Altered Gut Microbiota in Irritable Bowel Syndrome and Its Association with Food Components


 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Setting and Subjects
2.2. Data Collection
2.3. Outcome Meaures
2.3.1. Assessment of Daily Food Components
2.3.2. Fecal Sample DNA Extraction and Microbiome Sequencing
2.4. Statistical Analysis
3. Results
3.1. Demographic Characteristics of the IBS and HC Groups
3.2. Food Componnets in the IBS and HC Groups
3.3. Fecal Microbiota Pattern in the IBS and HC Groups
3.3.1. Total Number of OTUs
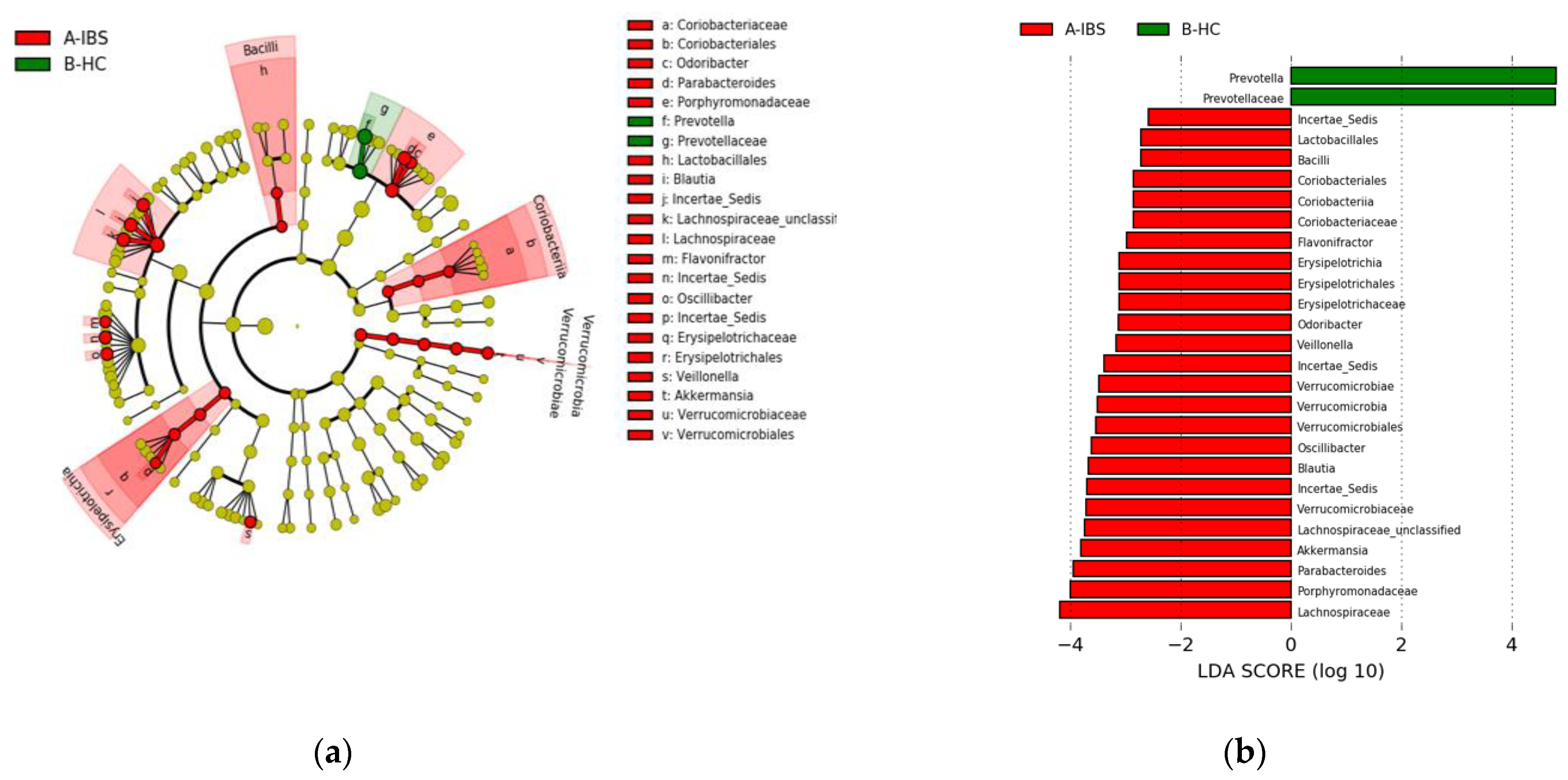
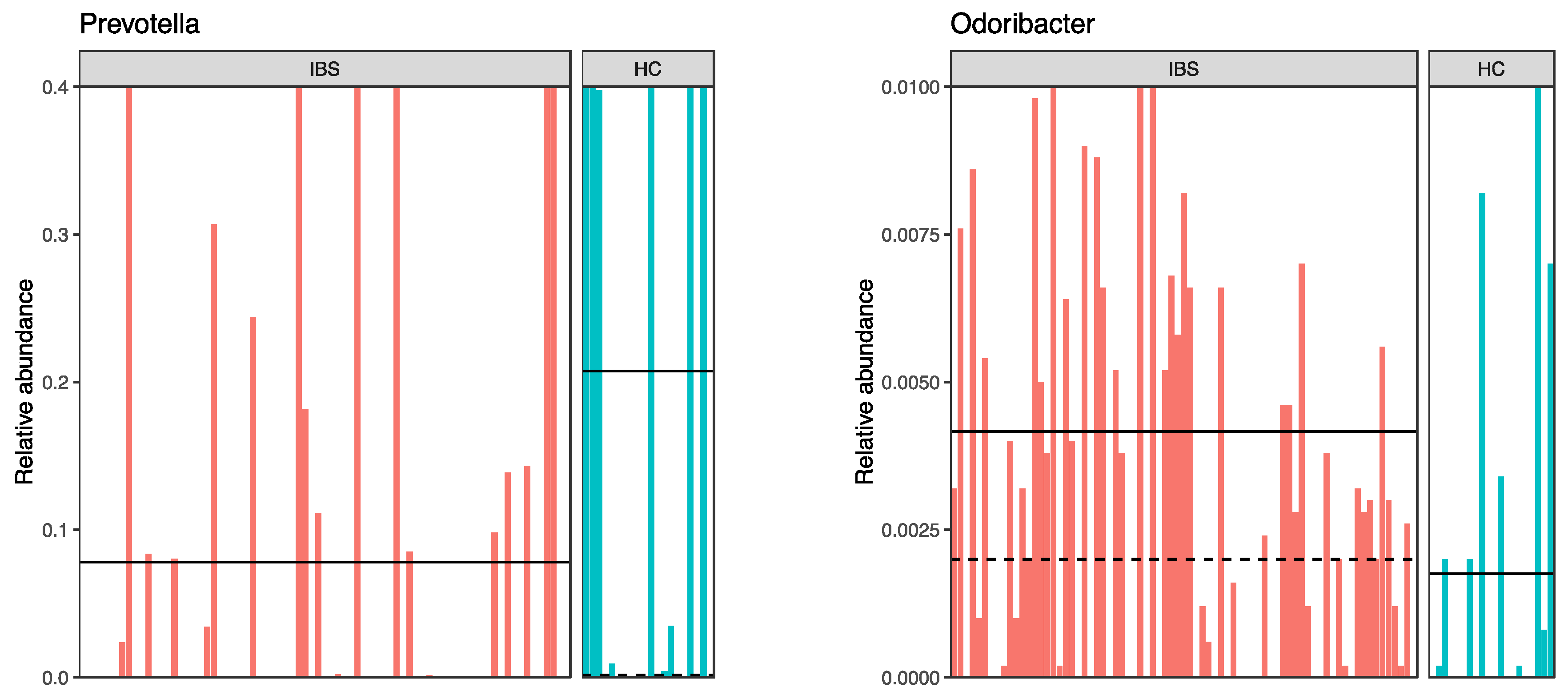
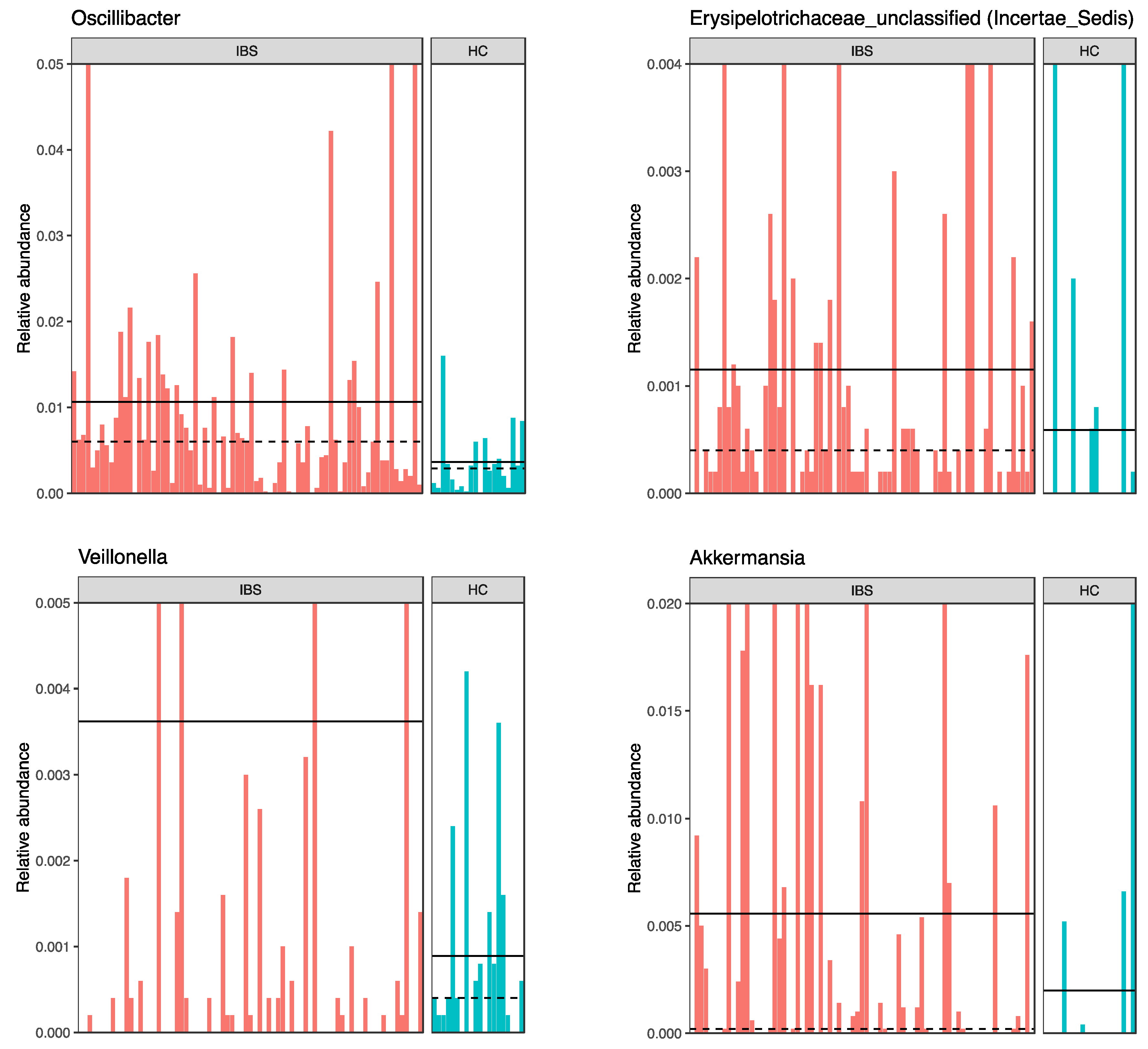
3.3.2. Fecal Microbiota Composition in the IBS Compared to the HC
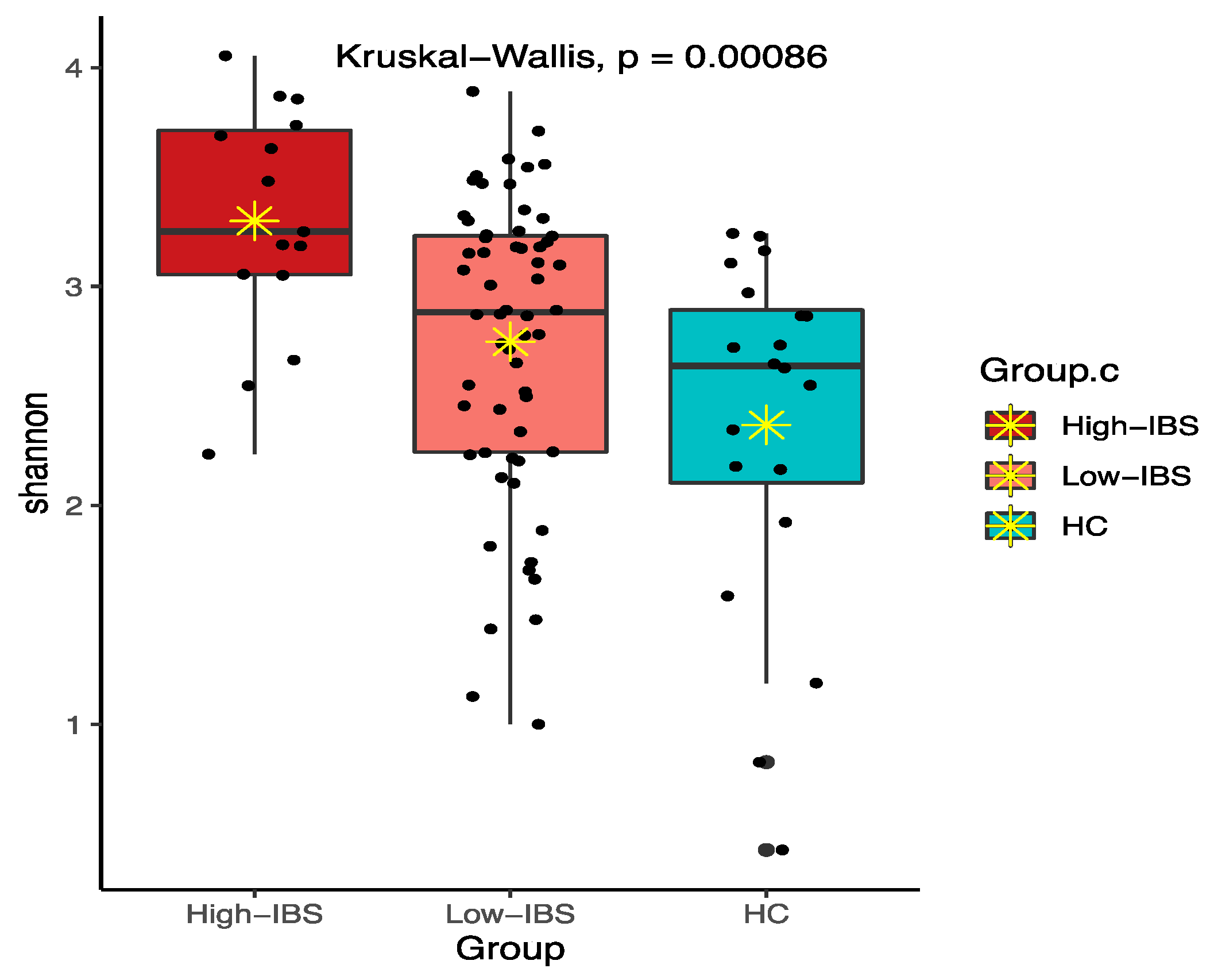
3.3.3. Fecal Microbiota Diversity in the IBS Compared to the HC Group
3.4. Associations between Fecal Microbiota Diversity and Food Component Intakes
3.5. Fecal Microbiota Patterns Associated with the Daily Caffeine Intake
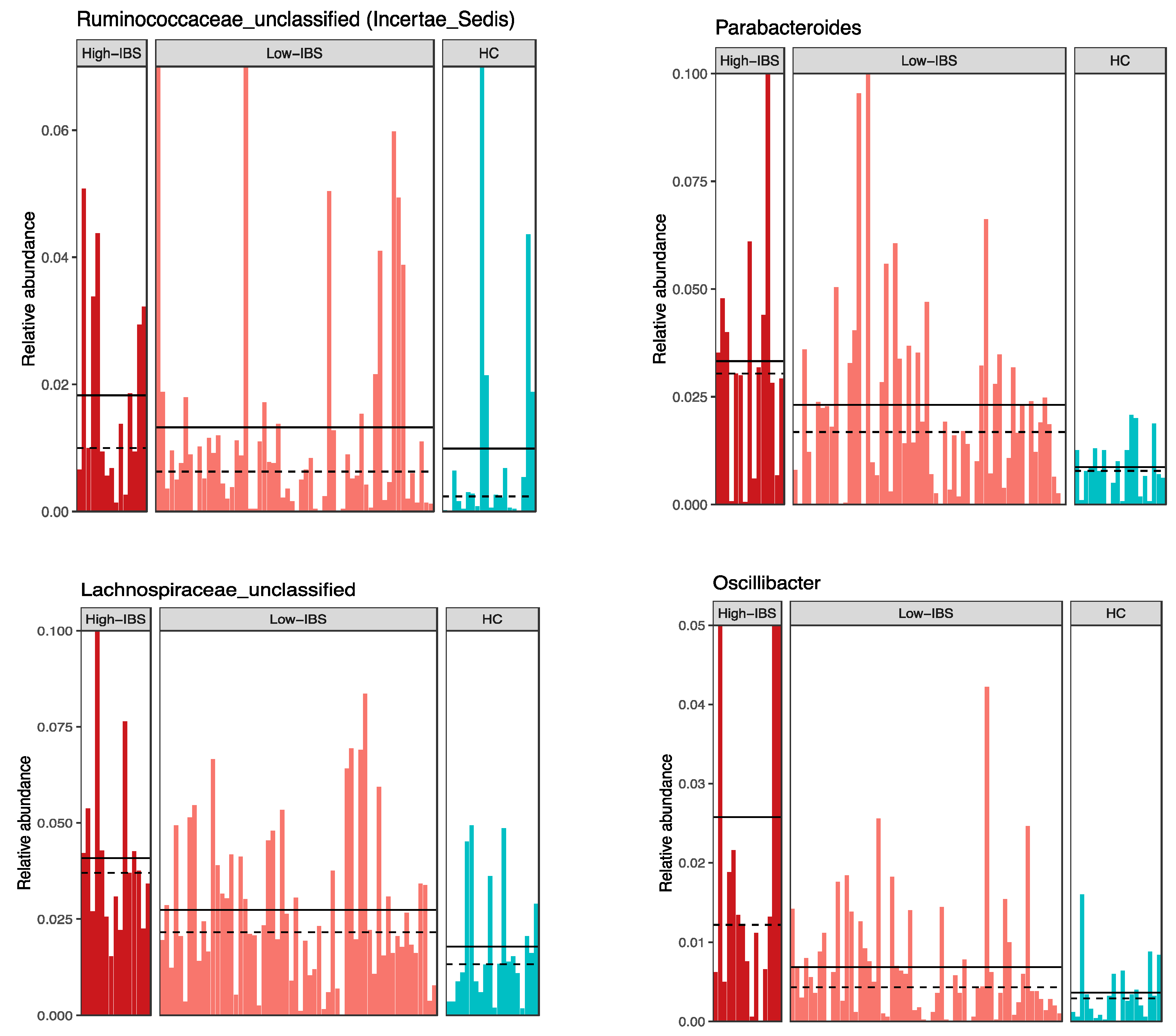
3.6. Associations between Fecal Microbiota Diversity and Food Component Intakes in the High-IBS, Low-IBS, and HC Groups
4. Discussion
4.1. Differences in Food Components between IBS and HC
4.2. Differences in the Gut Microbiota between IBS and HC
4.3. Correlations between Food Components and the Gut Microbiota in IBS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herndon, C.C.; Wang, Y.; Lu, C.-L. Targeting the gut microbiota for the treatment of irritable bowel syndrome. Kaohsiung J. Med. Sci. 2020, 36, 160–170. [Google Scholar] [CrossRef]
- Dimidi, E.; Rossi, M.; Whelan, K. Irritable bowel syndrome and diet: Where are we in 2018? Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Mazzawi, T.; El-Salhy, M. Effect of diet and individual dietary guidance on gastrointestinal endocrine cells in patients with irritable bowel syndrome. Int. J. Mol. Med. 2017, 40, 943–952. [Google Scholar] [CrossRef]
- Valeur, J.; Småstuen, M.C.; Knudsen, T.; Lied, G.A.; Røseth, A.G. Exploring Gut Microbiota Composition as an Indicator of Clinical Response to Dietary FODMAP Restriction in Patients with Irritable Bowel Syndrome. Dig. Dis. Sci. 2018, 63, 429–436. [Google Scholar] [CrossRef]
- Rajilić-Stojanović, M.; Jonkers, D.M.; Salonen, A.H.; Hanevik, K.; Raes, J.; Jalanka, J.; De Vos, W.M.; Manichanh, C.; Golic, N.; Enck, P.; et al. Intestinal Microbiota And Diet in IBS: Causes, Consequences, or Epiphenomena? Am. J. Gastroenterol. 2015, 110, 278–287. [Google Scholar] [CrossRef] [Green Version]
- El-Salhy, M.; Hatlebakk, J.G.; Hausken, T. Diet in Irritable Bowel Syndrome (IBS): Interaction with Gut Microbiota and Gut Hormones. Nutrients 2019, 11, 1824. [Google Scholar] [CrossRef] [Green Version]
- Altobelli, E.; Del Negro, V.; Angeletti, P.M.; Latella, G. Low-FODMAP Diet Improves Irritable Bowel Syndrome Symptoms: A Meta-Analysis. Nutrients 2017, 9, 940. [Google Scholar] [CrossRef]
- Pozuelo, M.; Panda, S.; Santiago, A.; Mendez, S.; Accarino, A.; Santos, J.; Guarner, F.; Azpiroz, F.; Manichanh, C. Reduction of butyrate- and methane-producing microorganisms in patients with Irritable Bowel Syndrome. Sci. Rep. 2015, 5, 12693. [Google Scholar] [CrossRef] [Green Version]
- Chong, P.P.; Chin, V.K.; Looi, C.Y.; Wong, W.F.; Madhavan, P.; Yong, V.C. The Microbiome and Irritable Bowel Syndrome—A Review on the Pathophysiology, Current Research and Future Therapy. Front. Microbiol. 2019, 10, 1136. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Rajilić-Stojanović, M. Function of the microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 5–16. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Ooi, S.L.; Correa, D.; Park, S.C. Probiotics, prebiotics, and low FODMAP diet for irritable bowel syndrome—What is the current evidence? Complement. Ther. Med. 2019, 43, 73–80. [Google Scholar] [CrossRef]
- Harris, L.A.; Baffy, N. Modulation of the gut microbiota: A focus on treatments for irritable bowel syndrome. Postgrad. Med. 2017, 129, 872–888. [Google Scholar] [CrossRef]
- Mansour, A.; Mohajeri-Tehrani, M.R.; Karimi, S.; Sanginabadi, M.; Poustchi, H.; Enayati, S.; Asgarbeik, S.; Nasrollahzadeh, J.; Hekmatdoost, A. Short term effects of coffee components consumption on gut microbiota in patients with non-alcoholic fatty liver and diabetes: A pilot randomized placebo-controlled, clinical trial. EXCLI J. 2020, 19, 241–250. [Google Scholar] [PubMed]
- Gurwara, S.; Dai, A.; Ajami, N.; El-Serag, H.B.; Graham, D.Y.; Jiao, L. 196 Caffeine Consumption and the Colonic Mucosa-Associated Gut Microbiota. Am. J. Gastroenterol. 2019, 114, S119–S120. [Google Scholar] [CrossRef]
- Cong, X.; Ramesh, D.; Perry, M.; Xu, W.; Bernier, K.M.; Young, E.E.; Walsh, S.; Starkweather, A. Pain self-management plus nurse-led support in young adults with irritable bowel syndrome: Study protocol for a pilot randomized control trial. Res. Nurs. Health 2018, 41, 121–130. [Google Scholar] [CrossRef]
- Kristal, A.R.; Feng, Z.; Coates, R.J.; Oberman, A.; George, V. Associations of race/ethnicity, education, and dietary intervention with the validity and reliability of a food frequency questionnaire: The Women’s Health Trial Feasibility Study in Minority Populations. Am. J. Epidemiol. 1997, 146, 856–869. [Google Scholar] [CrossRef]
- (NIH), National Institutes of Health. Diet History Questionnaire II (DHQ II): Diet*Calc Software. 2020. Available online: https://epi.grants.cancer.gov/dhq2/dietcalc/ (accessed on June 2020).
- Cong, X.; Judge, M.; Xu, W.; Diallo, A.; Janton, S.; Brownell, E.A.; Maas, K.; Graf, J. Influence of Feeding Type on Gut Microbiome Development in Hospitalized Preterm Infants. Nurs. Res. 2017, 66, 123–133. [Google Scholar] [CrossRef]
- Cong, X.; Xu, W.; Janton, S.; Henderson, W.A.; Matson, A.; McGrath, J.M.; Maas, K.; Graf, J. Gut Microbiome Developmental Patterns in Early Life of Preterm Infants: Impacts of Feeding and Gender. PLoS ONE 2016, 11, e0152751. [Google Scholar] [CrossRef]
- (FDA), U.S.; Food and Drug Administration. Spilling the Beans: How Much Caffeine Is Too Much. 30 October 2020. Available online: https://www.fda.gov/consumers/consumer-updates/spilling-beans-how-much-caffeine-too-much (accessed on June 2020).
- Cho, H.-W. How Much Caffeine is Too Much for Young Adolescents? Osong Public Health Res. Perspect. 2018, 9, 287–288. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.; Naghibi, M.M.; Garcha, D. The Role of Bacteria, Probiotics and Diet in Irritable Bowel Syndrome. Foods 2018, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Salhy, M.; Østgaard, H.; Hausken, T.; Gundersen, D. Diet and effects of diet management on quality of life and symptoms in patients with irritable bowel syndrome. Mol. Med. Rep. 2012, 5, 1382–1390. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.A.; Locke, G.R.; Weaver, A.L.; Zinsmeister, A.R.; Talley, N.J. Diet and Functional Gastrointestinal Disorders: A Population-Based Case-Control Study. Am. J. Gastroenterol. 2005, 100, 2743–2748. [Google Scholar] [CrossRef]
- Reding, K.W.; Cain, K.C.; Jarrett, M.E.; Eugenio, M.D.; Heitkemper, M.M. Relationship Between Patterns of Alcohol Consumption and Gastrointestinal Symptoms Among Patients with Irritable Bowel Syndrome. Am. J. Gastroenterol. 2013, 108, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Heizer, W.D.; Southern, S.; McGovern, S. The Role of Diet in Symptoms of Irritable Bowel Syndrome in Adults: A Narrative Review. J. Am. Diet. Assoc. 2009, 109, 1204–1214. [Google Scholar] [CrossRef]
- Böhn, L.; Störsrud, S.; Liljebo, T.M.; Collin, L.; Lindfors, P.; Törnblom, H.; Simrén, M. Diet Low in FODMAPs Reduces Symptoms of Irritable Bowel Syndrome as Well as Traditional Dietary Advice: A Randomized Controlled Trial. Gastroenterology 2015, 149, 1399–1407.e2. [Google Scholar] [CrossRef] [Green Version]
- Eswaran, S.; Muir, J.; Chey, W.D. Fiber and Functional Gastrointestinal Disorders. Am. J. Gastroenterol. 2013, 108, 718–727. [Google Scholar] [CrossRef]
- Staudacher, H.M.; Whelan, K. Altered gastrointestinal microbiota in irritable bowel syndrome and its modification by diet: Probiotics, prebiotics and the low FODMAP diet. Proc. Nutr. Soc. 2016, 75, 306–318. [Google Scholar] [CrossRef]
- Backhed, F. Host-Bacterial Mutualism in the Human Intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Cong, X.; Xu, W.; Romisher, R.; Poveda, S.; Forte, S.; Starkweather, A.; Henderson, W.A. Gut microbiome and infant health: Brain-gut-microbiota axis and host genetic factors. Yale J. Biol. Med. 2016, 89, 299. [Google Scholar] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, X.; Tian, Z.; Li, L.; Zeng, Z.; Chen, M.; Xiong, L. Fecal Microbiota Alterations Associated With Diarrhea-Predominant Irritable Bowel Syndrome. Front. Microbiol. 2018, 9, 1600. [Google Scholar] [CrossRef]
- Rangel, I.; Sundin, J.; Fuentes, S.; Repsilber, D.; De Vos, W.M.; Brummer, R.J. The relationship between faecal-associated and mucosal-associated microbiota in irritable bowel syndrome patients and healthy subjects. Aliment. Pharmacol. Ther. 2015, 42, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Ringel-Kulka, T.; Benson, A.K.; Carroll, I.M.; Kim, J.; Legge, R.M.; Ringel, Y. Molecular characterization of the intestinal microbiota in patients with and without abdominal bloating. Am. J. Physiol. Liver Physiol. 2016, 310, G417–G426. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Tap, J.; Furet, J.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G.; et al. Gut microbiota richness promotes its stability upon increased dietary fibre intake in healthy adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M.; et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.A.; Verdi, S.; Maxan, M.-E.; Shin, C.M.; Zierer, J.; Bowyer, R.C.E.; Martin, T.; Williams, F.M.K.; Menni, C.; Bell, J.T.; et al. Gut microbiota associations with common diseases and prescription medications in a population-based cohort. Nat. Commun. 2018, 9, 2655. [Google Scholar] [CrossRef] [Green Version]
- Brunkwall, L.; Ericson, U.; Nilsson, P.M.; Orho-Melander, M.; Ohlsson, B. Self-reported bowel symptoms are associated with differences in overall gut microbiota composition and enrichment of Blautia in a population-based cohort. J. Gastroenterol. Hepatol. 2020. [Google Scholar] [CrossRef]
- Jeffery, I.B.; Das, A.; O’Herlihy, E.; Coughlan, S.; Cisek, K.; Moore, M.; Bradley, F.; Carty, T.; Pradhan, M.; Dwibedi, C.; et al. Differences in Fecal Microbiomes and Metabolomes of People With vs Without Irritable Bowel Syndrome and Bile Acid Malabsorption. Gastroenterology 2020, 158, 1016–1028.e8. [Google Scholar] [CrossRef] [PubMed]
- Hugerth, L.W.; Andreasson, A.; Talley, N.J.; Forsberg, A.M.; Kjellström, L.; Schmidt, P.T.; Agreus, L.; Engstrand, L. No distinct microbiome signature of irritable bowel syndrome found in a Swedish random population. Gut 2019, 69, 1076–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittayanon, R.; Lau, J.T.; Yuan, Y.; Leontiadis, G.I.; Tse, F.; Surette, M.; Moayyedi, P. Gut Microbiota in Patients With Irritable Bowel Syndrome—A Systematic Review. Gastroenterology 2019, 157, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labus, J.S.; Hollister, E.B.; Jacobs, J.P.; Kirbach, K.; Oezguen, N.; Gupta, A.; Acosta, J.; Luna, R.A.; Aagaard, K.M.; Versalovic, J.; et al. Differences in gut microbial composition correlate with regional brain volumes in irritable bowel syndrome. Microbiome 2017, 5, 49. [Google Scholar] [CrossRef]
- Lee, B.J.; Bak, Y.T. Irritable Bowel Syndrome, Gut Microbiota and Probiotics. J. Neurogastroenterol. Motil. 2011, 17, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Staudacher, H.M.; Lomer, M.C.E.; Anderson, J.L.; Barrett, J.S.; Muir, J.G.; Irving, P.M.; Whelan, K. Fermentable Carbohydrate Restriction Reduces Luminal Bifidobacteria and Gastrointestinal Symptoms in Patients with Irritable Bowel Syndrome. J. Nutr. 2012, 142, 1510–1518. [Google Scholar] [CrossRef]
- Shepherd, S.J.; Parker, F.C.; Muir, J.G.; Gibson, P.R. Dietary Triggers of Abdominal Symptoms in Patients With Irritable Bowel Syndrome: Randomized Placebo-Controlled Evidence. Clin. Gastroenterol. Hepatol. 2008, 6, 765–771. [Google Scholar] [CrossRef]
- Halmos, E.P.; Christophersen, C.T.; Bird, A.R.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. Diets that differ in their FODMAP content alter the colonic luminal microenvironment. Gut 2015, 64, 93–100. [Google Scholar] [CrossRef]
- Barrett, J.S. How to institute the low-FODMAP diet. J. Gastroenterol. Hepatol. 2017, 32, 8–10. [Google Scholar] [CrossRef] [Green Version]
- James, S.C.; Fraser, K.; Young, W.; McNabb, W.C.; Roy, N.C. Gut Microbial Metabolites and Biochemical Pathways Involved in Irritable Bowel Syndrome: Effects of Diet and Nutrition on the Microbiome. J. Nutr. 2020, 150, 1012–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannampalli, P.; Shaker, R.; Sengupta, J.N. Colonic butyrate-algesic or analgesic? Neurogastroenterol. Motil. 2011, 23, 975–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, S.; Salazar, N.; Ruiz-Saavedra, S.; Gómez-Martín, M.; Reyes-Gavilán, C.G.D.L.; Gueimonde, M. Long-Term Coffee Consumption is Associated with Fecal Microbial Composition in Humans. Nutrients 2020, 12, 1287. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Watanabe, S.; Xiao, J.; Nagatomo, R.; Ogawa, H.; Tsunematsu, T.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; et al. Effect of coffee or coffee components on gut microbiome and short-chain fatty acids in a mouse model of metabolic syndrome. Sci. Rep. 2018, 8, 16173. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographics | N | HC (n = 21) | IBS (n = 80) | p-Value | |
|---|---|---|---|---|---|
| Gender | |||||
| Female | 72 | 11 (52.38%) | 61 (76.25%) | 0.060 | |
| Male | 29 | 10 (47.619%) | 19 (23.75%) | ||
| Race | |||||
| White | 71 | 9 (42.86%) | 62 (77.50%) | 0.070 | |
| Asian | 16 | 6 (28.57%) | 10 (12.50%) | ||
| African–American | 12 | 4 (19.05%) | 8 (10.00%) | ||
| Not reported | 2 | 2 (9.52%) | 0 (0.00%) | ||
| Ethnicity | |||||
| Non-Hispanic | 84 | 16 (76.19%) | 68 (85.00%) | 0.360 | |
| Hispanic | 11 | 4 (19.05%) | 7 (8.75%) | ||
| Not reported | 6 | 1 (4.76%) | 5 (6.25%) | ||
| Education | |||||
| High school or lower | 8 | 2 (9.52%) | 6 (7.50%) | 0.151 | |
| Some college | 63 | 16 (76.19%) | 47 (58.75%) | ||
| Associate degree | 3 | 1 (4.76%) | 1 (1.25%) | ||
| Bachelor degree | 16 | 2 (9.52%) | 14 (17.50%) | ||
| Master degree | 12 | 0 (0.00%) | 12 (15.00%) | ||
| Primary caregiver | |||||
| Parent/legal guardian | 53 | 14 (66.67%) | 39 (48.75%) | 0.117 | |
| Self | 46 | 6 (28.57%) | 40 (50.00%) | ||
| Other | 2 | 1 (4.76%) | 1 (1.25%) | ||
| Employment status | 0.269 | ||||
| Student | 75 | 18 (85.71%) | 57 (71.25%) | ||
| Working now | 22 | 2 (9.52%) | 20 (25.00%) | ||
| Unemployed | 4 | 1 (4.76%) | 3 (3.75%) | ||
| Marital status | |||||
| Never married | 98 | 21 (100.00%) | 77 (96.25%) | 1 | |
| Married | 3 | 0 (0.00%) | 3 (3.75%) | ||
| IBS subtype | |||||
| IBS-C | 9 | N/A | 9 (11.00%) | ||
| IBS-D | 5 | N/A | 5 (7.00%) | ||
| IBS-M | 66 | N/A | 66 (82.00%) | ||
| Medical care setting type | |||||
| Primary | 15 | N/A | 15 (19.00%) | ||
| Secondary | 6 | N/A | 6 (7.00%) | ||
| Primary + secondary | 22 | N/A | 22 (28.00%) | ||
| None | 37 | N/A | 37 (46.00%) | ||
| Mean (SD) | Range | ||||
| HC | IBS | IBS | HC | p-value | |
| Age (years) | 20.14 (1.39) | 20.39 (2.57) | 18–23 | 18–28 | 0.071 |
| Household members | 4.19 (1.81) | 3.29 (1.48) | 1–9 | 1–7 | 0.034 |
| Duration of IBS (years) | N/A | 4.01 (2.67) | 1–13 | N/A | N/A |
| Mean (SD) | Median (Range) | |||
|---|---|---|---|---|
| HC | IBS | HC | IBS | |
| Food energy (kcal) | 1965.82 (791.06) | 1793.41 (761.95) | 1837.46 (705.94–3768.59) | 1692.51 (320.12–4223.59) |
| Protein (g) | 83.31 (37.13) | 73.29 (39.40) | 83.05 (21.84–147.22) | 60.48 (10.61–216.97) |
| Fat (g) | 87.80 (41.55) | 77.46 (36.56) | 96.45 (21.81–182.24) | 73.40 (9.90–176.68) |
| Cholesterol (mg) | 278.76 (84.29) | 228.06 (123.41) | 291.71 (52.14–507.28) | 205.45 (9.02–543.25) |
| Carbohydrate (g) | 211.73 (84.29) | 201.33 (84.90) | 194.76 (86.60–398.42) | 186.39 (27.94–453.60) |
| Dietary fiber (g) | 19.79 (9.06) | 18.44 (10.03) | 16.97 (5.55–39.52) | 17.31 (3.54–66.61) |
| Alcohol (g) | 4.99 (4.56) | 5.56 (5.19) | 4.06 (0.01–15.60) | 4.30 (0.00–19.16) |
| Caffeine (mg) * | 82.93 (94.67) | 246.42 (297.42) | 38.24 (0.55–293.77) | 129.92 (0.06–1273.84) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barandouzi, Z.A.; Lee, J.; Maas, K.; Starkweather, A.R.; Cong, X.S. Altered Gut Microbiota in Irritable Bowel Syndrome and Its Association with Food Components. J. Pers. Med. 2021, 11, 35. https://doi.org/10.3390/jpm11010035
Barandouzi ZA, Lee J, Maas K, Starkweather AR, Cong XS. Altered Gut Microbiota in Irritable Bowel Syndrome and Its Association with Food Components. Journal of Personalized Medicine. 2021; 11(1):35. https://doi.org/10.3390/jpm11010035
Chicago/Turabian StyleBarandouzi, Zahra A., Joochul Lee, Kendra Maas, Angela R. Starkweather, and Xiaomei S. Cong. 2021. "Altered Gut Microbiota in Irritable Bowel Syndrome and Its Association with Food Components" Journal of Personalized Medicine 11, no. 1: 35. https://doi.org/10.3390/jpm11010035
APA StyleBarandouzi, Z. A., Lee, J., Maas, K., Starkweather, A. R., & Cong, X. S. (2021). Altered Gut Microbiota in Irritable Bowel Syndrome and Its Association with Food Components. Journal of Personalized Medicine, 11(1), 35. https://doi.org/10.3390/jpm11010035