
TMS-EEG Research to Elucidate the Pathophysiological Neural Bases in Patients with Schizophrenia: A Systematic Review



Abstract
:1. Introduction
1.1. Overview of Schizophrenia
1.2. Technical Advance of Combined TMS-EEG
1.3. TMS-EEG Neurophysiology
1.3.1. GABAergic (Short-Interval Intracortical Inhibition: SICI and Long-Interval Intracortical Inhibition: LICI) and Glutamatergic (Intracortical Facilitation: ICF) Neurophysiological Functions
1.3.2. Cholinergic (Short-Latency Afferent Inhibition: SAI) Neurophysiological Function
1.3.3. Other Neurophysiological Measures in TMS-EEG (Power, ERSP, Coherence, and Natural Frequency)
1.4. Objectives of This Systematic Review
2. Materials and Methods
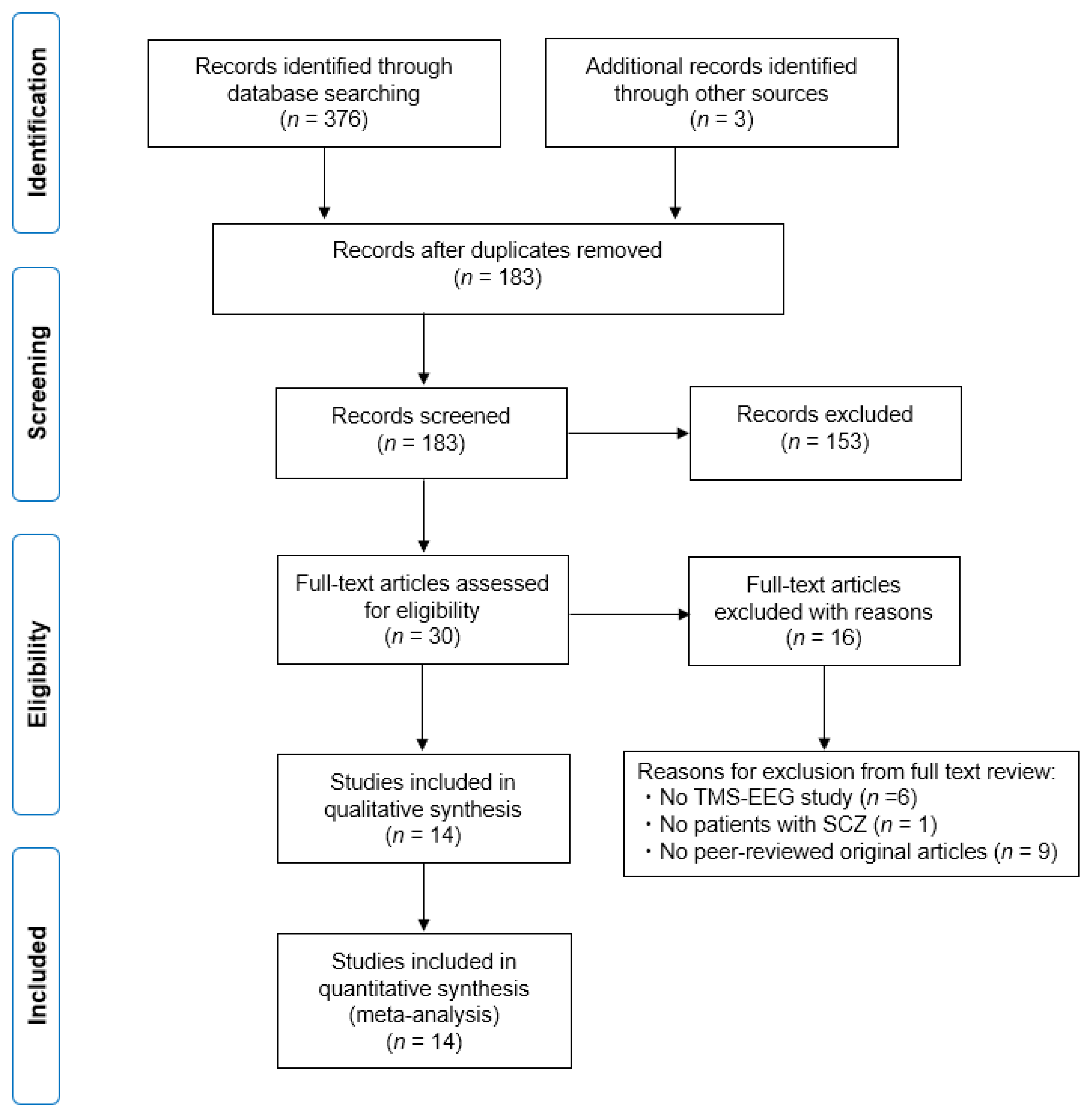
2.1. Study Search and Selection Strategy
2.2. Data Extraction
2.3. Outcome Measures
2.4. Risk of Bias Assessment
3. Results
3.1. Characteristics of the Included Studies
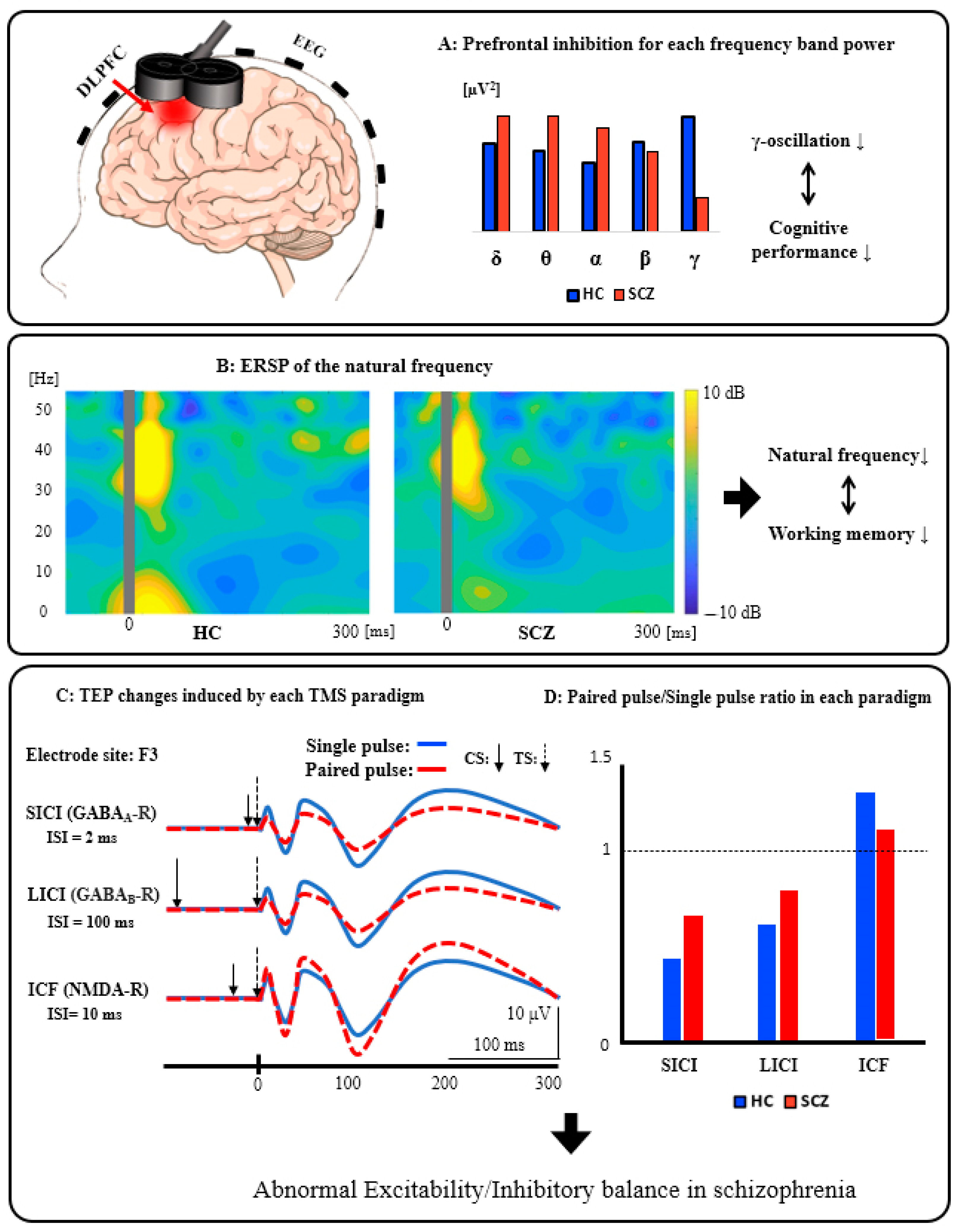
3.2. Schematic Summary of the TMS-EEG Findings from the DLPFC in Patients with SCZ
3.3. TMS-Evoked Potentials (TEP) Analyses
3.3.1. Single-Pulse TMS Paradigm
3.3.2. Paired-Pulse TMS Paradigm
3.4. Time-Frequency Analyses
3.4.1. Single-Pulse TMS Paradigm
3.4.2. Paired-Pulse TMS Paradigm
3.5. Connectivity Analyses
3.5.1. Single-Pulse TMS Paradigm
3.5.2. Paired-Pulse TMS Paradigm
3.6. Risk of Bias
4. Discussion
4.1. Summary of This Review
4.2. Evidence to Support the E/I Imbalance Hypothesis in SCZ
4.3. Evidence to Support the GABA Hypothesis in SCZ
4.4. Evidence to Support the Glutamate Hypothesis in SCZ
4.5. Potential Evidence to Support the Cholinergic Hypothesis in SCZ
4.6. Abnormalities of TMS-Induced Gamma Oscillations in SCZ
4.7. Insights from This Systematic Review
4.8. Limitations of This Review and TMS-EEG Study in General
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Saha, S.; Chant, D.; Welham, J.; McGrath, J. A Systematic Review of the Prevalence of Schizophrenia. PLoS Med. 2005, 2, e141. [Google Scholar] [CrossRef]
- Radhu, N.; Ravindran, L.N.; Levinson, A.J.; Daskalakis, Z.J. Inhibition of the cortex using transcranial magnetic stimulation in psychiatric populations: Current and future directions. J. Psychiatry Neurosci. 2012, 37, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogasch, N.C.; Daskalakis, Z.J.; Fitzgerald, P.B. Cortical Inhibition, Excitation, and Connectivity in Schizophrenia: A Review of Insights from Transcranial Magnetic Stimulation. Schizophr. Bull. 2013, 40, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Van Os, J.; Kapur, S. Schizophrenia. Lancet 2009, 374, 635–645. [Google Scholar] [CrossRef]
- Coyle, J.T. Substance use disorders and schizophrenia: A question of shared glutamatergic mechanisms. Neurotox. Res. 2006, 10, 221–233. [Google Scholar] [CrossRef]
- Lewis, D.A.; Hashimoto, T.; Volk, D.W. Cortical inhibitory neurons and schizophrenia. Nat. Rev. Neurosci. 2005, 6, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y.; Coyle, J.T. Glutamate hypothesis in schizophrenia. Psychiatry Clin. Neurosci. 2019, 73, 204–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, J.D.; Anticevic, A.; Gancsos, M.; Ichinose, M.; Corlett, P.R.; Krystal, J.H.; Wang, X.-J. Linking Microcircuit Dysfunction to Cognitive Impairment: Effects of Disinhibition Associated with Schizophrenia in a Cortical Working Memory Model. Cereb. Cortex 2012, 24, 859–872. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.G.; Williams, G.V.; Goldman-Rakic, P.S. Destruction and creation of spatial tuning by disinhibition: GABA(A) blockade of prefrontal cortical neurons engaged by working memory. J. Neurosci. 2000, 20, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foss-Feig, J.H.; Adkinson, B.D.; Ji, J.L.; Yang, G.; Srihari, V.H.; McPartland, J.C.; Krystal, J.H.; Murray, J.D.; Anticevic, A. Searching for Cross-Diagnostic Convergence: Neural Mechanisms Governing Excitation and Inhibition Balance in Schizophrenia and Autism Spectrum Disorders. Biol. Psychiatry 2017, 81, 848–861. [Google Scholar] [CrossRef] [Green Version]
- Canitano, R.; Pallagrosi, M. Autism Spectrum Disorders and Schizophrenia Spectrum Disorders: Excitation/Inhibition Imbalance and Developmental Trajectories. Front. Psychiatry 2017, 8, 69. [Google Scholar] [CrossRef]
- Marsman, A.; Mandl, R.C.; Klomp, D.W.; Bohlken, M.M.; Boer, V.O.; Andreychenko, A.; Cahn, W.; Kahn, R.S.; Luijten, P.R.; Pol, H.E.H. GABA and glutamate in schizophrenia: A 7 T 1H-MRS study. NeuroImage Clin. 2014, 6, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Vajawat, B.; Rao, N.P. Frontal GABA in schizophrenia: A meta-analysis of 1H-MRS studies. World J. Biol. Psychiatry 2020, 1–13. [Google Scholar] [CrossRef]
- Rowland, L.M.; Kontson, K.; West, J.; Edden, R.A.; Zhu, H.; Wijtenburg, S.A.; Holcomb, H.H.; Barker, P.B. In Vivo Measurements of Glutamate, GABA, and NAAG in Schizophrenia. Schizophr. Bull. 2012, 39, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Schür, R.R.; Draisma, L.W.; Wijnen, J.P.; Boks, M.P.; Koevoets, M.G.; Joëls, M.; Klomp, D.W.; Kahn, R.S.; Vinkers, C.H. Brain GABA levels across psychiatric disorders: A systematic literature review and meta-analysis of1H-MRS studies. Hum. Brain Mapp. 2016, 37, 3337–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egerton, A.; Modinos, G.; Ferrera, D.; McGuire, P. Neuroimaging studies of GABA in schizophrenia: A systematic review with meta-analysis. Transl. Psychiatry 2017, 7, e1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, Y. Toward the establishment of neurophysiological indicators for neuropsychiatric disorders using transcranial magnetic stimulation-evoked potentials: A systematic review. Psychiatry Clin. Neurosci. 2020, 74, 12–34. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.; Jalinous, R.; Freeston, I. Non-invasive magnetic stimulation of human motor cortex. Lancet 1985, 325, 1106–1107. [Google Scholar] [CrossRef]
- McClintock, S.M.; Freitas, C.; Oberman, L.; Lisanby, S.H.; Pascual-Leone, A. Transcranial Magnetic Stimulation: A Neuroscientific Probe of Cortical Function in Schizophrenia. Biol. Psychiatry 2011, 70, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Daskalakis, Z.J.; Farzan, F.; Radhu, N.; Fitzgerald, P.B. Combined transcranial magnetic stimulation and electroencephalography: Its past, present and future. Brain Res. 2012, 1463, 93–107. [Google Scholar] [CrossRef]
- Siebner, H.R.; Hartwigsen, G.; Kassuba, T.; Rothwell, J.C. How does transcranial magnetic stimulation modify neuronal activity in the brain? Implications for studies of cognition. Cortex 2009, 45, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafi, M.M.; Westover, M.B.; Fox, M.D.; Pascual-Leone, A. Exploration and modulation of brain network interactions with noninvasive brain stimulation in combination with neuroimaging. Eur. J. Neurosci. 2012, 35, 805–825. [Google Scholar] [CrossRef] [Green Version]
- Shafi, M.M.; Westover, M.B.; Oberman, L.; Cash, S.S.; Pascual-Leone, A. Modulation of EEG Functional Connectivity Networks in Subjects Undergoing Repetitive Transcranial Magnetic Stimulation. Brain Topogr. 2014, 27, 172–191. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.; Taylor, P.C.; Rushworth, M.F. Imaging causal interactions during sensorimotor processing. Cortex 2008, 44, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Ilić, T.V.; Meintzschel, F.; Cleff, U.; Ruge, D.; Kessler, K.R.; Ziemann, U. Short-interval paired-pulse inhibition and facilitation of human motor cortex: The dimension of stimulus intensity. J. Physiol. 2002, 545, 153–167. [Google Scholar] [CrossRef]
- Tamburin, S.; Fiaschi, A.; Marani, S.; Andreoli, A.; Manganotti, P.; Zanette, G. Enhanced intracortical inhibition in cerebellar patients. J. Neurol. Sci. 2004, 217, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Kujirai, T.; Caramia, M.D.; Rothwell, J.C.; Day, B.L.; Thompson, P.D.; Ferbert, A.; Wroe, S.; Asselman, P.; Marsden, C.D. Corticocortical inhibition in human motor cortex. J. Physiol. 1993, 471, 501–519. [Google Scholar] [CrossRef]
- Cash, R.F.H.; Noda, Y.; Zomorrodi, R.; Radhu, N.; Farzan, F.; Rajji, T.K.; Fitzgerald, P.B.; Chen, R.; Daskalakis, Z.J.; Blumberger, D.M. Characterization of Glutamatergic and GABAA-Mediated Neurotransmission in Motor and Dorsolateral Prefrontal Cortex Using Paired-Pulse TMS–EEG. Neuropsychopharmacology 2016, 42, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Di Lazzaro, V.; Oliviero, A.; Meglio, M.; Cioni, B.; Tamburrini, G.; Tonali, P.; Rothwell, J. Direct demonstration of the effect of lorazepam on the excitability of the human motor cortex. Clin. Neurophysiol. 2000, 111, 794–799. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Oliviero, A.; Tonali, P.A.; Marra, C.; Daniele, A.; Profice, P.; Saturno, E.; Pilato, F.; Masullo, C.; Rothwell, J.C. Noninvasive in vivo assessment of cholinergic cortical circuits in AD using transcranial magnetic stimulation. Neurology 2002, 59, 392–397. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Oliviero, A.; Saturno, E.; Dileone, M.; Pilato, F.; Nardone, R.; Ranieri, F.; Musumeci, G.; Fiorilla, T.; Tonali, P. Effects of lorazepam on short latency afferent inhibition and short latency intracortical inhibition in humans. J. Physiol. 2005, 564 Pt 2, 661–668. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Pilato, F.; Dileone, M.; Tonali, P.A.; Ziemann, U. Dissociated effects of diazepam and lorazepam on short-latency afferent inhibition. J. Physiol. 2005, 569 Pt 1, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Corwell, B.; Cohen, L.G. Modulation of Plasticity in Human Motor Cortex after Forearm Ischemic Nerve Block. J. Neurosci. 1998, 18, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Peurala, S.H.; Müller-Dahlhaus, J.F.M.; Arai, N.; Ziemann, U. Interference of short-interval intracortical inhibition (SICI) and short-interval intracortical facilitation (SICF). Clin. Neurophysiol. 2008, 119, 2291–2297. [Google Scholar] [CrossRef]
- McDonnell, M.N.; Orekhov, Y.; Ziemann, U. The role of GABA(B) receptors in intracortical inhibition in the human motor cortex. Exp. Brain Res. 2006, 173, 86–93. [Google Scholar] [CrossRef]
- Ziemann, U.; Reis, J.; Schwenkreis, P.; Rosanova, M.; Strafella, A.; Badawy, R.; Müller-Dahlhaus, F. TMS and drugs revisited 2014. Clin. Neurophysiol. 2015, 126, 1847–1868. [Google Scholar] [CrossRef] [PubMed]
- Schwenkreis, P.; Witscher, K.; Janssen, F.; Addo, A.; Dertwinkel, R.; Zenz, M.; Malin, J.-P.; Tegenthoff, M. Influence of the N-methyl-d-aspartate antagonist memantine on human motor cortex excitability. Neurosci. Lett. 1999, 270, 137–140. [Google Scholar] [CrossRef]
- Fujiki, M.; Hikawa, T.; Abe, T.; Ishii, K.; Kobayashi, H. Reduced short latency afferent inhibition in diffuse axonal injury patients with memory impairment. Neurosci. Lett. 2006, 405, 226–230. [Google Scholar] [CrossRef]
- Nardone, R.; Golaszewski, S.; Ladurner, G.; Tezzon, F.; Trinka, E. A Review of Transcranial Magnetic Stimulation in the in vivo Functional Evaluation of Central Cholinergic Circuits in Dementia. Dement. Geriatr. Cogn. Disord. 2011, 32, 18–25. [Google Scholar] [CrossRef]
- Cucurachi, L.; Immovilli, P.; Granella, F.; Pavesi, G.; Cattaneo, L. Short-latency afferent inhibition predicts verbal memory performance in patients with multiple sclerosis. J. Neurol. 2008, 255, 1949. [Google Scholar] [CrossRef]
- Noda, Y.; Barr, M.S.; Zomorrodi, R.; Cash, R.F.H.; Rajji, T.K.; Farzan, F.; Chen, R.; George, T.P.; Daskalakis, Z.J.; Blumberger, D.M. Reduced Short-Latency Afferent Inhibition in Prefrontal but not Motor Cortex and Its Association With Executive Function in Schizophrenia: A Combined TMS-EEG Study. Schizophr. Bull. 2017, 44, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Noda, Y.; Cash, R.F.H.; Zomorrodi, R.; Dominguez, L.G.; Farzan, F.; Rajji, T.K.; Barr, M.S.; Chen, R.; Daskalakis, Z.J.; Blumberger, D.M. A combined TMS-EEG study of short-latency afferent inhibition in the motor and dorsolateral prefrontal cortex. J. Neurophysiol. 2016, 116, 938–948. [Google Scholar] [CrossRef] [Green Version]
- Chorlian, D.B.; Rangaswamy, M.; Porjesz, B. EEG coherence: Topography and frequency structure. Exp. Brain Res. 2009, 198, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Rosanova, M.; Casali, A.; Bellina, V.; Resta, F.; Mariotti, M.; Massimini, M. Natural frequencies of human corticothalamic circuits. J. Neurosci. 2009, 29, 7679–7685. [Google Scholar] [CrossRef] [PubMed]
- Radhu, N.; de Jesus, D.R.; Ravindran, L.N.; Zanjani, A.; Fitzgerald, P.B.; Daskalakis, Z.J. A meta-analysis of cortical inhibition and excitability using transcranial magnetic stimulation in psychiatric disorders. Clin. Neurophysiol. 2013, 124, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Başar, E.; Güntekin, B. Review of delta, theta, alpha, beta, and gamma response oscillations in neuropsychiatric disorders. Suppl. Clin. Neurophysiol. 2013, 62, 303–341. [Google Scholar]
- Schmitt, A.; Hasan, A.; Gruber, O.; Falkai, P. Schizophrenia as a disorder of disconnectivity. Eur. Arch. Psychiatry Clin. Neurosci. 2011, 261 (Suppl. 2), 150–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vittala, A.; Murphy, N.; Maheshwari, A.; Krishnan, V. Understanding Cortical Dysfunction in Schizophrenia with TMS/EEG. Front. Neurosci. 2020, 14, 554. [Google Scholar] [CrossRef]
- Ferrarelli, F.; Riedner, B.A.; Peterson, M.J.; Tononi, G. Altered prefrontal activity and connectivity predict different cognitive deficits in schizophrenia. Hum. Brain Mapp. 2015, 36, 4539–4552. [Google Scholar] [CrossRef]
- Ferrarelli, F.; Sarasso, S.; Guller, Y.; Riedner, B.A.; Peterson, M.J.; Bellesi, M.; Massimini, M.; Postle, B.R.; Tononi, G. Reduced Natural Oscillatory Frequency of Frontal Thalamocortical Circuits in Schizophrenia. Arch. Gen. Psychiatry 2012, 69, 766–774. [Google Scholar] [CrossRef] [Green Version]
- Ferrarelli, F.; Kaskie, R.E.; Graziano, B.; Reis, C.C.; Casali, A.G. Abnormalities in the evoked frontal oscillatory activity of first-episode psychosis: A TMS/EEG study. Schizophr. Res. 2019, 206, 436–439. [Google Scholar] [CrossRef]
- Andrews, S.C.; Enticott, P.G.; Hoy, K.E.; Thomson, R.H.; Fitzgerald, P.B. No evidence for mirror system dysfunction in schizophrenia from a multimodal TMS/EEG study. Psychiatry Res. 2015, 228, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Frantseva, M.; Cui, J.; Farzan, F.; Chinta, L.V.; Velazquez, J.L.P.; Daskalakis, Z.J. Disrupted Cortical Conductivity in Schizophrenia: TMS-EEG Study. Cereb. Cortex 2012, 24, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzan, F.; Barr, M.S.; Levinson, A.J.; Chen, R.; Wong, W.; Fitzgerald, P.B.; Daskalakis, Z.J. Evidence for gamma inhibition deficits in the dorsolateral prefrontal cortex of patients with schizophrenia. Brain 2010, 133 Pt 5, 1505–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhu, N.; Dominguez, L.G.; Greenwood, T.A.; Farzan, F.; Semeralul, M.O.; Richter, M.A.; Kennedy, J.L.; Blumberger, D.M.; Chen, R.; Fitzgerald, P.B.; et al. Investigating Cortical Inhibition in First-Degree Relatives and Probands in Schizophrenia. Sci. Rep. 2017, 7, 43629. [Google Scholar] [CrossRef] [PubMed]
- Canali, P.; Sarasso, S.; Rosanova, M.; Casarotto, S.; Sferrazza-Papa, G.; Gosseries, O.; Fecchio, M.; Massimini, M.; Mariotti, M.; Cavallaro, R.; et al. Shared reduction of oscillatory natural frequencies in bipolar disorder, major depressive disorder and schizophrenia. J. Affect. Disord. 2015, 184, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Ferrarelli, F.; Massimini, M.; Peterson, M.J.; Riedner, B.A.; Lazar, M.; Murphy, M.J.; Huber, R.; Rosanova, M.; Alexander, A.L.; Kalin, N.; et al. Reduced Evoked Gamma Oscillations in the Frontal Cortex in Schizophrenia Patients: A TMS/EEG Study. Am. J. Psychiatry 2008, 165, 996–1005. [Google Scholar] [CrossRef]
- Noda, Y.; Barr, M.S.; Zomorrodi, R.; Cash, R.F.H.; Farzan, F.; Rajji, T.K.; Chen, R.; Daskalakis, Z.J.; Blumberger, D.M. Evaluation of short interval cortical inhibition and intracortical facilitation from the dorsolateral prefrontal cortex in patients with schizophrenia. Sci. Rep. 2017, 7, 17106. [Google Scholar] [CrossRef] [Green Version]
- Lett, T.A.; Kennedy, J.L.; Radhu, N.; Dominguez, L.G.; Chakravarty, M.M.; Nazeri, A.; Farzan, F.; Walter, H.; Heinz, A.; Mulsant, B.H.; et al. Prefrontal White Matter Structure Mediates the Influence of GAD1 on Working Memory. Neuropsychopharmacology 2016, 41, 2224–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhu, N.; Dominguez, L.G.; Farzan, F.; Richter, M.A.; Semeralul, M.O.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. Evidence for inhibitory deficits in the prefrontal cortex in schizophrenia. Brain 2015, 138 Pt 2, 483–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levit-Binnun, N.; Litvak, V.; Pratt, H.; Moses, E.; Zaroor, M.; Peled, A. Differences in TMS-evoked responses between schizophrenia patients and healthy controls can be observed without a dedicated EEG system. Clin. Neurophysiol. 2010, 121, 332–339. [Google Scholar] [CrossRef]
- Casali, A.G.; Casarotto, S.; Rosanova, M.; Mariotti, M.; Massimini, M. General indices to characterize the electrical response of the cerebral cortex to TMS. NeuroImage 2010, 49, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M. Grouping of brain rhythms in corticothalamic systems. Neuroscience 2006, 137, 1087–1106. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M.; Timofeev, I.; Dürmüller, N.; Grenier, F. Dynamic Properties of Corticothalamic Neurons and Local Cortical Interneurons Generating Fast Rhythmic (30–40 Hz) Spike Bursts. J. Neurophysiol. 1998, 79, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, P.B.; Brown, T.L.; Marston, N.A.; Oxley, T.J.; De Castella, A.; Daskalakis, Z.J.; Kulkarni, J. A transcranial magnetic stimulation study of abnormal cortical inhibition in schizophrenia. Psychiatry Res. 2003, 118, 197–207. [Google Scholar] [CrossRef]
- Tarumi, R.; Tsugawa, S.; Noda, Y.; Plitman, E.; Honda, S.; Matsushita, K.; Chavez, S.; Sawada, K.; Wada, M.; Matsui, M.; et al. Levels of glutamatergic neurometabolites in patients with severe treatment-resistant schizophrenia: A proton magnetic resonance spectroscopy study. Neuropsychopharmacology 2020, 45, 632–640. [Google Scholar] [CrossRef]
- Iwata, M.; Ota, K.T.; Li, X.-Y.; Sakaue, F.; Li, N.; Dutheil, S.; Banasr, M.; Duric, V.; Yamanashi, T.; Kaneko, K.; et al. Psychological Stress Activates the Inflammasome via Release of Adenosine Triphosphate and Stimulation of the Purinergic Type 2X7 Receptor. Biol. Psychiatry 2016, 80, 12–22. [Google Scholar] [CrossRef]
- Davenport, E.C.; Szulc, B.R.; Drew, J.; Taylor, J.; Morgan, T.; Higgs, N.F.; López-Doménech, G.; Kittler, J.T. Autism and Schizophrenia-Associated CYFIP1 Regulates the Balance of Synaptic Excitation and Inhibition. Cell Rep. 2019, 26, 2037–2051.e6. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Curr. Mol. Med. 2015, 15, 146–167. [Google Scholar] [CrossRef]
- Nair, P.C.; McKinnon, R.A.; Miners, J.O.; Bastiampillai, T. Binding of clozapine to the GABAB receptor: Clinical and structural insights. Mol. Psychiatry 2020, 25, 1910–1919. [Google Scholar] [CrossRef]
- Wu, Y.; Blichowski, M.; Daskalakis, Z.J.; Wu, Z.; Liu, C.C.; Cortez, M.A.; Snead, O.C. Evidence that clozapine directly interacts on the GABAB receptor. NeuroReport 2011, 22, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Kaster, T.S.; de Jesus, D.; Radhu, N.; Farzan, F.; Blumberger, D.M.; Rajji, T.K.; Fitzgerald, P.B.; Daskalakis, Z.J. Clozapine potentiation of GABA mediated cortical inhibition in treatment resistant schizophrenia. Schizophr. Res. 2015, 165, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Benes, F.M.; Berretta, S. GABAergic interneurons: Implications for understanding schizophrenia and bipolar disorder. Neuropsychopharmacology 2001, 25, 1–27. [Google Scholar] [CrossRef]
- Krystal, J.H.; D’Souza, D.C.; Mathalon, D.; Perry, E.; Belger, A.; Hoffman, R. NMDA receptor antagonist effects, cortical glutamatergic function, and schizophrenia: Toward a paradigm shift in medication development. Psychopharmacology 2003, 169, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Kegeles, L.S.; Mao, X.; Stanford, A.D.; Girgis, R.; Ojeil, N.; Xu, X.; Gil, R.; Slifstein, M.; Abi-Dargham, A.; Lisanby, S.H.; et al. Elevated prefrontal cortex gamma-aminobutyric acid and glutamate-glutamine levels in schizophrenia measured in vivo with proton magnetic resonance spectroscopy. Arch. Gen. Psychiatry 2012, 69, 449–459. [Google Scholar] [PubMed] [Green Version]
- Tani, M.; Akashi, N.; Hori, K.; Konishi, K.; Kitajima, Y.; Tomioka, H.; Inamoto, A.; Hirata, A.; Tomita, A.; Koganemaru, T.; et al. Anticholinergic Activity and Schizophrenia. Neurodegener. Dis. 2015, 15, 168–174. [Google Scholar] [CrossRef]
- Ter Braack, E.M.; de Vos, C.C.; van Putten, M.J. Masking the Auditory Evoked Potential in TMS-EEG: A Comparison of Various Methods. Brain Topogr. 2015, 28, 520–528. [Google Scholar] [CrossRef]
- Terry, A.V.; Callahan, P.M. α7 nicotinic acetylcholine receptors as therapeutic targets in schizophrenia: Update on animal and clinical studies and strategies for the future. Neuropharmacology 2020, 170, 108053. [Google Scholar] [CrossRef]
- Guan, Z.-Z.; Zhang, X.; Blennow, K.; Nordberg, A. Decreased protein level of nicotinic receptor α7 subunit in the frontal cortex from schizophrenic brain. Neuroreport 1999, 10, 1779–1782. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.F.; Kem, W.R.; Freedman, R. Alpha-7 nicotinic receptor agonists: Potential new candidates for the treatment of schizophrenia. Psychopharmacology 2004, 174, 54–64. [Google Scholar] [CrossRef]
- Volman, V.; Behrens, M.M.; Sejnowski, T.J. Downregulation of Parvalbumin at Cortical GABA Synapses Reduces Network Gamma Oscillatory Activity. J. Neurosci. 2011, 31, 18137–18148. [Google Scholar] [CrossRef] [PubMed]
- Farzan, F.; Barr, M.S.; Wong, W.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. Suppression of gamma-oscillations in the dorsolateral prefrontal cortex following long interval cortical inhibition: A TMS-EEG study. Neuropsychopharmacology 2009, 34, 1543–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzan, F.; Barr, M.S.; Sun, Y.; Fitzgerald, P.B.; Daskalakis, Z.J. Transcranial magnetic stimulation on the modulation of gamma oscillations in schizophrenia. Ann. N. Y. Acad. Sci. 2012, 1265, 25–35. [Google Scholar] [CrossRef] [PubMed]
- McNally, J.M.; McCarley, R.W. Gamma band oscillations: A key to understanding schizophrenia symptoms and neural circuit abnormalities. Curr. Opin. Psychiatry 2016, 29, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Senkowski, D.; Gallinat, J. Dysfunctional Prefrontal Gamma-Band Oscillations Reflect Working Memory and Other Cognitive Deficits in Schizophrenia. Biol. Psychiatry 2015, 77, 1010–1019. [Google Scholar] [CrossRef]
- Brignani, D.; Manganotti, P.; Rossini, P.M.; Miniussi, C. Modulation of cortical oscillatory activity during transcranial magnetic stimulation. Hum. Brain Mapp. 2008, 29, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Veniero, D.; Brignani, D.; Thut, G.; Miniussi, C. Alpha-generation as basic response-signature to transcranial magnetic stimulation (TMS) targeting the human resting motor cortex: A TMS/EEG co-registration study. Psychophysiology 2011, 48, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Whitham, E.M.; Pope, K.J.; Fitzgibbon, S.P.; Lewis, T.; Clark, C.R.; Loveless, S.; Broberg, M.; Wallace, A.; DeLosAngeles, D.; Lillie, P.; et al. Scalp electrical recording during paralysis: Quantitative evidence that EEG frequencies above 20Hz are contaminated by EMG. Clin. Neurophysiol. 2007, 118, 1877–1888. [Google Scholar] [CrossRef]
- Kaskie, R.E.; Ferrarelli, F. Investigating the neurobiology of schizophrenia and other major psychiatric disorders with Transcranial Magnetic Stimulation. Schizophr. Res. 2018, 192, 30–38. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| (A) | Single-Pulse TMS Paradigm | ||||||||||||
| Authors, Year | Patient Group | Age (Mean ± SD) | Number of Subjects (Female) | Clinical Severity (Mean ± SD) | Medication | Control Group | Age (Mean ± SD) | Number of Subjects (Female) | Stimulation Parameters | Areas of Stimulation | Cognitive Measures | Analyses | Neurophysiological Findings |
| Levit-Binnun et al., 2009 | SCZ | 38 ± 8 | 8(0) | PANSS | Five patients were on atypical antipsychotics (ziprasidone) and two were on typical antipsychotics (haloperidol and fluphenazine: mean dose equivalent of 415 mg chlorpromazine). | HC | 29 ± 10 | 6(3) | Single-pulse TMS Sham | Over the Cz electrode | N/A | TMS-evoked potential (TEP) analysis Amplitude Latency | In HCs, clear frontal negativity and parietal positivity were observed at 29 ms after TMS stimulation, but in SCZ, there was no frontal negativity and parietal positivity was greatly reduced. |
| Noda et al., 2018 | Chronic SCZ | 41 ± 10 | 12(4) | PANSS total: 50 ± 6.2 PANSS negative: 12 ± 3.4 PANSS positive: 11 ± 3.0 PANSS general: 24 ± 2.8 | Patients were taking chlorpromazine equivalent dose (330 ± 290 mg/day) of antipsychotics. | HC | 39 ± 12 | 12(6) | Short-latency afferent inhibition (SAI) | Left primary motor cortex (M1) Dorsolateral prefrontal cortex (DLPFC) | Wechsler Test of Adult Reading Letter-Number Span Test Hopkins Verbal Learning Test Trail Making Test | TMS-evoked potential (TEP) analysis Amplitude of components | Patients with SCZ had significantly smaller modulation of N100 by DLPFC-SAI compared to HC, suggesting impaired cholinergic neurophysiological function in DLPFC. Furthermore, reduced DLPFC-SAI correlated with executive dysfunction as measured by TMT. |
| (B) | Paired-Pulse TMS Paradigm | ||||||||||||
| Authors, Year | Patient Group | Age (Mean ± SD) | Number of Subjects (Female) | Clinical Severity (Mean ± SD) | Medication | Control Group | Age (Mean ± SD) | Number of Subjects (Female) | Stimulation Parameters | Areas of Stimulation | Cognitive/Clinical Measures | Analyses | Neurophysiological Findings |
| Noda et al., 2017 | SCZ | 41 ± 10 | 12(4) | PANSS total: 50 ± 6.2 PANSS negative: 12 ± 3.4 PANSS positive: 11 ± 3.0 PANSS general: 24 ± 2.8 | Patients were taking chlorpromazine equivalent dose (330 ± 290 mg/day) of antipsychotics. | HC | 39 ± 12 | 12(6) | Short interval intracortical inhibition (SICI) Intracortical facilitation (ICF) | Left dorsolateral prefrontal cortex (DLPFC) | Wechsler Test of Adult Reading Letter-Number Span Test Trail Making Test Hopkins Verbal Learning Test | TMS-evoked potential (TEP) analysis Amplitude of components Frequency band powers Time-frequency analysis | Patients with SCZ showed reduced inhibition in TEP P60 by DLPFC-SICI compared with HC, which was correlated with the longest span of the LNST. Further, patients with SCZ showed reduced facilitation in TEP P60 and N100 by DLPFC-ICF compared with HC, which were correlated with the total score of the PANSS. |
| (A) | Single-Pulse TMS Paradigm | ||||||||||||
| Authors, Year | Patient Group | Age (Mean ± SD) | Number of Subjects (Female) | Clinical Severity (Mean ± SD) | Medication | Control Group | Age (Mean ± SD) | Number of Subjects (Female) | Stimulation Parameters | Areas of Stimulation | Cognitive Measures | Analyses | Neurophysiological Findings |
| Ferrarelli et al., 2012 | SCZ | 33 ± 6.2 | 20(7) | PANSS general: 39 ± 11 PANSS negative: 22 ± 6.0 PANSS positive: 18 ± 6.3 | Eighteen patients were taking second-generation antipsychotics while two were on first-generation antipsychotics. | HC | 32 ± 7.8 | 20(4) | Single-pulse | Posterior parietal cortex Motor cortex Premotor cortex Prefrontal cortex | The word memory Penn Word Recognition Test The facial memory Penn Facial Memory Test | Time-Frequency Analysis Event-related spectra perturbation (ERSP) Intertrial coherence (ITC) | In patients with SCZ, the natural frequency response was generally attenuated compared with HC when single-pulse TMS was applied to the prefrontal cortex. Further, the lowered natural frequency in the prefrontal areas in SCZ was related to the PANSS positive scores and reaction time in the word memory task. |
| Frantseva et al., 2014 | SCZ or schizoaffective disorder | 37 ± 10 | 16(4) | PANSS total: 65 ± 18 PANSS negative: 18 ± 6.1 PANSS positive: 16 ± 4.3 PANSS global: 30 ± 8.6 | Fourteen patients with schizophrenia were treated with antipsychotic medications (clozapine: n = 6, mean dose 400 ± 55 mg/day; risperidone: n = 3, mean dose 3.2 ± 2.5 mg; haloperidol: n = 2, mean dose 2.0 ± 1.4 mg; quetiapine: n = 1, 100 mg; perphenazine: n = 1, 16 mg; olanzapine: n = 1, 7.5 mg) and two patients did not take any psychotropic medications. | HC | 36 ± 7.9 | 16(5) | Single-pulse (single monophasic TMS pulse) Sham | Left motor cortex | N/A | Time-domain Analysis Frequency-domain Analysis Time-Frequency Analysis | TMS-induced cortical activation in the gamma band between 400 and 700 ms at the M1 was positively correlated with positive symptoms in patients with SCZ. In contrast, the activation in theta and delta bands at 200 ms after TMS was positively correlated with negative symptoms in patients with SCZ. |
| Canali et al., 2015 | Chronic undifferentiated SCZ | 38 ± 9 | 12(3) | PANSS general: 37 ± 5 PANSS negative: 18 ± 4 PANSS positive: 18 ± 4 | All patients were taking antipsychotics (typical antipsychotics: n = 5; atypical antipsychotics: n = 7). | HC | 39 ± 15 | 12(7) | Single-pulse | Premotor cortex | N/A | Time-Frequency analysis Event-related spectral perturbation (ERSP) | Natural frequency in the frontal cortex was significantly slower in patients with bipolar disorder (20 ± 3.7 Hz), major depressive disorder (19 ± 5.0 Hz), and SCZ (20 ± 4.2 Hz) than HC (27 ± 3.2 Hz). However, frontal natural frequencies among the patient groups (i.e., bipolar disorder, major depressive disorder, and SCZ) were not significantly different. There was no correlation among natural frequency in the frontal area, PANSS scores, and medication dose in these populations. |
| Ferrarelli et al., 2019 | First-episode psychosis(FEP) | 23 ± 5.2 | 16(4) | Scale for the assessment of positive symptoms (SAPS): 18 ± 13 Scale for the assessment of negative symptoms scores (SANS): 31 ± 12 | Nine FEP patients were antipsychotic naïve and seven patients were taking antipsychotics less than 1 month. | HC | 23 ± 6.3 | 11(3) | Single-pulse | Left primary motor cortex (M1) | N/A | Time-Frequency analysis Time domain: The global mean field power (GMFP) Frequency domain: Relative spectral power (RSP)/Cumulated RSP (cRSP) | GMFP for the time domain was not significantly different between patients with FEP and HC. When RSP was assessed for the frequency domain, patients with FEP showed a significantly decreased beta/low-gamma TEP activities at the fronto-central area relative to HC. The lower RSP was associated with both worse scores on the SAPS and the SANS. TMS-evoked fast oscillations over the fronto-central areas were impaired from the time of onset, suggesting that these deficits may be related to the clinical symptoms. |
| Andrews et al., 2015 | SCZ or schizoaffective disorder | 44 ± 11 | 19 | PANSS general: 34 ± 8.1 PANSS negative: 16 ± 5.5 PANSS positive: 15 ± 6.1 | Patients were taking chlorpromazine equivalent dose (67~1307 mg/day) of antipsychotics. | HC | 38 ± 13 | 19 | Single-pulse TMS during the observation of hand movements designed to elicit mirror system activity | Primary motor cortex (M1) | NimStim Static Face Task Cognitive and Affective Mental Inference Task (Inference Task) | Frequency analysis mu rhythm (8–13 Hz) TMS induced motor-evoked potentials (MEPs) | Patients with SCZ showed lower accuracy on the facial affect recognition and theory of mind tasks than HC. No significant differences in the degree of mu suppression and motor resonance between the patients with SCZ and HC. |
| (B) | Paired-Pulse TMS Paradigm | ||||||||||||
| Authors, Year | Patient Group | Age (Mean ± SD) | Number of Subjects (Female) | Clinical Severity (Mean ± SD) | Medication | Control Group | Age (Mean ± SD) | Number of Subjects (Female) | Stimulation Parameters | Areas of Stimulation | Cognitive Measures | Analyses | Neurophysiological Findings |
| Radhu et al., 2015 | SCZ | 36 | 38(13) | Brief Psychiatric Rating Scale (BPRS) | All patients were treated with antipsychotics. | HC | 34 | 46(23) | Long-Interval Cortical Inhibition (LICI) | Left motor cortex Dorsolateral prefrontal cortex (DLPFC) | N/A | Time-Frequency analysis | LICI was significantly reduced in patients with SCZ compared with other groups in the DLPFC not in M1. |
| Lett et al., 2016 | SCZ or schizoaffective disorder | 35 ± 10 | 23(5) | N/A | Not reported | HC | 35 ± 11 | 33(18) | Long-Interval Cortical Inhibition (LICI) | Dorsolateral prefrontal cortex (DLPFC) | IQ Wechsler Test of Adult ReadingWorking memory Letter-number sequencing task Digit-span forward taskSelective attention Stroop Neuropsychological Screening Test | Time-Frequency analysis Cluster-Based analysis | Variation of the GAD1 gene in patients with SCZ may play a pivotal role in GABA(B)ergic inhibitory neurotransmission and working memory performance in the DLPFC. |
| (A) | Single-Pulse TMS Paradigm | ||||||||||||
| Authors, Year | Patient Group | Age (Mean ± SD) | Number of Subjects (Female) | Clinical Severity (Mean ± SD) | Medication | Control Group | Age (Mean ± SD) | Number of Subjects (female) | Stimulation Parameters | Areas of Stimulation | Cognitive Measures | Analyses | Neurophysiological Findings |
| Ferrarelli et al., 2015 | SCZ | 33 ± 6.2 | 20(7) | PANSS general: 39 ± 11 PANSS negative: 22 ± 5.8 PANSS positive: 18 ± 6.3 | Patients were taking chlorpromazine equivalent dose (314 ± 129 mg/day) of antipsychotics. | HC | 32 ± 7.8 | 20(4) | Single-pulse TMS | Prefrontal cortex (PFC) Premotor cortex Motor cortex Parietal cortex | Episodic Memory Word Memory Delayed Executive Function Penn Conditional Exclusion Test | TMS-evoked potential (TEP) analysis Significant current density (SCD) Connectivity analysis Significant current scattering (SCS) | Both SCD and SCS were most impaired in the DLPFC after single-pulse TMS in patients with SCZ compared with HC. No difference in SCD and SCS were observed in the parietal cortex and M1 after single-pulse TMS. SCD and performance in episodic memory were negatively correlated, whereas higher SCS values were associated with a lower executive function. |
| Ferrarelli et al., 2008 | SCZ | 34 ± 8.0 | 16(3) | PANSS | N/A | HC | 35 ± 7.0 | 14(3) | Single-pulse TMS | Right premotor cortex | N/A | TMS-evoked potential (TEP) analysis Amplitude The global mean field power (GMFP) Event-related spectral perturbation phase Connectivity analysis | Patients with SCZ indicated significantly decreased amplitude and synchronization of TMS-evoked gamma oscillations particularly in the frontocentral region during the 100 ms after TMS pulse compared with HC. In the source modeling analysis, cortical propagation of TMS-evoked gamma oscillations was more localized compared with HC. |
| (B) | Paired-Pulse TMS Paradigm | ||||||||||||
| Authors, Year | Patient Group | Age (Mean ± SD) | Number of Subjects (Female) | Clinical Severity (Mean ± SD) | Medication | Control Group | Age (Mean ± SD) | Number of Subjects (Female) | Stimulation Parameters | Areas of Stimulation | Cognitive Measures | Analyses | Neurophysiological Findings |
| Farzan et al., 2010 | SCZ | 38 ± 10 | 14(4) | PANSS total: 66 ± 18 PANSS negative: 18 ± 6.3 PANSS positive: 17 ± 4.4 PANSS global: 31 ± 9.2 | Two patients were unmedicated (one medication-naive; one medication-free for 6 months) and 12 patients were on medication (n = 5, 390.0 ± 54.8 mg clozapine; n = 3, 3.2 ± 2.5 mg risperidone; n = 2, 2 ± 1.4 mg haloperidol; n-1, 100 mg of quetiapine; n = 1, 16 mg perphenazine) | HC | 37 ± 7.6 | 14(5) | Long-Interval Cortical Inhibition (LICI) Sham | Left motor cortex Dorsolateral prefrontal cortex (DLPFC) | N/A | Time-Frequency analysis | Patients with SCZ had significant deficits of cortical inhibition and inhibitory modulation of gamma oscillations in the DLPFC but not in M1 compared with the other groups. |
| Radhu et al., 2017 | (a) SCZ or schizoaffective disorder (b) First-degree relatives of patients with SCZ | (a) 30 (b) 54 | (a) 19(9) (b) 30(17) | Schizotypal Personality Questionnaire The 24-construct Brief Psychiatric Rating Scale | Patients were taking clozapine (150~475 mg/day). | HC | 33 | 49(25) | Long-Interval Cortical Inhibition (LICI) | Motor cortex Dorsolateral prefrontal cortex (DLPFC) | N/A | Time-Frequency analysis | The degree of cortical inhibition as indexed by LICI was significantly decreased in patients with SCZ compared to HC. Further, no significant difference in the degree of cortical inhibition between HC and first-degree relatives of patients with SCZ. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Honda, S.; Nakajima, S.; Wada, M.; Yoshida, K.; Daskalakis, Z.J.; Mimura, M.; Noda, Y. TMS-EEG Research to Elucidate the Pathophysiological Neural Bases in Patients with Schizophrenia: A Systematic Review. J. Pers. Med. 2021, 11, 388. https://doi.org/10.3390/jpm11050388
Li X, Honda S, Nakajima S, Wada M, Yoshida K, Daskalakis ZJ, Mimura M, Noda Y. TMS-EEG Research to Elucidate the Pathophysiological Neural Bases in Patients with Schizophrenia: A Systematic Review. Journal of Personalized Medicine. 2021; 11(5):388. https://doi.org/10.3390/jpm11050388
Chicago/Turabian StyleLi, Xuemei, Shiori Honda, Shinichiro Nakajima, Masataka Wada, Kazunari Yoshida, Zafiris J. Daskalakis, Masaru Mimura, and Yoshihiro Noda. 2021. "TMS-EEG Research to Elucidate the Pathophysiological Neural Bases in Patients with Schizophrenia: A Systematic Review" Journal of Personalized Medicine 11, no. 5: 388. https://doi.org/10.3390/jpm11050388
APA StyleLi, X., Honda, S., Nakajima, S., Wada, M., Yoshida, K., Daskalakis, Z. J., Mimura, M., & Noda, Y. (2021). TMS-EEG Research to Elucidate the Pathophysiological Neural Bases in Patients with Schizophrenia: A Systematic Review. Journal of Personalized Medicine, 11(5), 388. https://doi.org/10.3390/jpm11050388