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Madrid, A.T.; Porterfield, J.S. A Simple Micro-Culture Method for the Study of Group B Arboviruses. Bull. World Health Organ. 1969, 40, 113–121. [Google Scholar]
- Clopper, C.J.; Pearson, E.S. The Use of Confidence or Fiducial Limits Illustrated in the Case of the Binomial. Biometrika 1934, 26, 404–413. [Google Scholar] [CrossRef]
- Rodrigues, L.C. Microcephaly and Zika Virus Infection. Lancet 2016, 387, 2070–2072. [Google Scholar] [CrossRef] [Green Version]
- Musso, D.; Despres, P. Serological Diagnosis of Flavivirus-Associated Human Infections. Diagnostics 2020, 10, 302. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, R.; Shen, H.; Wang, M.; Yin, Z.; Cheng, A. Structures and Functions of the Envelope Glycoprotein in Flavivirus Infections. Viruses 2017, 9, 338. [Google Scholar] [CrossRef] [Green Version]
- Roehrig, J.T.; Butrapet, S.; Liss, N.M.; Bennett, S.L.; Luy, B.E.; Childers, T.; Boroughs, K.L.; Stovall, J.L.; Calvert, A.E.; Blair, C.D.; et al. Mutation of the Dengue Virus Type 2 Envelope Protein Heparan Sulfate Binding Sites or the Domain III Lateral Ridge Blocks Replication in Vero Cells Prior to Membrane Fusion. Virology 2013, 441, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Yap, T.L.; Hong, S.Y.; Soh, J.H.; Ravichandraprabhu, L.; Lim, V.W.X.; Chan, H.-M.; Ong, T.Z.X.; Chua, Y.P.; Koh, S.E.; Wang, H.; et al. Engineered NS1 for Sensitive, Specific Zika Virus Diagnosis from Patient Serology. Emerg. Infect. Dis. 2021, 27, 1427–1437. [Google Scholar] [CrossRef]
- Volk, D.E.; Beasley, D.W.C.; Kallick, D.A.; Holbrook, M.R.; Barrett, A.D.T.; Gorenstein, D.G. Solution Structure and Antibody Binding Studies of the Envelope Protein Domain III from the New York Strain of West Nile Virus. J. Biol. Chem. 2004, 279, 38755–38761. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Shresta, S. Antigenic Cross-Reactivity between Zika and Dengue Viruses: Is It Time to Develop a Universal Vaccine? Curr. Opin. Immunol. 2019, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Franco, L.; Di Caro, A.; Carletti, F.; Vapalahti, O.; Renaudat, C.; Zeller, H.; Tenorio, A. Recent Expansion of Dengue Virus Serotype 3 in West Africa. Eurosurveillance 2010, 15, 19490. [Google Scholar] [CrossRef] [PubMed]
- Puschnik, A.; Lau, L.; Cromwell, E.A.; Balmaseda, A.; Zompi, S.; Harris, E. Correlation between Dengue-Specific Neutralizing Antibodies and Serum Avidity in Primary and Secondary Dengue Virus 3 Natural Infections in Humans. PLoS Negl. Trop. Dis. 2013, 7, e2274. [Google Scholar] [CrossRef] [Green Version]
- Heinz, F.X.; Stiasny, K. The Antigenic Structure of Zika Virus and Its Relation to Other Flaviviruses: Implications for Infection and Immunoprophylaxis. Microbiol. Mol. Biol. Rev. 2017, 81, e00055-16. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specificity Panel | |
| Characterizations | n = |
| Related flaviviruses | |
| POS IgG DENV | 23 |
| POS IgG WNV | 9 |
| POS IgG YF | 16 |
| Alphavirus | |
| POS IgG CHIKV | 8 |
| Other interfering conditions | |
| Pos PCR Malaria | 13 |
| POS Rheumatoid Factor | 10 |
| Healthy donors | |
| NEG All ** | 16 |
| Symptomatic samples | |
| ZIKV IgG negative | 93 |
| TOTAL | 188 |
| Viral Strain | Position in E Sequence | EDIII Sequence |
|---|---|---|
| PF-25013-18 | 297–341 | LRLKGVSYSLCTAAFTFTKIPAETLHGTVTVEVQYAGTDGPCKVP |
| MR766 | LRLKGVSYSLCTAAFTFTKIPAETLHGTVTVEVQYAGTDGPCKVP | |
| PF-25013-18 | 342–387 | AQMAVDMQTLTPVGRLITANPVITESTENSKMMLELDPPFGDSYIV |
| MR766 | VQMAVDMQTLTPVGRLITANPVITESTENSKMMLELDPPFGDSYIV | |
| PF-25013-18 | 388–408 | IGVGEKKITHHWHRSGSTIGK |
| MR766 | IGVGEKKITHHWHRSGSTIGK |
| Parameter | Value |
|---|---|
| Area | 0.974 |
| Std. Error | 0.009 |
| 95% CI | 0.974–0.992 |
| p value | <0.0001 |
| Parameters | Values |
|---|---|
| Sensitivity | 100.0% [97.4–100.0] |
| Specificity | 94.7% [90.3–97.2] |
| Likelihood Ratio+ | 18.8% |
| Likelihood Ratio− | 0% |
| TP | 179 |
| TN | 178 |
| FP | 10 |
| FN | 0 |
| Accuracy | 97.3% [95.7–99.2] |
| Panel Tested | Condition Tested | Reference Test | Analysis Values | Sensitivity | Specificity | n | Analyzed Method | Reference Method | |
|---|---|---|---|---|---|---|---|---|---|
| pos | neg | ||||||||
| Sensitivity Panel | ZIKV IgG POS | ZIKV PRNT | estimate: | 100.0% | 94.7% | 367 | pos | 179 | 10 |
| 95% CI: | [97.4–100.0] | [90.3–97.2] | |||||||
| p-value | 2.2 × 10−16 | 2.2 × 10−16 | neg | 0 | 178 | ||||
| Specificity Panel | POS IgG Dengue | ZIKV PRNT | estimate: | 95.6% | 23 | pos | 1 | ||
| 95% CI: | [78.0–99.9] | ||||||||
| p-value | 5.72 × 10−6 | neg | 22 | ||||||
| POS IgG WNV | ZIKV PRNT | estimate: | 100.0% | 9 | pos | 0 | |||
| 95% CI: | [66.4–100.0] | ||||||||
| p-value | 0.0039 | neg | 9 | ||||||
| POS IgG YFV | ZIKV PRNT | estimate: | 93.7% | 16 | pos | 1 | |||
| 95% CI: | [69.8–99.8] | ||||||||
| p-value | 0.0005 | neg | 15 | ||||||
| POS IgG CHIKV | ZIKV PRNT | estimate: | 100.00% | 8 | pos | 0 | |||
| 95% CI: | [63.1–100.0] | ||||||||
| p-value | 0.007812 | neg | 8 | ||||||
| POS PCR Malaria | ZIKV PRNT | estimate: | 100.0% | 13 | pos | 0 | |||
| 95% CI: | [75.3–100.0] | ||||||||
| p-value | 0.0002 | neg | 13 | ||||||
| POS Rheumatoid Factor | ZIKV PRNT | estimate: | 100.0% | 10 | pos | 0 | |||
| 95% CI: | [69.1–100.0] | ||||||||
| p-value | 0.0020 | neg | 10 | ||||||
| NEG all ** | ZIKV PRNT | estimate: | 100.0% | 16 | pos | 0 | |||
| 95% CI: | [79.4–100.0] | ||||||||
| p-value | 3.052 × 10−5 | neg | 16 | ||||||
| Symptomatic IgG Neg ZIKV | ZIKV PRNT | estimate: | 91.4% | 93 | pos | 8 | |||
| 95% CI: | [83.7–96.2] | ||||||||
| p-value | 2.2 × 10−16 | neg | 85 ** | ||||||
| Type of Test | Advantages | Disadvantages |
|---|---|---|
| EDIII IgG ELISA | High sensitivity and specificity Easy to set up High throughput Short “hands-on time” (4 h) Cheap | Semi quantitative |
| Standard Indirect IgG ELISA | High throughput Short “hands-on time” (5 h) | Low specificity (high cross reactivity) Use of animal tissue (mouse brain) Semi quantitative |
| PRNT Test | High sensitivity and specificity Measures neutralizing activity Quantitative method | Requires a BSL 3 Lab Requires highly trained personnel Expensive Unable to differentiate antibody classes Slow (4–6 days) to obtain results |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndiaye, O.; Diagne, C.T.; Abd El Wahed, A.; Dia, F.; Dia, M.; Faye, A.; Leal, S.D.V.; dos Santos, M.; Lima Mendonça, M.d.L.d.; da Silva Leite, C.C.; et al. Use of Envelope Domain III Protein for the Detection of IgG Type Antibodies Specific to Zika Virus by Indirect ELISA. Diagnostics 2023, 13, 462. https://doi.org/10.3390/diagnostics13030462
Ndiaye O, Diagne CT, Abd El Wahed A, Dia F, Dia M, Faye A, Leal SDV, dos Santos M, Lima Mendonça MdLd, da Silva Leite CC, et al. Use of Envelope Domain III Protein for the Detection of IgG Type Antibodies Specific to Zika Virus by Indirect ELISA. Diagnostics. 2023; 13(3):462. https://doi.org/10.3390/diagnostics13030462
Chicago/Turabian StyleNdiaye, Oumar, Cheikh Tidiane Diagne, Ahmed Abd El Wahed, Fatou Dia, Moussa Dia, Adama Faye, Silvania Da Veiga Leal, Menilita dos Santos, Maria da Luz de Lima Mendonça, Carolina Cardoso da Silva Leite, and et al. 2023. "Use of Envelope Domain III Protein for the Detection of IgG Type Antibodies Specific to Zika Virus by Indirect ELISA" Diagnostics 13, no. 3: 462. https://doi.org/10.3390/diagnostics13030462
APA StyleNdiaye, O., Diagne, C. T., Abd El Wahed, A., Dia, F., Dia, M., Faye, A., Leal, S. D. V., dos Santos, M., Lima Mendonça, M. d. L. d., da Silva Leite, C. C., Bouh Boye, C. S., Bryant, J. E., Desprès, P., Faye, O., Sall, A. A., & Faye, O. (2023). Use of Envelope Domain III Protein for the Detection of IgG Type Antibodies Specific to Zika Virus by Indirect ELISA. Diagnostics, 13(3), 462. https://doi.org/10.3390/diagnostics13030462