
Personalized Medicine Based on Nasal Epithelial Cells: Comparative Studies with Rectal Biopsies and Intestinal Organoids


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cohort and Ethical Approval
2.2. Rectal Biopsies
2.3. Primary Airway Cell Cultures
2.4. Ussing Chamber Measurements
2.5. Human Intestinal Organoids Culture
2.6. Forskolin-Induced Swelling (FIS) Assay
2.7. Statistical Analyses
3. Results
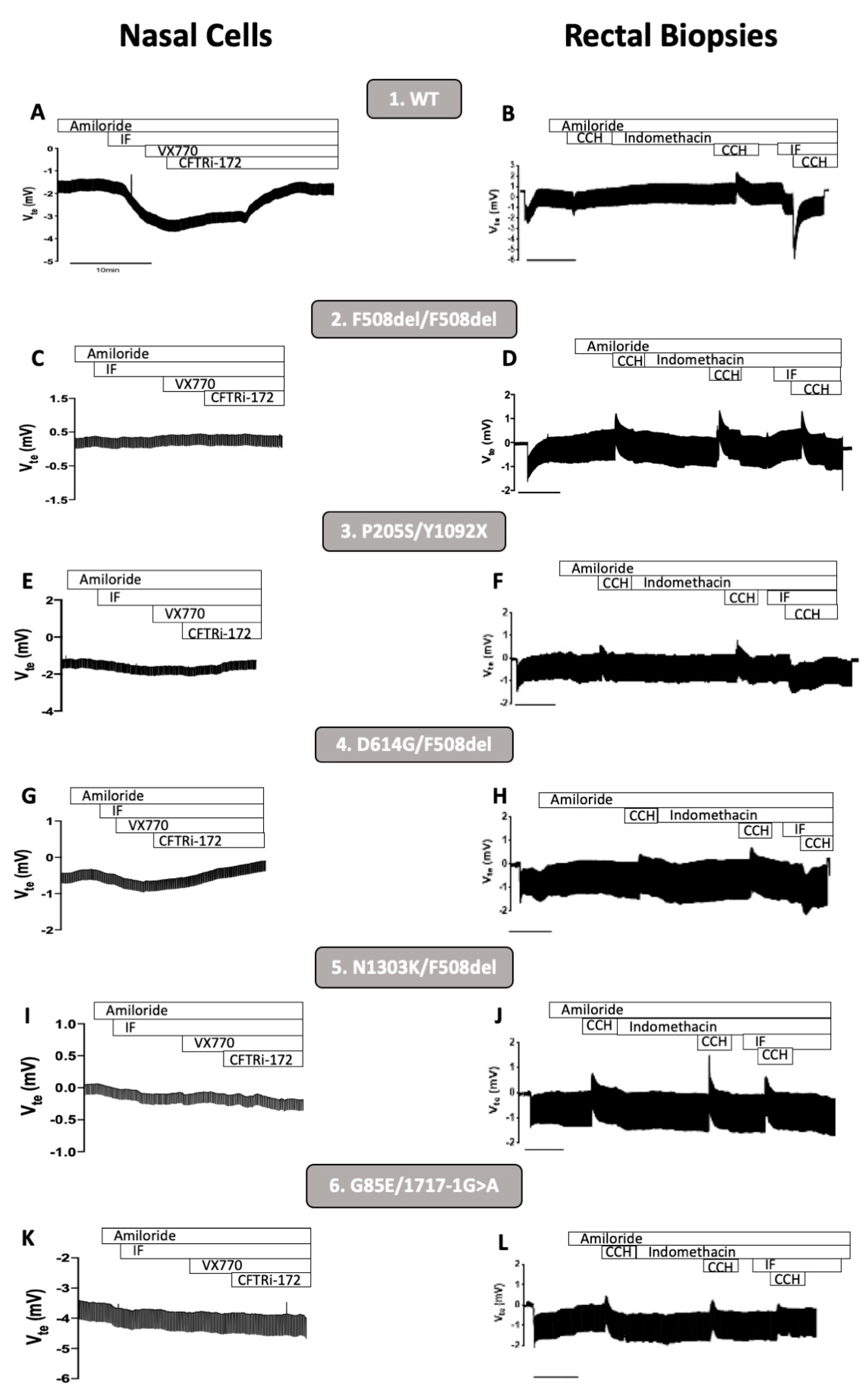
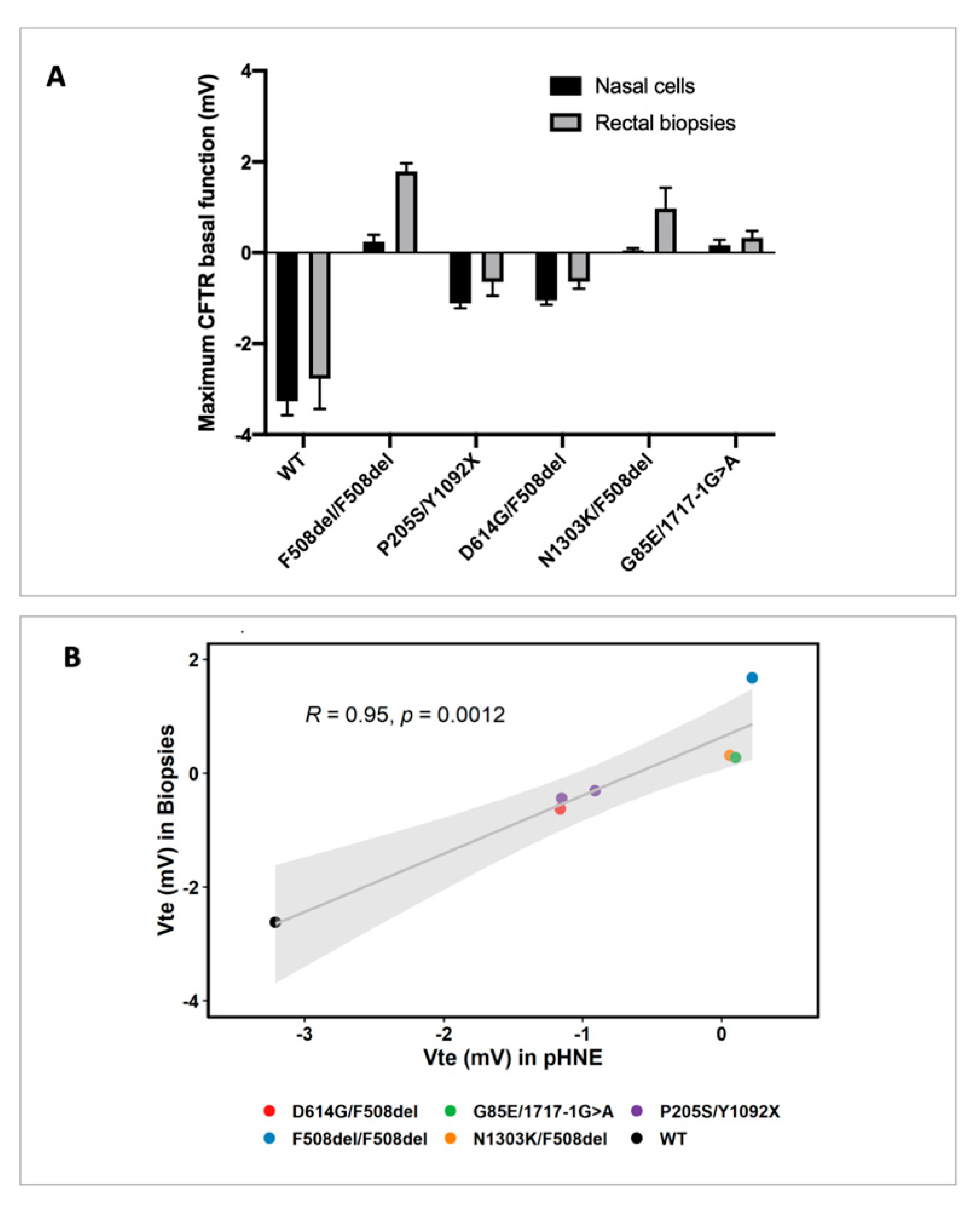
3.1. CFTR Basal Function in Primary Human Nasal Epithelial (pHNE) Cells Correlates with the One Measured in Rectal Biopsies
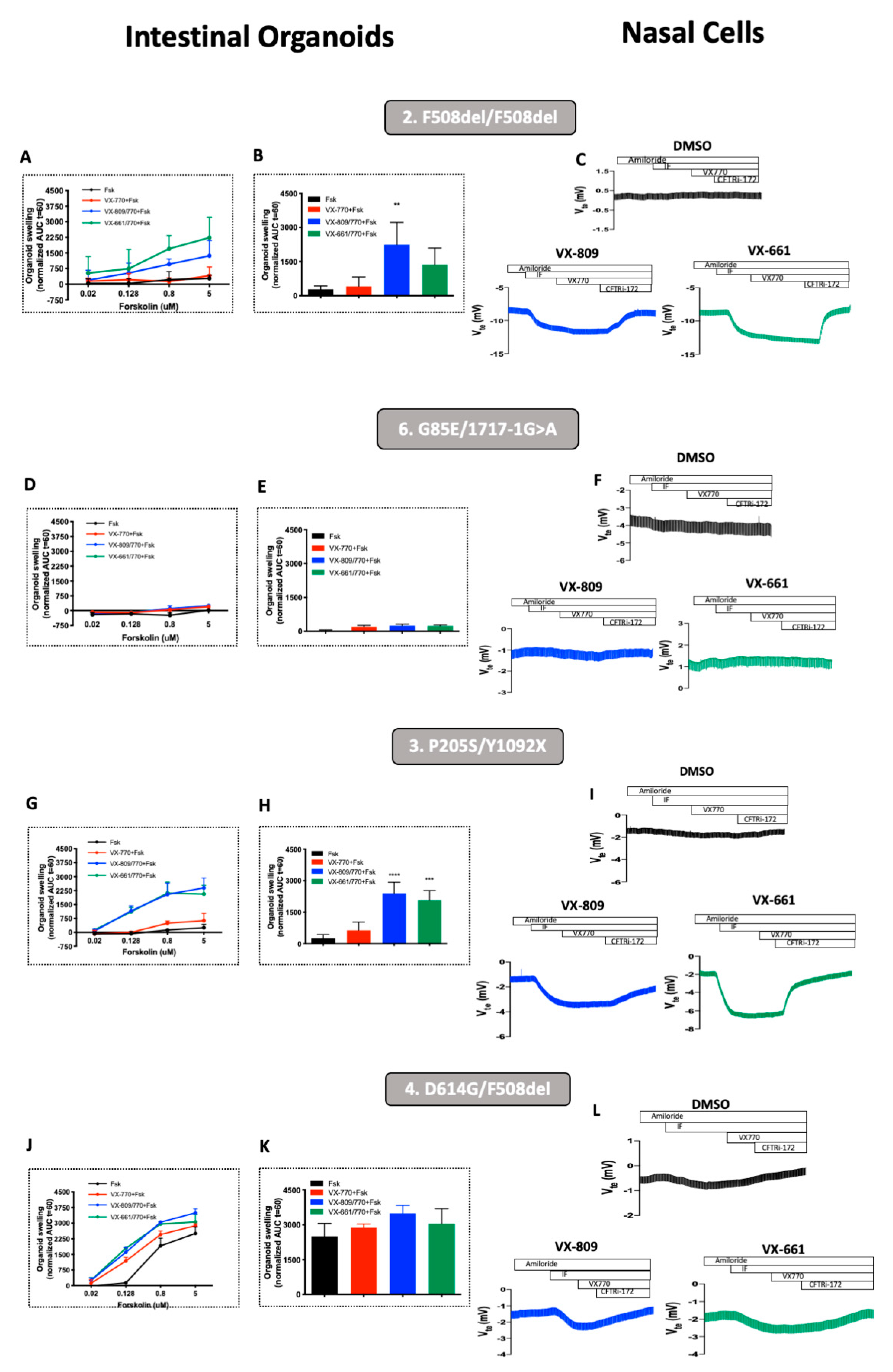
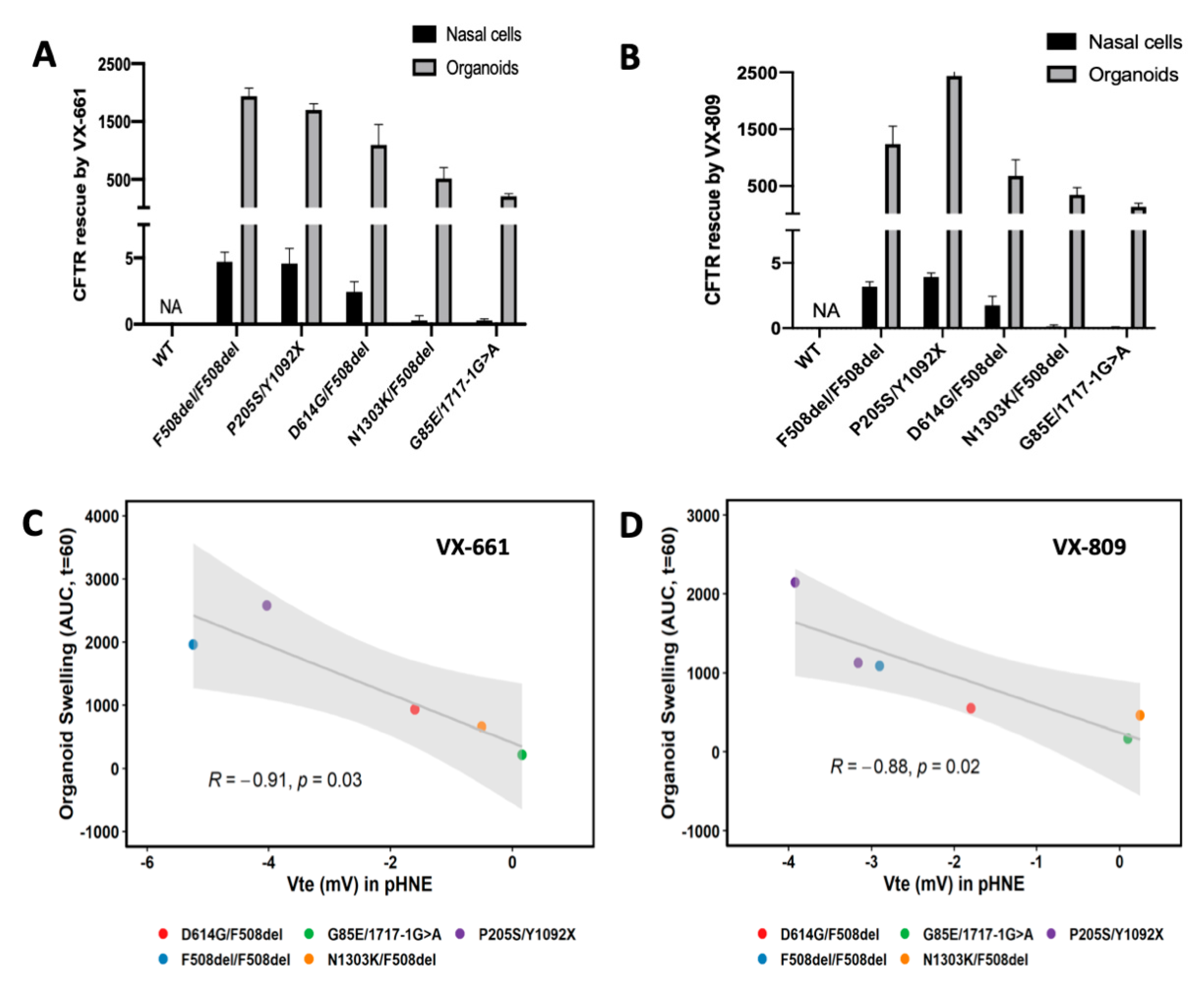
3.2. Rescue of CFTR-Mediated Cl− Secretion by CFTR Modulators pHNEs Correlate with CFTR Responses Measured in Intestinal Organoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | area under the curve |
| CCH | Carbachol |
| CF | Cystic Fibrosis |
| CFTR | Cystic Fibrosis Transmembrane Conductance Regulator |
| Cl− | chloride |
| ENaC | Epithelial Sodium Channel |
| Fsk | forskolin |
| FIS | forskolin induced swelling assay |
| HCO3− | bicarbonate |
| IBMX | 3-Isobutyl-1-methylxanthine |
| pHNE | primary human nasal epithelial cells |
| pHBE | primary human bronchial epithelial cells |
| PwCF | people with CF |
| Vte | Voltage |
References
- De Boeck, K.; Amaral, M.D. Progress in therapies for cystic fibrosis. Lancet Respir. 2016, 2600, 1–13. [Google Scholar] [CrossRef]
- Elborn, J.S.; Davies, J.; Mall, M.A.; Flume, P.A.; Plant, B. Current strategies for the long-term assessment, monitoring, and management of cystic fi brosis patients treated with CFTR modulator therapy. J. Cyst. Fibros. 2016, 16, 11–12. [Google Scholar]
- Lopes-Pacheco, M. CFTR Modulators: The Changing Face of Cystic Fibrosis in the Era of Precision Medicine. Front. Pharmacol. 2020, 10, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbertson, L.; Walker, A.W.; Oliver, A.E.; Rogers, G.B.; Rivett, D.W.; Hampton, T.H.; Ashare, A.; Elborn, J.S.; De Soyza, A.; Carroll, M.P.; et al. Lung function and microbiota diversity in cystic fibrosis. Microbiome 2020, 8, 1–13. [Google Scholar] [CrossRef]
- Cantin, A.M.; Hartl, D.; Konstan, M.W.; Chmiel, J.F. Inflammation in cystic fibrosis lung disease: Pathogenesis and therapy. J. Cyst. Fibros. 2015, 14, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, A.; Waters, V. Microbiology of Cystic Fibrosis Airway Disease. Semin. Respir. Crit. Care Med. 2019, 40, 727–736. [Google Scholar] [CrossRef]
- Cystic Fibrosis Mutation Database. Available online: http://www.genet.sickkids.on.ca (accessed on 20 April 2021).
- Middleton, P.G.; Hospital, W.; Mall, M.A.; Republic, C.; Lands, L.C.; Medicine, P.R.; Cystic, P.; Clinic, F.; Mckone, E.F.; Polineni, D.; et al. Elexacaftor–Tezacaftor–Ivacaftor for Cystic Fibrosis with a Single Phe508del Allele. N. Engl. J. Med. 2019, 381, 1809–1819. [Google Scholar] [CrossRef]
- Pranke, I.; Golec, A.; Hinzpeter, A.; Edelman, A.; Sermet-Gaudelus, I. Emerging therapeutic approaches for cystic fibrosis. From gene editing to personalized medicine. Front. Pharmacol. 2019, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Ridley, K.; Condren, M. Elexacaftor-tezacaftor-ivacaftor: The first triple-combination cystic fibrosis transmembrane conductance regulator modulating therapy. J. Pediatr. Pharmacol. Ther. 2020, 25, 192–197. [Google Scholar] [CrossRef]
- Harutyunyan, M.; Huang, Y.; Mun, F.; Yang, F.; Arora, K.; Naren, A. Personalized medicine in CF: From modulator development to therapy for cystic fibrosis patients with rare CFTR mutations. Am. J. Physiol. Lung. Cell Mol. Physiol. 2018, 314, L529–L543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skov, M.; Hansen, C.R.; Pressler, T. Cystic fibrosis—An example of personalized and precision medicine. Apmis 2019, 127, 352–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elborn, S.; Vallieres, E. Cystic fibrosis gene mutations: Evaluation and assessment of disease severity. Adv. Genom. Genet. 2014, 4, 161. [Google Scholar] [CrossRef] [Green Version]
- Mou, H.; Brazauskas, K.; Rajagopal, J. Personalized medicine for cystic fibrosis: Establishing human model systems. Pediatr. Pulmonol. 2015, 50, S14–S23. [Google Scholar] [CrossRef]
- Marson, F.A.L.; Bertuzzo, C.S.; Ribeiro, J.D. Personalized or precision medicine? The example of cystic fibrosis. Front. Pharmacol. 2017, 8, 390. [Google Scholar] [CrossRef] [PubMed]
- Awatade, N.T.; Uliyakina, I.; Farinha, C.M.; Clarke, L.A.; Mendes, K.; Solé, A.; Pastor, J.; Margarida, M.; Amaral, M.D. Measurements of Functional Responses in Human Primary Lung Cells as a Basis for Personalized Therapy for Cystic Fibrosis. EBioMedicine 2015, 2, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Awatade, N.T.; Ramalho, S.; Silva, I.A.L.; Felício, V.; Botelho, H.M.; de Poel, E.; Vonk, A.; Beekman, J.M.; Farinha, C.M.; Amaral, M.D. R560S: A class II CFTR mutation that is not rescued by current modulators. J. Cyst. Fibros. 2019, 18, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Berkers, G.; Kruisselbrink, E.; Vonk, A.; De Jonge, H.R.; Janssens, H.M.; Bronsveld, I.; Van De Graaf, E.A.; Nieuwenhuis, E.E.S.; Houwen, R.H.J.; et al. Characterizing responses to CFTR-modulating drugs using rectal organoids derived from subjects with cystic fibrosis. Sci. Transl. Med. 2016, 8, 344ra84. [Google Scholar] [CrossRef]
- Veit, G.; Xu, H.; Dreano, E.; Avramescu, R.G.; Bagdany, M.; Beitel, L.K.; Roldan, A.; Hancock, M.A.; Lay, C.; Li, W.; et al. Structure-guided combination therapy to potently improve the function of mutant CFTRs. Nat. Med. 2018, 24, 1732–1742. [Google Scholar] [CrossRef]
- Silva, I.A.L.; Doušová, T.; Ramalho, S.; Centeio, R.; Clarke, L.A.; Railean, V.; Botelho, H.M.; Holubová, A.; Valášková, I.; Yeh, J.T.; et al. Organoids as a personalized medicine tool for ultra-rare mutations in cystic fibrosis The case of S955P and 1717-2A > G. Biochim. Biophys. Acta-Mol. Basis Dis. 2020, 1866, 165905. [Google Scholar] [CrossRef]
- Laselva, O.; Eckford, P.D.; Bartlett, C.; Ouyang, H.; Gunawardena, T.N.; Gonska, T.; Moraes, T.J.; Bear, C.E. Functional rescue of c.3846G>A (W1282X) in patient-derived nasal cultures achieved by inhibition of nonsense mediated decay and protein modulators with complementary mechanisms of action. J. Cyst. Fibros. 2020, 19, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Laselva, O.; Moraes, T.J.; He, G.; Bartlett, C.; Szàrics, I.; Ouyang, H.; Gunawardena, T.N.A.; Strug, L.; Bear, C.E.; Gonska, T. The cftr mutation c.3453g > c (d1152h) confers an anion selectivity defect in primary airway tissue that can be rescued by ivacaftor. J. Pers. Med. 2020, 10. [Google Scholar] [CrossRef]
- Wu, Y.S.; Jiang, J.; Ahmadi, S.; Lew, A.; Laselva, O.; Xia, S.; Bartlett, C.; Ip, W.; Wellhauser, L.; Ouyang, H.; et al. ORKAMBI-mediated rescue of mucociliary clearance in cystic fibrosis primary respiratory cultures is enhanced by arginine uptake, arginase inhibition, and promotion of nitric oxide signaling to the cystic fibrosis transmembrane conductance regulator channe. Mol. Pharmacol. 2019, 96, 515–525. [Google Scholar] [CrossRef]
- Amato, F.; Scudieri, P.; Musante, I.; Tomati, V.; Caci, E.; Comegna, M.; Maietta, S.; Manzoni, F.; Di Lullo, A.M.; De Wachter, E.; et al. Two CFTR mutations within codon 970 differently impact on the chloride channel functionality. Hum. Mutat. 2019, 40, 742–748. [Google Scholar] [CrossRef]
- de Winter-De Groot, K.M.; Janssens, H.M.; van Uum, R.T.; Dekkers, J.F.; Berkers, G.; Vonk, A.; Kruisselbrink, E.; Oppelaar, H.; Vries, R.; Clevers, H.; et al. Stratifying infants with cystic fibrosis for disease severity using intestinal organoid swelling as a biomarker of CFTR function. Eur. Respir. J. 2018, 52. [Google Scholar] [CrossRef]
- Scudieri, P.; Musante, I.; Venturini, A.; Guidone, D.; Genovese, M.; Cresta, F.; Caci, E.; Palleschi, A.; Poeta, M.; Santamaria, F.; et al. Ionocytes and CFTR Chloride Channel Expression in Normal and Cystic Fibrosis Nasal and Bronchial Epithelial Cells. Cells 2020, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pedemonte, N.; Bertozzi, F.; Caci, E.; Sorana, F.; Di Fruscia, P.; Tomati, V.; Ferrera, L.; Rodríguez-Gimeno, A.; Berti, F.; Pesce, E.; et al. Discovery of a picomolar potency pharmacological corrector of the mutant CFTR chloride channel. Sci. Adv. 2020, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Randell, S.; Fulcher, M.; O’Neal, W.; Olsen, J. Primary epithelial cell models for cystic fibrosis research. Methods Mol. Biol. 2011, 742, 285–310. [Google Scholar] [PubMed]
- Van Mourik, P.; Beekman, J.M.; Van Der Ent, C.K. Intestinal organoids to model cystic fibrosis. Eur. Respir. J. 2019, 54, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.A.L.; Duarte, A.; Marson, F.A.L.; Centeio, R.; Doušová, T.; Kunzelmann, K.; Amaral, M.D. Assessment of Distinct Electrophysiological Parameters in Rectal Biopsies for the Choice of the Best Diagnosis/Prognosis Biomarkers for Cystic Fibrosis. Front. Physiol. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Mall, M.; Hirtz, S.; Gonska, T.; Kunzelmann, K. Assessment of CFTR function in rectal biopsies for the diagnosis of cystic fibrosis. J. Cyst. Fibros. 2004, 3, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Hirtz, S.; Gonska, T.; Seydewitz, H.H.; Thomas, J.; Greiner, P.; Kuehr, J.; Brandis, M.; Eichler, I.; Rocha, H.; Lopes, A.I.; et al. CFTR Cl-Channel Function in Native Human Colon Correlates With the Genotype and Phenotype in Cystic Fibrosis. Gastroenterology 2004, 127, 1085–1095. [Google Scholar] [CrossRef]
- Sousa, M.; Servidoni, M.F.; Vinagre, A.M.; Ramalho, A.S.; Ribeiro, M.A.; Uliyakina, I.; Marson, F.A.; Bonadia, L.C.; Ribeiro, D.; Bertuzzo, C.S.; et al. Measurements of CFTR-Mediated Cl 2 Secretion in Human Rectal Biopsies Constitute a Robust Biomarker for Cystic Fibrosis Diagnosis and Prognosis. PLoS ONE 2012, 7, e47708. [Google Scholar] [CrossRef] [PubMed]
- Ratjen, F.; Döring, G. Cystic fibrosis. Lancet 2003, 361, 681–689. [Google Scholar] [CrossRef]
- Servidoni, M.F.; Sousa, M.; Vinagre, A.M.; Cardoso, S.R.; Ribeiro, M.A.; Meirelles, L.R.; de Carvalho, R.B.; Kunzelmann, K.; Ribeiro, A.F.; Ribeiro, J.D.; et al. Rectal forceps biopsy procedure in cystic fibrosis: Technical aspects and patients perspective for clinical trials feasibility. BMC Gastroenterol. 2013, 13, 8–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonk, A.M.; van Mourik, P.; Ramalho, A.S.; Silva, I.A.L.; Statia, M.; Kruisselbrink, E.; Suen, S.W.F.; Dekkers, J.F.; Vleggaar, F.P.; Houwen, R.H.J.; et al. Protocol for Application, Standardization and Validation of the Forskolin-Induced Swelling Assay in Cystic Fibrosis Human Colon Organoids. STAR Protoc. 2020, 1, 100019. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.M.; Mendes, F.; Dragomir, A.; Doull, I.J.M.; Carvalho-Oliveira, I.; Bebok, Z.; Clancy, J.P.; Eubanks, V.; Sorscher, E.J.; Roomans, G.M.; et al. Assessment of CFTR localisation in native airway epithelial cells obtained by nasal brushing. J. Cyst. Fibros. 2004, 3, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Hagemeijer, M.; Vonk, A.; Awatade, N.; Silva, I.; Tischer, C.; Hilsenstein, V.; Beekman, J.; Amaral, M.; Botelho, H. An open-source high-content analysis workflow for CFTR function measurements using the forskolin-induced swelling assay. Bioinformatics 2020, btaa1073. [Google Scholar] [CrossRef]
- Ramalho, A.S.; Förstová, E.; Vonk, A.M.; Ferrante, M.; Verfailli, C.; Dupont, L.; Boon, M.; Proesmans, M.; Beekma, J.M.; Sarouk, I.; et al. Correction of CFTR function in intestinal organoids to guide treatment of cystic fibrosis. Eur. Respir. J. 2021, 57, 1902426. [Google Scholar] [CrossRef]
- Laselva, O.; McCormack, J.; Bartlett, C.; Ip, W.; Gunawardena, T.N.A.; Ouyang, H.; Eckford, P.D.W.; Gonska, T.; Moraes, T.J.; Bear, C.E. Preclinical studies of a rare cf-causing mutation in the second nucleotide binding domain (C.3700a>g) show robust functional rescue in primary nasal cultures by novel cftr modulators. J. Pers. Med. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Amaral, M.D. How to determine the mechanism of action of CFTR modulator compounds: A gateway to theranostics. Eur. J. Med. Chem. 2021, 210, 112989. [Google Scholar] [CrossRef]
- Molinski, S.V.; Ahmadi, S.; Ip, W.; Ouyang, H.; Villella, A.; Miller, J.P.; Lee, P.; Kulleperuma, K.; Du, K.; Di Paola, M.; et al. Orkambi® and amplifier co-therapy improves function from a rare CFTR mutation in gene-edited cells and patient tissue. EMBO Mol. Med. 2017, 9, 1224–1243. [Google Scholar] [CrossRef]
- Kato, A.; Romero, M.F. Regulation of Electroneutral NaCl Absorption by the Small Intestine. Annu. Rev. Physiol. 2011. [Google Scholar] [CrossRef] [Green Version]
- Phuan, P.W.; Haggie, P.M.; Tan, J.A.; Rivera, A.A.; Finkbeiner, W.E.; Nielson, D.W.; Thomas, M.M.; Janahi, I.A.; Verkman, A.S. CFTR modulator therapy for cystic fibrosis caused by the rare c.3700A>G mutation. J. Cyst. Fibros. 2020, S1569-1993, 1–8. [Google Scholar]
- Roth, E.K.; Hirtz, S.; Duerr, J.; Wenning, D.; Eichler, I.; Seydewitz, H.H.; Amaral, M.D.; Mall, M.A. The K + channel opener 1-EBIO potentiates residual function of mutant CFTR in rectal biopsies from cystic fibrosis patients. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkers, G.; van Mourik, P.; Vonk, A.M.; Kruisselbrink, E.; Dekkers, J.F.; de Winter-de Groot, K.M.; Arets, H.G.M.; Marck-van der Wilt, R.E.P.; Dijkema, J.S.; Vanderschuren, M.M.; et al. Rectal Organoids Enable Personalized Treatment of Cystic Fibrosis. Cell Rep. 2019, 26, 1701–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laselva, O.; Bartlett, C.; Gunawardena, T.N.A.; Ouyang, H.; Eckford, P.D.W.; Moraes, T.J.; Bear, C.E.; Gonska, T. Rescue of multiple class II CFTR mutations by elexacaftor+ tezacaftor+ivacaftor mediated in part by the dual activities of Elexacaftor as both corrector and potentiator. Eur. Respir. J. 2020, 2002774. [Google Scholar] [CrossRef]
- Veit, G.; Roldan, A.; Hancock, M.A.; da Fonte, D.F.; Xu, H.; Hussein, M.; Frenkiel, S.; Matouk, E.; Velkov, T.; Lukacs, G.L. Allosteric folding correction of F508del and rare CFTR mutants by elexacaftor-tezacaftor-ivacaftor (Trikafta) combination. JCI Insight 2020, 5, 1–14. [Google Scholar] [CrossRef]
- Laselva, O.; Bartlett, C.; Popa, A.; Ouyang, H.; Gunawardena, T.N.A.; Gonska, T.; Moraes, T.J.; Bear, C.E. Emerging preclinical modulators developed for F508del-CFTR have the potential to be effective for ORKAMBI resistant processing mutants. J. Cyst. Fibros. 2021, 20, 106–119. [Google Scholar] [CrossRef]
- Brewington, J.J.; Filbrandt, E.T.; LaRosa, F.J.; Moncivaiz, J.D.; Ostmann, A.J.; Strecker, L.M.; Clancy, J.P. Brushed nasal epithelial cells are a surrogate for bronchial epithelial CFTR studies. JCI Insight 2018, 3, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual Code | Genotype |
|---|---|
| 1 | WT |
| 2 | F508del/F508del |
| 3 | P205S/Y1092X |
| 4 | D614G/F508del |
| 5 | N1303K/F508del |
| 6 | G85E/1717-1G>A |
| 7 | P205S/Y1092X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, I.A.L.; Railean, V.; Duarte, A.; Amaral, M.D. Personalized Medicine Based on Nasal Epithelial Cells: Comparative Studies with Rectal Biopsies and Intestinal Organoids. J. Pers. Med. 2021, 11, 421. https://doi.org/10.3390/jpm11050421
Silva IAL, Railean V, Duarte A, Amaral MD. Personalized Medicine Based on Nasal Epithelial Cells: Comparative Studies with Rectal Biopsies and Intestinal Organoids. Journal of Personalized Medicine. 2021; 11(5):421. https://doi.org/10.3390/jpm11050421
Chicago/Turabian StyleSilva, Iris A. L., Violeta Railean, Aires Duarte, and Margarida D. Amaral. 2021. "Personalized Medicine Based on Nasal Epithelial Cells: Comparative Studies with Rectal Biopsies and Intestinal Organoids" Journal of Personalized Medicine 11, no. 5: 421. https://doi.org/10.3390/jpm11050421
APA StyleSilva, I. A. L., Railean, V., Duarte, A., & Amaral, M. D. (2021). Personalized Medicine Based on Nasal Epithelial Cells: Comparative Studies with Rectal Biopsies and Intestinal Organoids. Journal of Personalized Medicine, 11(5), 421. https://doi.org/10.3390/jpm11050421