1. Introduction
After their maturation in the thymus, the naïve CD4+ T lymphocytes are activated through antigen recognition. This recognition consists of the binding of TCR and CD4 receptors with MHC class II, which carries the specific antigen on the surface of antigen presenting cells (APCs). However, this interaction is not enough to trigger an immune reaction since a co-stimulatory signal is necessary. There are different co-stimulatory molecules that allow this process, namely, CD28 receptors in T lymphocytes being the most important [
1]. These co-stimulatory signals can also regulate the cytoskeleton rearrangement and adhesion of T lymphocytes. Therefore, it is known that apart from being critical for the activation, proliferation, and survival of CD4+ T lymphocytes, CD28 is also involved in the migration process of these cells [
2].
The lack of this CD28 molecule during activation results in partial activation or even an anergic state of T lymphocytes. However, this is not the case with CD4+CD28
null T lymphocytes that, instead of being anergic, have enhanced effector functions increasing their inflammatory features, such as cytokine production (IFNγ an TNF), accumulation of molecules involved in cellular cytotoxicity (perforin and granzyme), expression of NK cell-related receptors (NKRs), loss of their sensitivity to apoptosis induction or their resistance to the suppressive actions of regulatory T (Treg) cells [
3].
It is known that these CD4+CD28
null T lymphocytes are expanded in some situations not only as an immunosenescence process [
4] but also under other clinical conditions involving chronic activation of the immune system, such as viral infections or autoimmune diseases [
5]. In rheumatoid arthritis (RA), it has been specifically shown that CD4+CD28
null T lymphocytes are significantly increased in the blood and synovial fluid of these patients, and it has been suggested that these cells have an active role in the maintenance of inflammation [
6,
7]. In some of these situations where CD4+CD28
null T lymphocytes are expanded, it has been demonstrated that interleukin 15 (IL-15) has a role in their proliferation and enhances the effector response of CD4+CD28
null T lymphocytes against their specific chronic antigens [
8,
9]. In fact, elevated serum and synovial fluid levels of IL-15 have been detected in RA patients, suggesting an important role of this cytokine in disease immunopathology and a correlation with clinical activity [
10,
11]. As a result, IL-15 has been recently proposed as a biomarker candidate for prediction of the development of RA, especially in the early phase of disease [
12].
Although the characteristics of CD4+CD28
null T lymphocytes have been widely studied in these pathologic situations, little is known about the migration patterns that they use to reach the target tissue perpetuating the damage. The migration of mature T lymphocytes consists of their recirculation through the blood to lymph nodes in order to facilitate the encounter with their specific antigen. This situation happens in physiologic conditions, but when a tissue is infected, T lymphocytes can migrate and go into the target tissue. Different adhesion molecules, such as integrins (LFA1, VLA-4), chemokine receptors (CX3CR1, CCR5) and hyaluronic acid receptors (CD44), as well as Rho-GTPase proteins that control the cytoskeleton dynamics of these cells (RhoA, Rac1/2, Cdc42), are essential for this process [
13,
14,
15]. Besides these proteins, there are a high number of genes that are regulating complex pathways related to the migration process, such as focal adhesion, leucocyte extravasation and actin cytoskeleton regulation. With respect to CD4+CD28
null T lymphocytes, as we have advanced above, they have been identified not just in the circulation of patients with inflammatory diseases but also in target tissues, such as the joints of RA patients [
6]. Some studies suggest that expression of chemokine receptors and adhesion molecules by CD4+CD28
null T lymphocytes could have an important role in guiding their migration and ability to infiltrate tissues, which allow these cells to cause local inflammation and tissue damage [
3].
The goal of this work is to study the adhesion and migration abilities of CD4+CD28null T lymphocytes in RA patients, as well as the effect that IL-15 could have in these processes, in order to find possible candidates for therapeutic targets.
2. Materials and Methods
2.1. Donors
Peripheral blood samples were obtained from 65 donors with a diagnosis of RA according to the American College of Rheumatology (ACR) and the European League Against Rheumatism (EULAR) classification criteria in the Hospital Universitario Central de Asturias (Oviedo, Spain). All these RA patients selected for the study were in the age range of 40–65 and had positive anti-cyclic citrullinated peptide (anti-CCP) and/or rheumatoid factor (RF) tests. The threshold for considering these values positive were ≥3 U/mL for anti-CCP and >14 Ku/L for RF. Experiments were performed with blood samples from those individuals whose percentage of CD4+CD28null T lymphocytes was 5% or higher of total CD4+ T lymphocytes. The different experiments focused on analysing the factors associated with the migratory ability of CD4+CD28null T lymphocytes were performed with 41 of these donors that fulfilled this inclusion criterium. The study was approved by the Hospital Central de Asturias (Oviedo, Spain) Ethics Committee.
The ethics committee approved the exemption of informed consent in this study since only excesses of peripheral blood (intended to be destroyed) extracted for clinical purposes were used and never for the purpose of being used in research. The samples were anonymized from the first moment after clinical studies necessary for the clinical follow-up of RA disease were performed, and there was no record in which the origin of the samples could be determined.
2.2. Quantification of CD4+CD28null T Lymphocytes
The percentage of CD4+CD28null T lymphocytes was determined by staining with anti-CD3 (FITC), anti-CD28 (PE) (eBioscience, San Diego, CA, USA), anti-CD8 (PerCP), and anti-CD4 (APC) (Immunostep, Salamanca, Spain) monoclonal antibodies. A total of 100 µL of whole blood from the donors were stained with the combination of labelled monoclonal antibodies for 30 min at room temperature. Samples were red blood lysed for 10 min at room temperature with FACS Lysing Solution (BD Biosciences, San José, CA, USA), washed in PBS, and analysed with BD Accuri C6 Software in the Accuri C6 (BD Biosciences, San José, CA, USA).
2.3. Cell Isolation and Culture Conditions
Peripheral blood mononuclear cells (PBMCs) were isolated from peripheral blood that had been anticoagulated with EDTA by centrifugation on Ficoll–Hypaque gradients (Lymphoprep, Nycomed, Oslo, Norway).
CD4+ T lymphocytes were enriched by the incubation of the peripheral blood anticoagulated with EDTA with the RosetteSep™ Human CD4+ T Cell Enrichment Cocktail (STEMCELL, Vancouver, Britids Columbia, Canada) for 20 min at room temperature. After this incubation, CD4+ T lymphocytes were obtained by centrifugation on Ficoll-Hypaque gradients (Lymphoprep; Nycomed).
The CD4+ purified cells were separated into CD4CD28+ and CD4+CD28null using the CD28 MicroBead Kit and the separation columns MACS MS (MiltenyiBiotec, Bergisch Gladbach, Germany).
According to manufacturer instructions, starting with fresh human whole blood, the CD4+ T cell content of the enriched fraction is typically 94 ± 5% (mean ± SD). Specifically, in our experiments, the purity of both CD4, CD28+, and CD28null isolated cells was evaluated by flow cytometry, and it was never lower than 95%.
Cultures were performed in RPMI 1640 medium containing 2 × 10−3 M l-glutamine and Hepes (BioWhitaker, Verviers, Belgium) and supplemented with 10% FCS (ICN Flow; Costa Mesa, CA, USA) and antibiotics. Cells at a concentration of 2 × 106 cells/mL were incubated at 37 °C and 5% carbon dioxide.
2.4. Expression of Chemokine Receptors and Adhesion Molecules
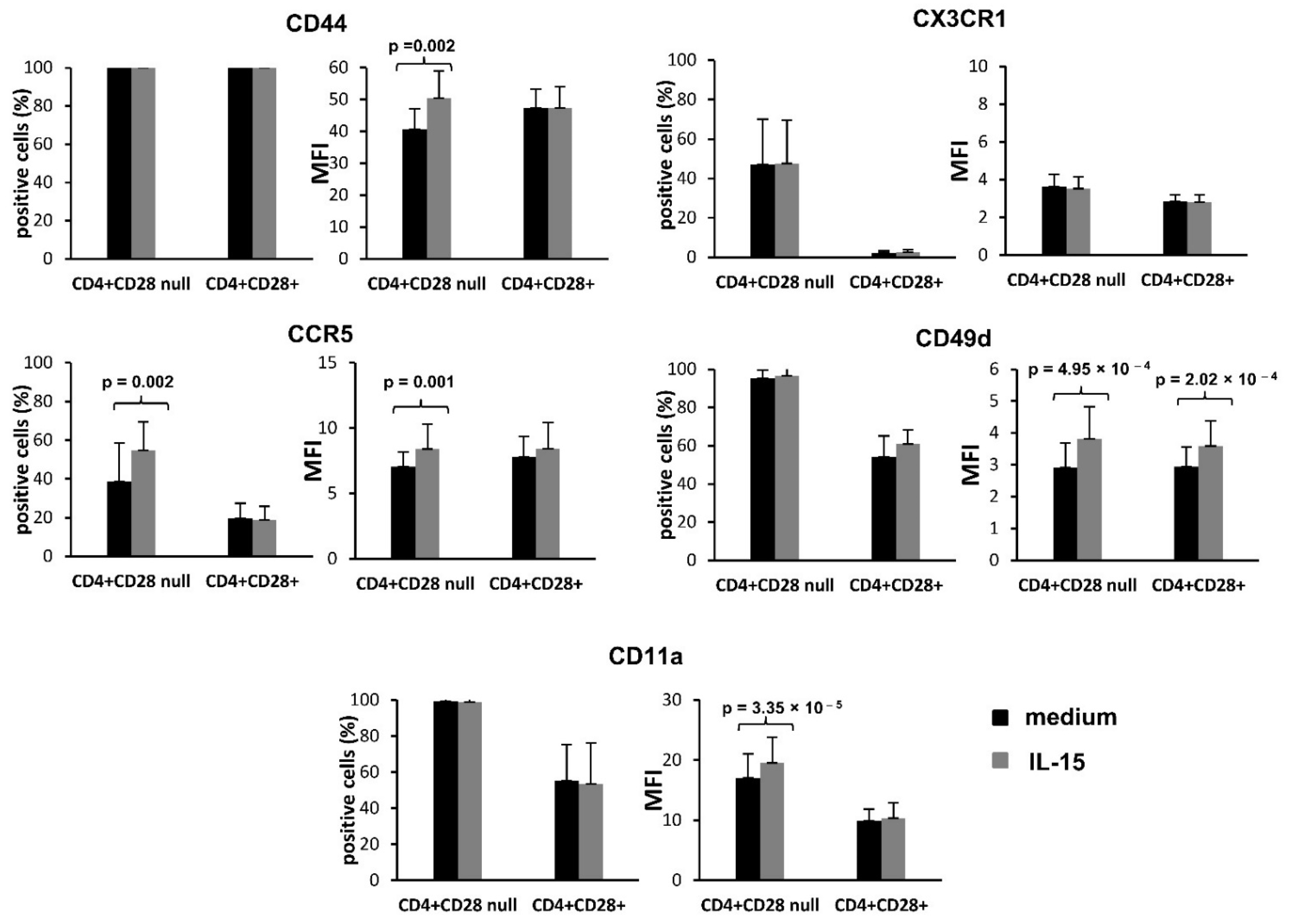
Basal expression of the different molecules was determined in peripheral blood obtained from seven RA patients, as described above. The effect of IL-15 on the expression of these molecules was studied in PBMCs obtained from 10 RA patients and cultured in medium alone or in the presence of IL-15 (50 ng/mL) (Peprotech INC, Rockyhill, NJ, USA) for 18 h. Overnight treatment with IL-15 has been seen to be the appropriate frame time for protein expression detection in previous studies [
8,
16]. The surface stain in both cases was made with anti-CD4 (APC) or anti-CD4 (PECy7), anti CD45RA (APCFire), anti-CD8 (PB), anti-CD28 (BV), anti-CD44 (FITC), anti-CX3CR1 (PE), anti-CCR5 (PECy7), and anti-CD11a (FITC) (Biolegend, San Diego, CA, USA), as well as anti-CD3 (PerCP) and anti-CD49d (APC) (BD Bioscience). Cells from whole blood were stained as described above. PBMCs were stained for 30 min at 4 °C. Then, cells were washed and resuspended in PBS until they were acquired in a Navios flow cytometer and analysed with Kaluza software (Beckman Coulter Life Science, Brea, CA, USA). The cytometer compensation was carried out using the VersaComp Antibody Capture Bead Kit (Beckman Coulter). The marker settings for determining the negative/positive cell populations were established using the FMO strategy.
2.5. Cell Migration Assays
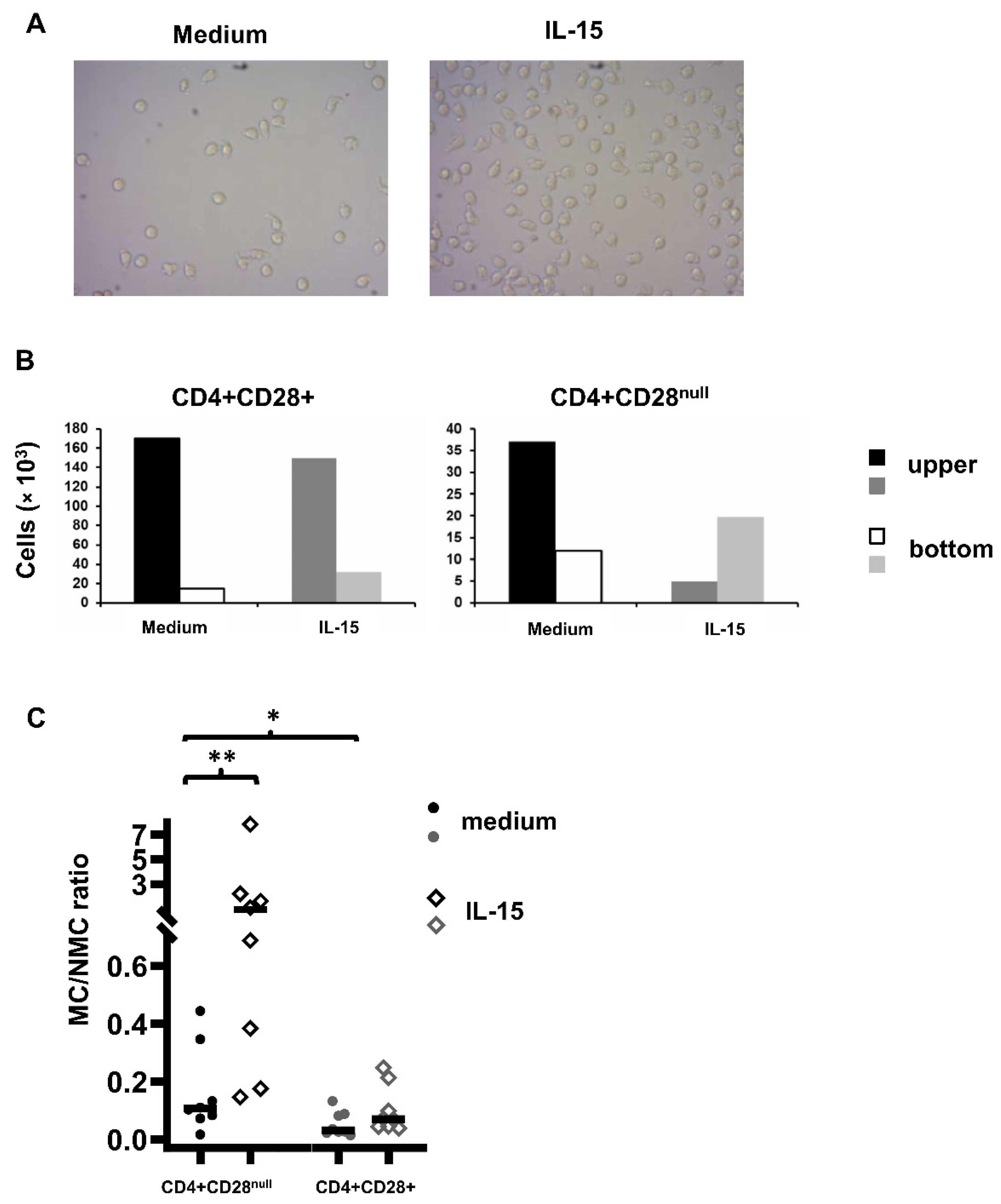
Cell migration assays were performed with CD4+ T lymphocytes isolated from eight patients using a transwell system with a pore size of 3 µm (Costar, Kennebunk, ME, USA). Briefly, between 2 × 105 and 5 × 105 CD4+ T lymphocytes in 100 µL of medium were placed in the upper chamber of a transwell system suspended over a larger well containing 600 µL of culture medium or in the presence of IL-15 (50 ng/mL). Cells were allowed to migrate through the pores to the other side of the membrane. After 18 h, migratory cells (MCs) and non-migratory cells (NMCs) were collected. Then, they were surface stained with anti-CD45RA (FITC) (Biolegend), anti-CD28 (PE), and anti-CD4 (PerCP) (BD Bioscience) monoclonal antibodies. Flow cytometric analysis was made using the Accuri C6 (BD Biosciences). For the cell count, 20 µL of the sample was acquired at medium speed. We have tested different concentration of IL-15 (from 0.5 to 100 ng/mL) to perform these transwell assays and we have observed a significant increment in the cell migration capacity with all the IL-15 concentrations tested comparing to unstimulated conditions. The maximum peak of migration was already reached for both T CD4+ lymphocyte subsets studied with 50ng/mL (data not shown). Based on this, we decided to use this IL-15 concentration for the rest of experiments.
2.6. Migration RT-PCR Array
Isolated CD4+CD28+ and CD4+CD28
null T lymphocytes from eight patients were pooled and cultured in medium alone or in the presence of IL-15 stimulation (50 ng/mL) for 4 h. We have chosen 4 h as the time to perform the gene expression experiments based on the previous knowledge about PBMCs gene expression after its stimulation in cell culture and on data from our own experience [
16]. mRNA from these cells was extracted using the RNeasy Mini Kit (Qiagen, Düsseldorf, Germany) according to the manufacturer’s instructions, including the optional on-column DNase digestion step with RNAse-Free DNase set (Qiagen). Reverse transcription of mRNA isolated from each sample was carried out in a 20 μL final volume with the cDNA Synthesis Using the RT2 First Strand Kit (Qiagen), as indicated in the instructions provided by the manufacturer. cDNA was stored at −80 °C until required for RT-PCR array. An RT-PCR array of 96 wells was performed for each cDNA sample following manufacturer’s instructions (PAHS-128Z RT2 ProfilerTM PCR Array Human Cell Motility, Qiagen) and analysed using a 7500 Real Time PCR System (Applied Biosystem, Foster City, CA, USA).
RT-PCR array analysis was conducted at Qiagen’s GeneGlobe Data Analysis Center using a software tool available on the website.
All the samples passed the three control tests performed before the analysis: PCR array reproducibility, reverse transcription efficiency, and genomic DNA contamination. Two constitutively expressed genes, glyceraldehide-3-phosphate dehydrogenase and beta-2-microglobulin, were used for data normalization.
We compared two different conditions: CD4+CD28null (test sample) vs. CD4+CD28+ (control sample) and CD4+CD28null IL-15 stimulation (test sample) vs. CD4+CD28null basal conditions (control sample). Fold change 2^(ΔΔCT) that is the normalized gene expression (2^(−ΔCT)) in the test sample dividing the normalized gene expression (2^(−ΔCT)) in the control sample was calculated for each of the compared conditions.
In order to represent fold change results in a biologically meaningful way, fold regulation was calculated. When fold change values are greater than one, it indicates an upregulation and fold regulation is equal to the fold change in these cases. However, when fold change values are less than one, it indicates a downregulation, and fold regulation is calculated as the negative inverse of the fold change. Genes were considered significantly upregulated when the fold regulation was greater than 2 and downregulated when the fold regulation was lower than −2.
Log
2 of the fold change was calculated for the heat maps representation and pathway analysis in order to normalize and get a better visualization of the data. The pathway-based data integration and visualization was performed using the pathview R package (1.30.1 version) from the bioconductor bioinformatics repository [
17].
2.7. Activity Quantification of Rho GTPases Involved in Cellular Migration
Isolated CD4+CD28+ and CD4+CD28null T lymphocytes from eight patients were pooled and cultured in serum-free conditions for 1 h. After this, lymphocytes were cultured in completed medium alone or in the presence of IL-15 stimulation (50 ng/mL) for 5 min. It was well documented that the GTPases activation occurs in a very short period of time (a few minutes), and as it is said in the manufacturer instructions of G-LISA activation assays (Cytoskeleton, Inc., Denver, CO, USA), Rho proteins are generally activated very rapidly and transiently (30 s to 30 min). Following this, we decided to incubate under IL-15 stimulation for 5 min. After the incubation, cold PBS was added to the cells in order to stop the stimulation. Protein extracts were obtained from each condition and both basal levels of RhoA and activated forms of RhoA, Rac1, and Cdc42 were measured following the manufacturer instructions using G-LISA activation assays (Cytoskeleton, Inc., Denver, CO, USA). Calpeptine was used as the positive control for RhoA activation.
2.8. Statistical Analysis
Comparisons between groups were performed with the nonparametric Wilcoxon signed-rank test when data was not normally distributed or with Student’s t test for paired data when data was normally distributed. To compare the migratory abilities between different conditions, data were logarithmically transformed and a post hoc multiple comparisons with Bonferroni adjustment was performed. Analyses were performed using the PASW Statistics 17.0 statistical software package (IBM SPSS, Armonk, NY, USA), and p-values of 0.05 or less were considered significant.
4. Discussion
CD4+CD28
null T lymphocytes represent a cell subset with special peculiarities that make them different from the rest of the T helper cells. For instance, they have increased effector functions triggering a higher inflammatory capacity [
3]. The amount of this cell subset has been expanded in ageing and pathogenic situations, such as chronic viral infections and autoimmune diseases, being widely described in RA patients [
4]. In RA disease, it has been shown that CD4+CD28
null T lymphocytes and IL-15 levels are significantly increased in the blood and synovial fluid of these patients, suggesting an active role of both of them in the maintenance of inflammation in the target tissue, which could be correlated with the immunopathology and clinical activity of the disease [
9,
10,
11,
12]. Based on all this, we decided to study the migratory capacity of these CD4+CD28
null T lymphocytes and the effect that IL-15 could have in this process.
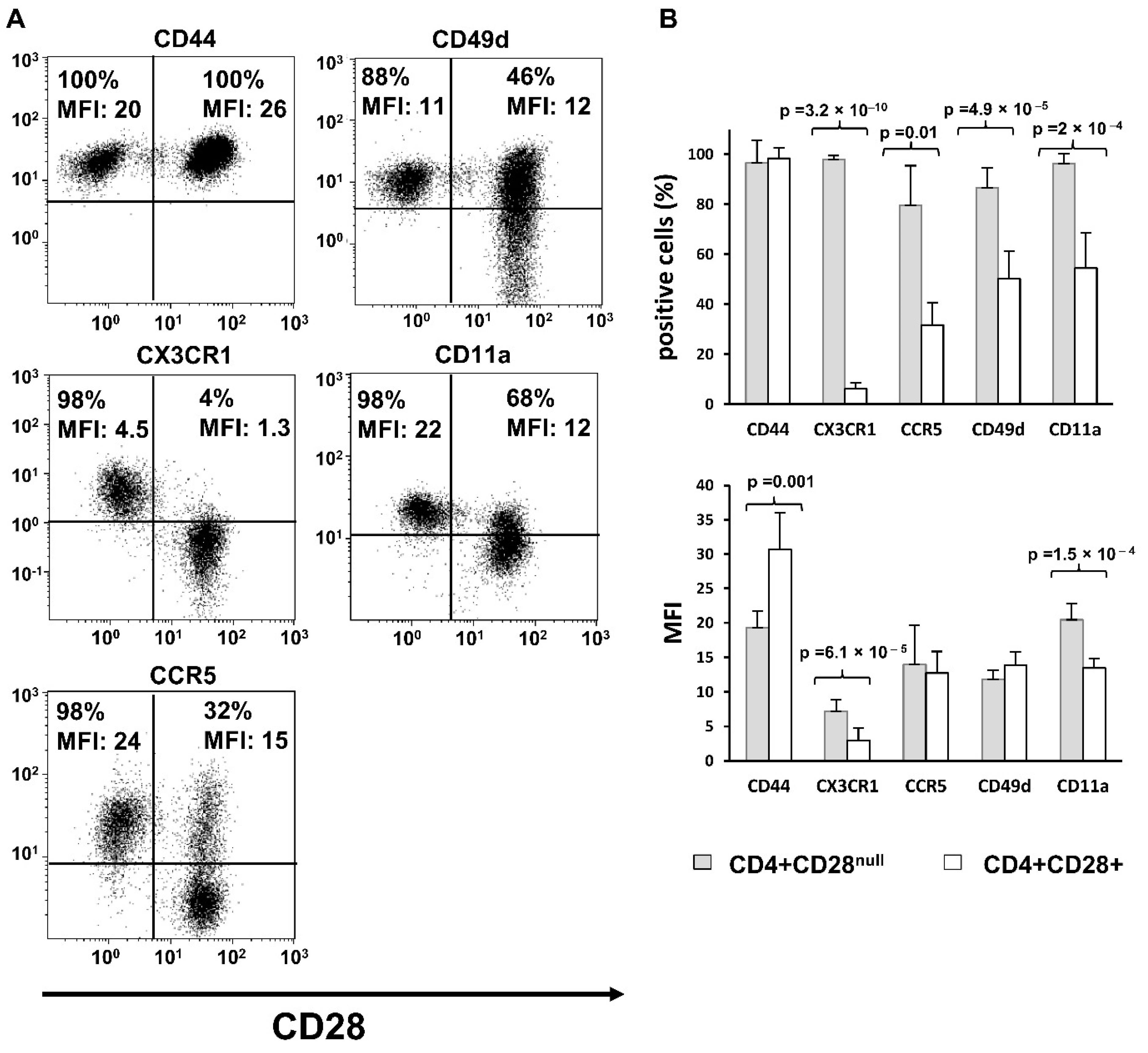
We have observed that the levels of expression of the adhesion molecules CD49d and CD11a and of the chemokine receptors CX3CR1 and CCR5, were higher in CD4+CD28
null T lymphocytes than in CD4+CD28+ T lymphocytes. Furthermore, a greater enhancing effect of IL-15 on expression of these molecules was also predominantly found in this T cell subtype. These results agree with the previously published studies demonstrating a greater expression of CD11a, CD49d and CX3CR1 in CD4+CD28
null T lymphocytes than in CD4+CD28+ T lymphocytes [
18]. Specifically, CX3CR1, the receptor of fractalkine/CX3CL1, was one of the molecules much more expressed in CD4+CD28
null T lymphocytes than in CD4+CD28+ T lymphocytes. It has been demonstrated that fractalkine/CX3CL1 plays an important role in the development of RA, suggesting a more aggressive role of this molecule in RA pathogenesis, as well as an association with atherosclerotic damage in these patients. CD4+CD28
null T lymphocytes may participate in both processes, being attracted by fractalkine to the inflammatory sites [
19,
20]. In contrast to the study of Broux and collaborators made on MS patients [
18], we did not find an increase in CX3CR1 expression in response to IL-15 treatment, which is probably due to the different culture times used. However, CCR5, LFA-1 (CD11a/CD18), and VLA-4 (CD49d/CD29) were positively regulated in both studies. Most of the effects on the migratory ability of lymphocytes caused by IL-15 have been demonstrated in NK cells, in which this cytokine acts as a chemotactic factor that stimulates adhesion of NK cells to endothelial cells. Moreover, it is known that IL-15 also enhances the transendothelial migration of T lymphocytes by the activation of LFA-1 binding capacity to its ICAM-1 ligand [
21]. Therefore, IL-15 is an important factor for NK cell recruitment to the target tissues, and this is possibly reproduced by CD4+CD28
null T lymphocytes [
22]. It should be noted that both cell types share other functional properties and surface expression of several markers. These results also agree with those observed in the gene expression array experiments where the gene
ITGB2 coding CD18 protein, which can form part of the LFA integrin, was found overexpressed in CD4+ CD28
null T lymphocytes when compared with CD4+CD28+ T lymphocytes.
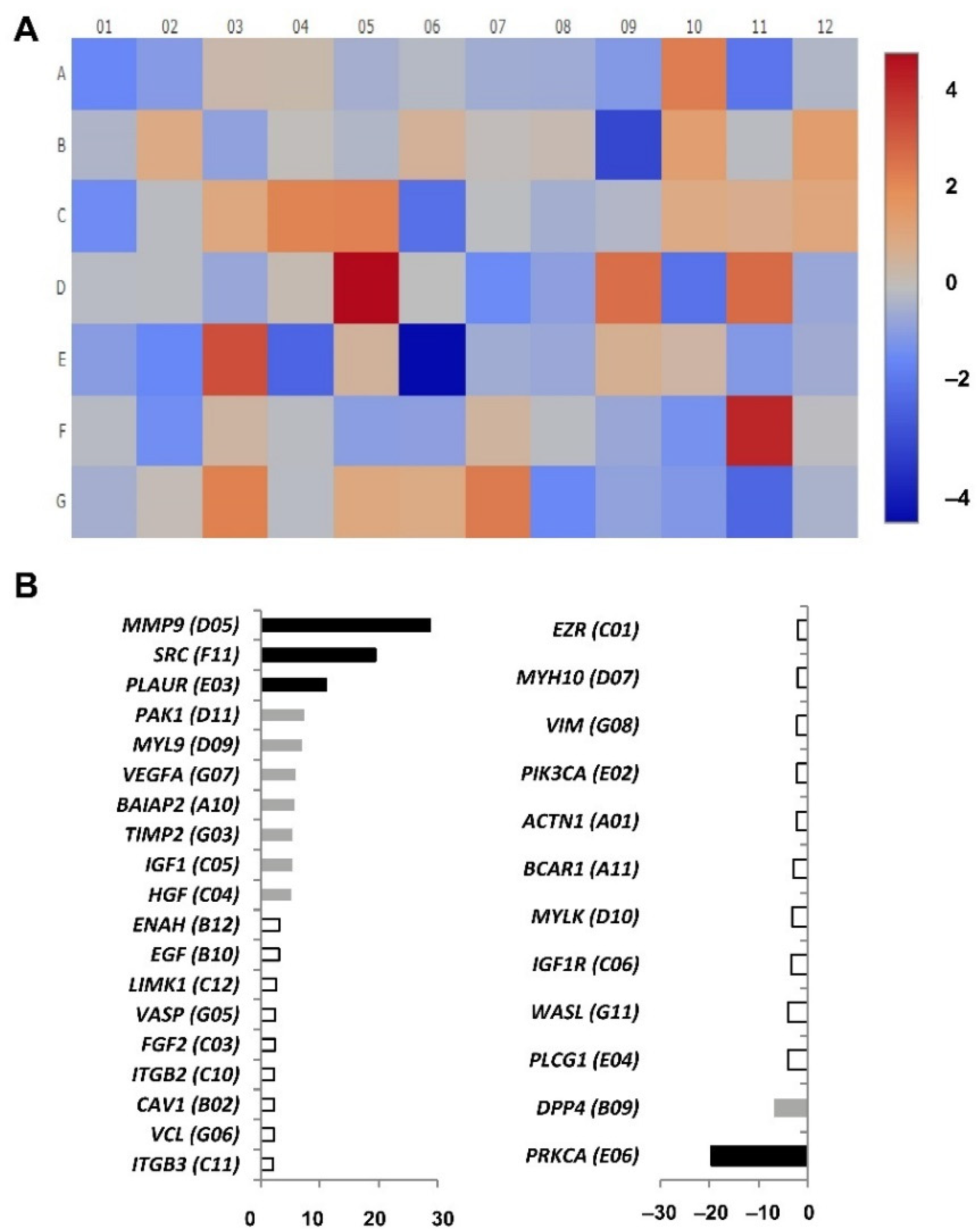
Apart from it, other genes importantly related to migration have an upregulated expression in CD4+CD28null T lymphocytes when compared with CD4+CD28+ T lymphocytes.
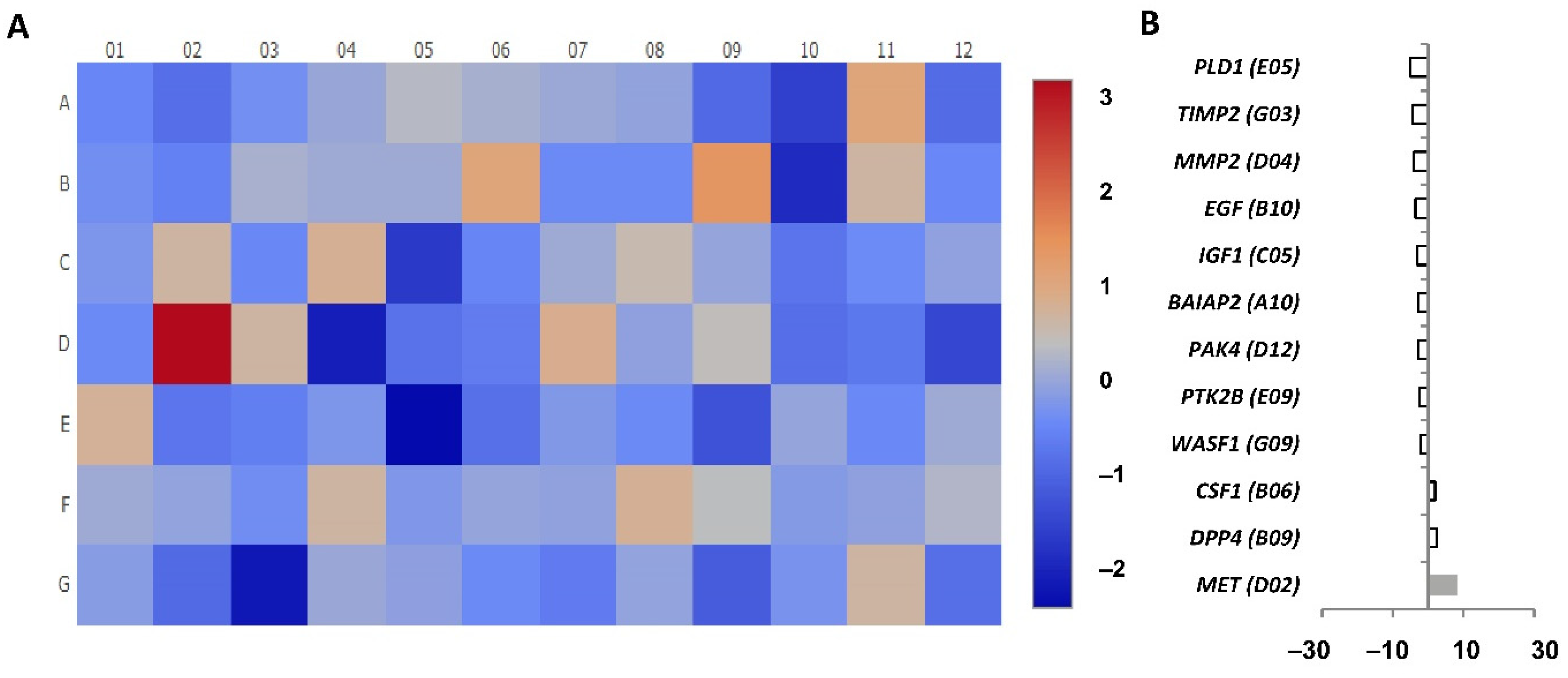
MMP9 was the gene with the highest expression difference detected between CD4+CD28
null T lymphocytes and CD4+CD28+ T lymphocytes, being its expression almost 30-fold higher in CD4+CD28
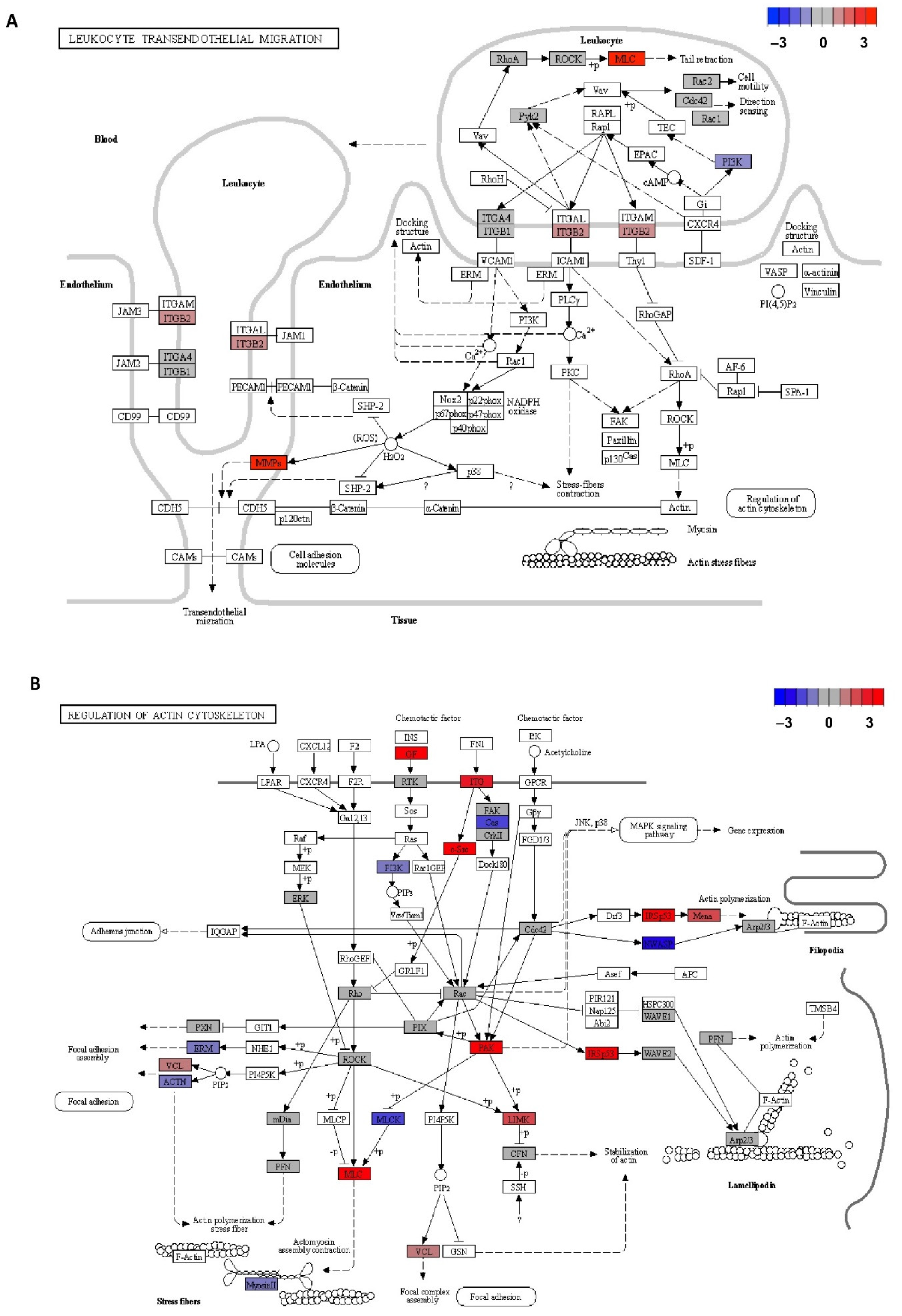
null cells. Lymphocyte binding to the endothelial cell during the transmigration process is mediated by receptors, such as vascular cell adhesion molecule 1 (VCAM-1). As is shown in
Figure 5A, this interaction induces NADPH oxidase activation and the production of reactive oxygen species (ROS) in a Rac-mediated manner, with subsequent activation of MMPs and loss of VE-cadherin-mediated adhesion [
23]. It is described that MMP9 and MMP2 are the metalloproteinases that most contribute to protease activity associated with infiltrating leucocytes [
24]. Specifically, MMP9 or gelatinase B is known to be produced by polymorphonuclear leucocytes, and it was the metalloproteinase activity first detected in rheumatoid synovial fluid [
25]. Therefore, it is not a surprise that the gene coding this protein was overexpressed in CD4+CD28
null T lymphocytes which as was said above, is an expanded subset in autoimmune diseases, such as RA.
The
SRC proto-oncogene was detected in CD4+CD28
null T lymphocytes with an almost 20-fold increase as compared to that in CD4+CD28+ T lymphocytes. This proto-oncogene may play a role in the regulation of embryonic development and cell growth, and it also has been seen to be involved in the promotion of some tumours and their malignant progression [
26]. From an immunological point of view,
SRC has been seen to be involved in TLR2 phosphorylation, which triggers Ca
2+ fluxes, and it has been postulated that through this signalling event the cell junctions could be modified to facilitate the migration of polymorphonuclear leucocytes across the epithelial barrier [
27]. It is noteworthy that TLRs, although not typical of T lymphocytes, are expressed in certain T lymphocyte subpopulations, such as CD4+CD28
null, where TLR4 and TLR2 have been found [
28,
29].
PLAUR, which is a urokinase-type plasminogen activator receptor, was another gene whose expression was upregulated more than 10-fold in CD4CD28
null T lymphocytes. Different studies indicate that Plaur protein plays a key role in promoting neutrophil adhesion and their transendothelial migration for recruitment into inflamed tissues [
30,
31].
Otherwise, Rho GTPases, RhoA, Rac, and Cdc42, are core proteins of the actin cytoskeleton regulation necessary for cell migration, as is seen in
Figure 5B. In the classic model of cell migration, it is well-known that the GTPases Rac and Cdc42 are active at the leading edge of the cell, promoting bulge formation, whereas RhoA GTPase is only active in the cell body and at the rear in order to provide the actomyosin-mediated force needed for forward movement. Although, in our experiments, none of these GTPase genes were found as differentially expressed, some of the downstream targets of these proteins were detected as upregulated in CD4+CD28
null T lymphocytes as compared with CD4+CD28+ T lymphocytes. Moreover, in the GLISAS experiments, we detected enhanced activity of the three Rho GTPases after IL-15 stimulation in CD4+CD28
null T lymphocytes, while this treatment triggered the increased activity of Cdc42 in CD4+CD28+ T lymphocytes. Specifically, Rac proteins are necessary for promoting lamellipodia formation, which are cytoskeletal actin projections on the leading edge of the cell necessary for cell migration [
32]. Some of the genes regulated by Rac were detected as upregulated in CD4+CD28
null T lymphocytes as compared with CD4+CD28+ T lymphocytes. This is the case with
IRSp53, also known as
BAIAP2, whose coded protein is an essential intermediate between Rac and WAVE. WAVE complex proteins act by stimulating rapid actin polymerization through the Arp2/3 complex (actin-related proteins 2 and 3) [
33].
PAK and LIMK are also proteins regulated by Rac whose coding genes were found upregulated in CD4+CD28
null T lymphocytes. These two proteins exert their control over cofilin (CNF) whose function is to perform the actin turnover of the microfilaments of the cytoskeleton. The Rac/PAK signalling pathway also regulates myosin light chain (MLC/MYL9) phosphorylation, which increases actomyosin contractility at the cell periphery, although MLC activation is mainly triggered by the Rho/ROCK signalling pathway. Again, the
MLC gene was found upregulated in CD4+CD28
null cells, in this case more than 5-fold. Thus, Rac/PAK and Rho/ROCK signalling triggers cofilin inactivation on the one hand and myosin II activation through the MLC pathway on the other hand, causing overall actin cytoskeleton reorganization that leads to stress fibre assembly and lamellipodium formation in the cell periphery. These alterations in cell morphology are performed in order to favour cell migration [
34]. Besides this, in this study, significant differences were also found in the gene expression of some of the downstream targets of Cdc42 GTPase. The results suggest that, in CD4+CD28
null cells, Cdc42 exerts its function preferably by activating Arp2/3 through the IRSp53-Mena pathway better than by the alternative NWASP pathway, since the
IRSp53 and
MENA genes were found to be upregulated, while
NWASP was downregulated in this T lymphocyte subset. The IRSP53-MENA complex triggers actin filament assembly into filopodia, which are cytoplasmatic projections that extend beyond the leading edge of the lamellipodia in migrating cells [
35].
Growth factors have been seen as frequent outside activators of this actin cytoskeleton regulatory signalling pathway necessary for cell migration [
36] (
Figure 5B). In the present study, it is seen that several growth factors genes are found upregulated in CD4+CD28
null T lymphocytes, as in the case of
FGF2,
GFI1,
HGF,
EGF, and
VEGFA. These findings could suggest that migration of CD4+CD28
null cells could be induced by these different growth factors that might act in an autocrine manner. Specifically, HGF/Met induces the proliferation and migration of endothelial and tumour cells through RhoA, Rac1, and Cdc42 activation [
37].
HGF expression has been found increased in CD4+CD28
null T lymphocytes, whereas
MET was the gene found with the most increased expression in these cells in response to IL-15 stimulation. Recently, it has been found that a fraction of murine cytotoxic lymphocytes expresses c-Met and displays augmented cytolytic activity [
38]. It is important to take into account that IL-15 enhances the cytotoxic properties of CD4+CD28
null T lymphocytes, and c-Met may be implicated in this regulation.
Taking all these results together, it seems clear that CD4+CD28null lymphocytes are overexpressing essential genes for remodelling the actin cytoskeleton, specifically some of the downstream targets of RhoA, Rac, and Cdc42 GTPases. This fact can partially explain the higher migratory capacity of these cells, and as it is shown by these gene expression experiments, it is set from a transcriptional level.
Other interesting data extracted from this study are the downregulated genes observed in CD4+CD28
null lymphocytes with respect to CD4+CD28+ lymphocytes. Among these genes, the most downregulated gene is protein kinase C alpha (
PRKCA/PKCA) with an expression difference of almost 20-fold. PKC family members phosphorylate a wide variety of protein targets. Among other functions, PKCA protein is involved in the regulation of the immune response of CD4+ T cells, and specifically, it has been implicated in T cell receptor/CD28-induced interleukin 2 gene expression [
39]. This fact could explain why this gene is downregulated in cells without CD28 expression. PKCA has also been involved in migration signalling through a VCAM-1-dependent pathway [
40,
41]. Either way, the migration capacity of CD4+CD28
null cells seems to be PKCA-independent.
DDP4, also known as CD26, is another downregulated gene in CD4+CD28
null T lymphocytes, although IL-15 induced an increase in
DDP4 expression in both subsets of CD4+ cells. The protein coded by this gene is a T lymphocyte costimulator, with dipeptidyl peptidase enzymatic activity implicated in the development, maturation and migration of CD4+ T lymphocytes [
42]. It is interesting that different previous reports have shown that CD26+ T lymphocytes exhibit strong migratory ability through endothelial cells, and these cells have been detected at high levels in the synovium and synovial fluids of RA patients, suggesting a role in inducing the inflammation and tissue destruction of these cells [
43,
44]. Delving into this, it has already been reported in progressive multiple sclerosis patients that this CD26 T lymphocyte costimulator is mainly expressed in CD28+ T lymphocytes, and its expression is much lower in CD4+CD28
null T lymphocytes [
45]. Therefore, comparing our data with this, it seems that the CD26 costimulator is minimally expressed in CD4+CD28
null cells, but it is overexpressed in their counterpart, CD4+CD28+ T lymphocytes, in RA patients.
As it was described in
Section 3, our RT-PCR array experiments also reflected that there are almost no differences in the gene expression levels of the migration genes studied between basal conditions and IL-15 stimulation on CD4+CD28
null T lymphocytes. Something similar was observed with IL-15 stimulation of CD4+CD28+ T lymphocytes (
Figure S1). This finding seems to suggest that the role that IL-15 has in lymphocyte motility, specifically in CD4+CD28
null T lymphocytes, which were observed with other experiments, could not be regulated at the transcription level. It is quite possible that the migration effects observed with IL-15 are then regulated through post-transcriptional and post-translational modifications.
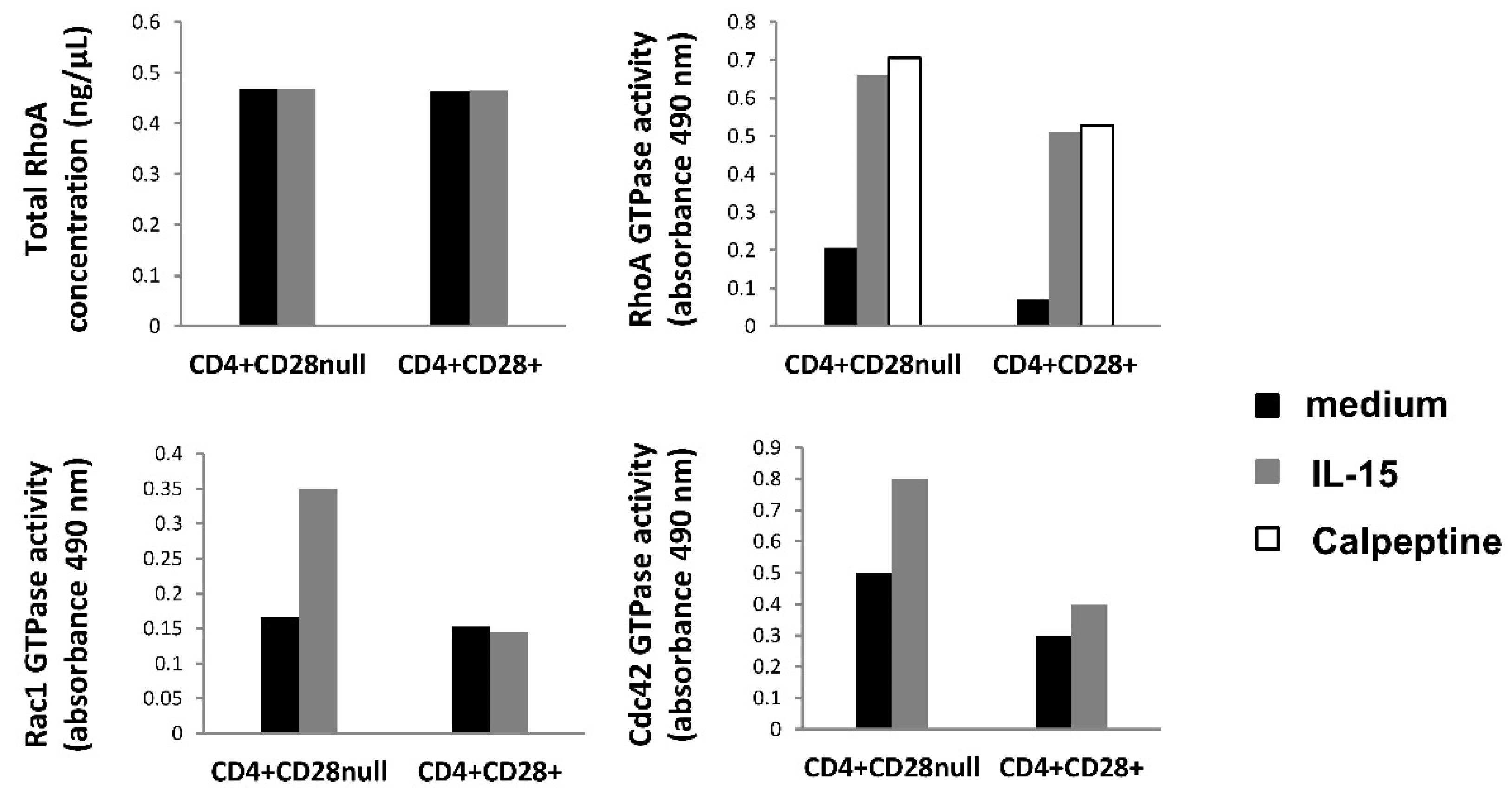
An important point to highlight from all these gene transcription results is the fact that none of the Rho GTPase genes showed any expression differences between the two different CD4+ T lymphocyte compartments. This fact fits with the RhoA basal levels measured in this work, which also reflected no differences between any of the conditions compared. Furthermore, our GLISAS experiments reflect that the activities of the proteins (RhoA, Rac1, and Cdc42) were higher in CD4+CD28
null T lymphocytes than in CD4+CD28+ T lymphocytes, and this basal activity increased after IL-15 stimulation, especially in CD4+CD28
null T lymphocytes. Hence, putting all these findings together, it may indicate that these proteins must be activated in these conditions by the post-translational mechanisms, which are already known for these RhoA GTPases [
46].
Until now, only one work that studied the effect of IL-15 on the migration capacity of the CD4+CD28
null T lymphocytes has been published, and in that case, it was made in multiple sclerosis patients [
18]. In this report, Broux and collaborators observed that IL-15 has more effect in CD4+CD28
null T lymphocytes than in CD4+CD28+ T lymphocytes, with regards to increasing their migration capacity. This migration capacity allows these cells to infiltrate the target tissue where they might trigger inflammatory damage. Our results agree quite well with this study, suggesting that this increase in the migration capacity probably depends on the T lymphocyte population itself more than on the particular disease. This fact makes it possible to extrapolate these results to any situation where the CD4+CD28
null T cell subset is expanded, as is the case with multiple sclerosis or RA.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






