
Differential Expression of Estrogen-Responsive Genes in Women with Psoriasis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Clinical Samples
2.3. Preparation of Skin Samples for LC–MS/MS Experiments
2.4. LC-MS/MS Analysis
2.5. Analysis of LC–MS/MS Data
2.6. Quantitative PCR
2.7. ELISA
2.8. Statistical Analysis
3. Results
3.1. The Blood of Female Psoriasis Patients Contains Less Estradiol and Progesterone, and More Testosterone Compared to the Healthy Volunteers
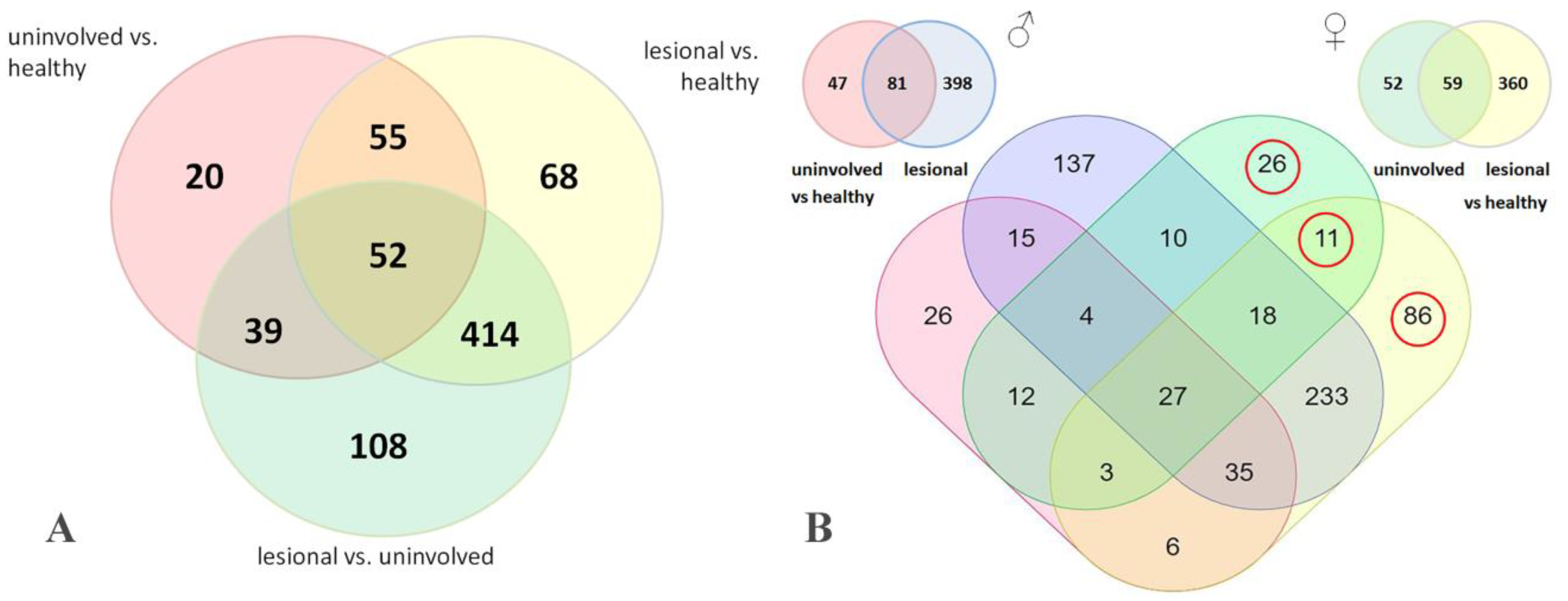
3.2. LC–MS/MS Study Identifies Six Estrogen-Responsive Proteins That Are Differentially Expressed in Male and Female Psoriatic Skin
3.3. The Identified Estrogen-Responsive Genes Were Differentially Expressed in Menopausal and Non-Menopausal Patients, and Their Expression Was Influenced by the Disease
3.4. Assessment of Protein Expression by ELISA Confirms a Differential Expression of KRT19 and HSPD1 in the Lesional Skin of Female Psoriasis Patients
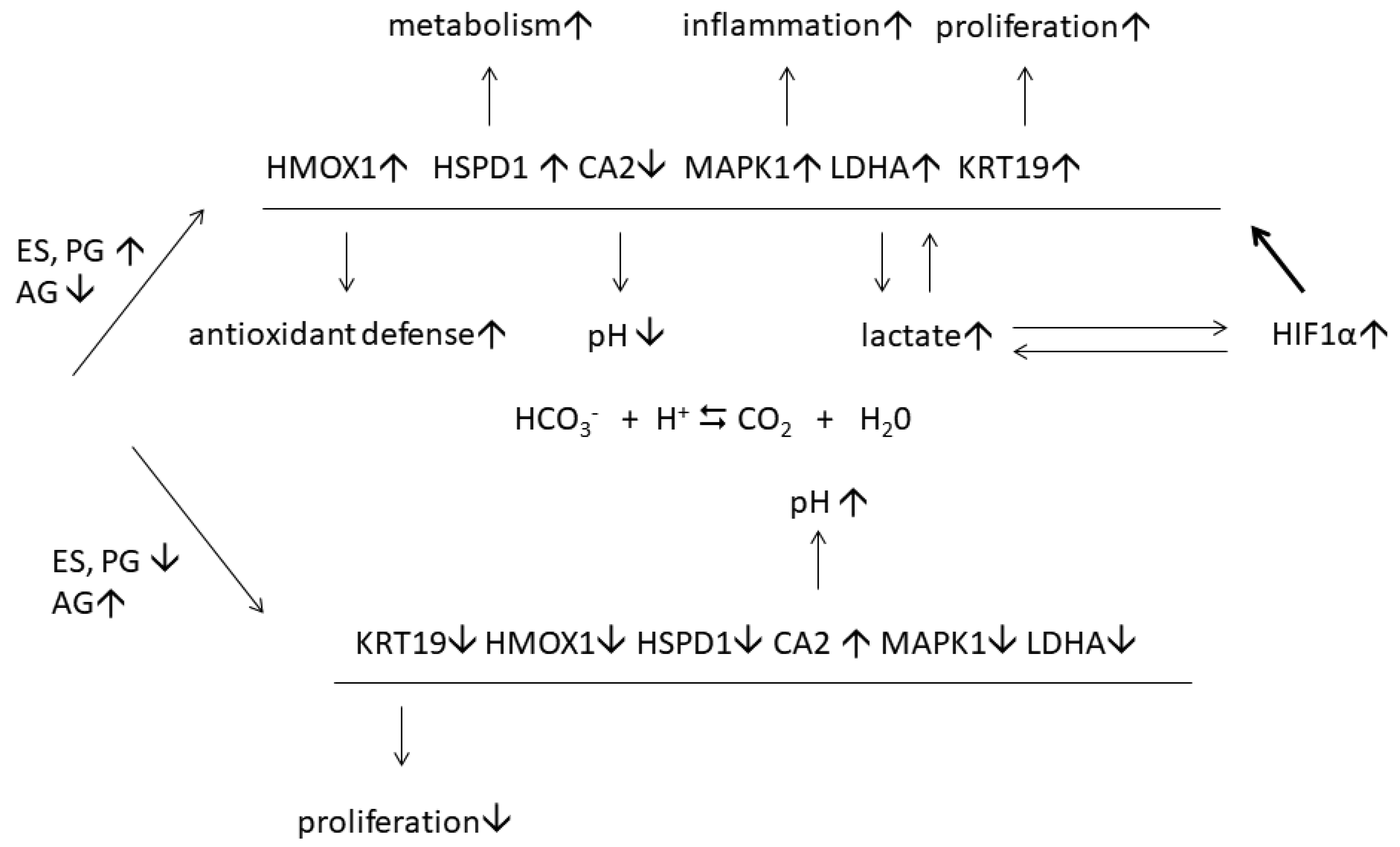
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deng, Y.; Chang, C.; Lu, Q. The Inflammatory Response in Psoriasis: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 50, 377–389. [Google Scholar] [CrossRef]
- Queiro, R.; Tejón, P.; Alonso, S.; Coto, P. Age at disease onset: A key factor for understanding psoriatic disease. Rheumatology 2014, 53, 1178–1185. [Google Scholar] [CrossRef] [Green Version]
- Naldi, L. Epidemiology of psoriasis. Curr. Drug Targets. Inflamm. Allergy 2004, 3, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Murase, J.E.; Chan, K.K.; Garite, T.J.; Cooper, D.M.; Weinstein, G.D. Hormonal effect on psoriasis in pregnancy and post partum. Arch. Dermatol. 2005, 141, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [Green Version]
- Shah, N.M.; Lai, P.F.; Imami, N.; Johnson, M.R. Progesterone-Related Immune Modulation of Pregnancy and Labor. Front. Endocrinol. 2019, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Kurizky, P.S.; Ferreira Cde, C.; Nogueira, L.S.; Mota, L.M. Treatment of psoriasis and psoriatic arthritis during pregnancy and breastfeeding. An. Bras. Dermatol. 2015, 90, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceovic, R.; Mance, M.; Bukvic Mokos, Z.; Svetec, M.; Kostovic, K.; Stulhofer Buzina, D. Psoriasis: Female skin changes in various hormonal stages throughout life—Puberty, pregnancy, and menopause. BioMed Res. Int. 2013, 2013, 571912. [Google Scholar] [CrossRef]
- Lee, Y.H.; Song, G.G. Association between circulating prolactin levels and psoriasis and its correlation with disease severity: A meta-analysis. Clin. Exp. Dermatol. 2018, 43, 27–35. [Google Scholar] [CrossRef]
- Allshouse, A.; Pavlovic, J.; Santoro, N. Menstrual Cycle Hormone Changes Associated with Reproductive Aging and How They May Relate to Symptoms. Obstet. Gynecol. Clin. N. Am. 2018, 45, 613–628. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, M.; Li, Z.; Song, Y. Follicle-Stimulating Hormone Positively Associates with Metabolic Factors in Perimenopausal Women. Int. J. Endocrinol. 2020, 2020, 7024321. [Google Scholar] [CrossRef] [PubMed]
- Dobrohotova, J.E.; Zelenskaia, E.M.; Korsunskaia, I.M. Livial and some aspects of its infuentce on the skin of postmenopausal women. Russ. Bull. Obstet. Gynecol. 2007, 7, 72–76. [Google Scholar]
- Morgulis, J.V.; Potekaev, N.N.; Korsunskaia, I.M. Non-invasive mechods to study the skin of menopausal females and clinical potential of hormone replacing therapy. Russ. J. Clin. Dermatol. Venereol. 2009, 7, 13–18. [Google Scholar]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. MCP 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.P.; Zerbato, J.M.; Zhao, W.; Braat, S.; Deleage, C.; Tennakoon, G.S.; Mason, H.; Dantanarayana, A.; Rhodes, A.; Rhodes, J.W.; et al. Intrahepatic CXCL10 is strongly associated with liver fibrosis in HIV-Hepatitis B co-infection. PLoS Pathog. 2020, 16, e1008744. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Qu, Y.; Chen, X.; Wen, Z.; Chen, P.; Cheng, Y. Radiotherapy Increases 12-LOX and CCL5 Levels in Esophageal Cancer Cells and Promotes Cancer Metastasis via THP-1-Derived Macrophages. Onco Targets Ther. 2020, 13, 7719–7733. [Google Scholar] [CrossRef]
- Edwards, C.K., 3rd; Green, J.S.; Volk, H.D.; Schiff, M.; Kotzin, B.L.; Mitsuya, H.; Kawaguchi, T.; Sakata, K.M.; Cheronis, J.; Trollinger, D.; et al. Combined anti-tumor necrosis factor-α therapy and DMARD therapy in rheumatoid arthritis patients reduces inflammatory gene expression in whole blood compared to DMARD therapy alone. Front. Immunol. 2012, 3, 366. [Google Scholar] [CrossRef] [Green Version]
- Piccinni, M.P.; Scaletti, C.; Maggi, E.; Romagnani, S. Role of hormone-controlled Th1- and Th2-type cytokines in successful pregnancy. J. Neuroimmunol. 2000, 109, 30–33. [Google Scholar] [CrossRef]
- Kanda, N.; Watanabe, S. Regulatory roles of sex hormones in cutaneous biology and immunology. J. Dermatol. Sci. 2005, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Raghunath, R.S.; Venables, Z.C.; Millington, G.W. The menstrual cycle and the skin. Clin. Exp. Dermatol. 2015, 40, 111–115. [Google Scholar] [CrossRef]
- Numata, I.; Okuyama, R.; Memezawa, A.; Ito, Y.; Takeda, K.; Furuyama, K.; Shibahara, S.; Aiba, S. Functional expression of heme oxygenase-1 in human differentiated epidermis and its regulation by cytokines. J. Investig. Dermatol. 2009, 129, 2594–2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, N.K.; Fitzgerald, H.K.; Malara, A.; Hambly, R.; Sweeney, C.M.; Kirby, B.; Fletcher, J.M.; Dunne, A. Naturally derived Heme-Oxygenase 1 inducers attenuate inflammatory responses in human dendritic cells and T cells: Relevance for psoriasis treatment. Sci. Rep. 2018, 8, 10287. [Google Scholar] [CrossRef] [PubMed]
- Ghoreschi, K.; Brück, J.; Kellerer, C.; Deng, C.; Peng, H.; Rothfuss, O.; Hussain, R.Z.; Gocke, A.R.; Respa, A.; Glocova, I.; et al. Fumarates improve psoriasis and multiple sclerosis by inducing type II dendritic cells. J. Exp. Med. 2011, 208, 2291–2303. [Google Scholar] [CrossRef] [PubMed]
- Keum, H.; Kim, T.W.; Kim, Y.; Seo, C.; Son, Y.; Kim, J.; Kim, D.; Jung, W.; Whang, C.H.; Jon, S. Bilirubin nanomedicine alleviates psoriatic skin inflammation by reducing oxidative stress and suppressing pathogenic signaling. J. Control. Release 2020, 325, 359–369. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Sabio, G.; Davis, R.J. TNF and MAP kinase signalling pathways. Semin. Immunol. 2014, 26, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Madak-Erdogan, Z.; Lupien, M.; Stossi, F.; Brown, M.; Katzenellenbogen, B.S. Genomic collaboration of estrogen receptor alpha and extracellular signal-regulated kinase 2 in regulating gene and proliferation programs. Mol. Cell. Biol. 2011, 31, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.J.; Li, C.Y.; Dai, H.Y.; Cai, D.X.; Wang, K.Y.; Xu, Y.H.; Chen, L.M.; Zhou, C.L. Expression and localization of the activated mitogen-activated protein kinase in lesional psoriatic skin. Exp. Mol. Pathol. 2007, 83, 413–418. [Google Scholar] [CrossRef]
- Urbańska, K.; Orzechowski, A. Unappreciated Role of LDHA and LDHB to Control Apoptosis and Autophagy in Tumor Cells. Int. J. Mol. Sci. 2019, 20, 85. [Google Scholar] [CrossRef] [Green Version]
- Macintyre, A.N.; Gerriets, V.A.; Nichols, A.G.; Michalek, R.D.; Rudolph, M.C.; Deoliveira, D.; Anderson, S.M.; Abel, E.D.; Chen, B.J.; Hale, L.P.; et al. The glucose transporter Glut1 is selectively essential for CD4 T cell activation and effector function. Cell Metab. 2014, 20, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, R.; Smith, J.; Rocher-Ros, V.; Nadkarni, S.; Montero-Melendez, T.; D’Acquisto, F.; Bland, E.J.; Bombardieri, M.; Pitzalis, C.; Perretti, M.; et al. Lactate Regulates Metabolic and Pro-inflammatory Circuits in Control of T Cell Migration and Effector Functions. PLoS Biol. 2015, 13, e1002202. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Kim, A.; Kim, G.T.; Yu, H.Y.; Lee, E.S.; Park, M.J.; Kim, Y.J.; Shim, S.M.; Park, T.S. Inhibition of lactate dehydrogenase A suppresses inflammatory response in RAW 264.7 macrophages. Mol. Med. Rep. 2019, 19, 629–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucino, V.; Certo, M.; Bulusu, V.; Cucchi, D.; Goldmann, K.; Pontarini, E.; Haas, R.; Smith, J.; Headland, S.E.; Blighe, K.; et al. Lactate Buildup at the Site of Chronic Inflammation Promotes Disease by Inducing CD4(+) T Cell Metabolic Rewiring. Cell Metab. 2019, 30, 1055–1074.e1058. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Jiang, K.; Hong, R.; Xu, F.; Xia, W.; Qin, G.; Lee, K.; Zheng, Q.; Lu, Q.; Zhai, Q.; et al. Identification and characterization of critical genes associated with tamoxifen resistance in breast cancer. PeerJ 2020, 8, e10468. [Google Scholar] [CrossRef]
- Saha, S.K.; Kim, K.; Yang, G.M.; Choi, H.Y.; Cho, S.G. Cytokeratin 19 (KRT19) has a Role in the Reprogramming of Cancer Stem Cell-Like Cells to Less Aggressive and More Drug-Sensitive Cells. Int. J. Mol. Sci. 2018, 19, 1423. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.K.; Choi, H.Y.; Kim, B.W.; Dayem, A.A.; Yang, G.M.; Kim, K.S.; Yin, Y.F.; Cho, S.G. KRT19 directly interacts with β-catenin/RAC1 complex to regulate NUMB-dependent NOTCH signaling pathway and breast cancer properties. Oncogene 2017, 36, 332–349. [Google Scholar] [CrossRef] [Green Version]
- Ju, J.H.; Yang, W.; Lee, K.M.; Oh, S.; Nam, K.; Shim, S.; Shin, S.Y.; Gye, M.C.; Chu, I.S.; Shin, I. Regulation of cell proliferation and migration by keratin19-induced nuclear import of early growth response-1 in breast cancer cells. Clin. Cancer Res. 2013, 19, 4335–4346. [Google Scholar] [CrossRef] [Green Version]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Pennell, L.M.; Galligan, C.L.; Fish, E.N. Sex affects immunity. J. Autoimmun. 2012, 38, J282–J291. [Google Scholar] [CrossRef]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Yi, J.M.; Kwon, H.Y.; Cho, J.Y.; Lee, Y.J. Estrogen and hypoxia regulate estrogen receptor alpha in a synergistic manner. Biochem. Biophys. Res. Commun. 2009, 378, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; AlTahan, A.; Jones, D.T.; Buffa, F.M.; Bridges, E.; Interiano, R.B.; Qu, C.; Vogt, N.; Li, J.L.; Baban, D.; et al. Estrogen receptor-α directly regulates the hypoxia-inducible factor 1 pathway associated with antiestrogen response in breast cancer. Proc. Natl. Acad. Sci. USA 2015, 112, 15172–15177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannou, M.; Sourli, F.; Mylonis, I.; Barbanis, S.; Papamichali, R.; Kouvaras, E.; Zafiriou, E.; Siomou, P.; Klimi, E.; Simos, G.; et al. Increased HIF-1 alpha immunostaining in psoriasis compared to psoriasiform dermatitides. J. Cutan. Pathol. 2009, 36, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- De Saedeleer, C.J.; Copetti, T.; Porporato, P.E.; Verrax, J.; Feron, O.; Sonveaux, P. Lactate activates HIF-1 in oxidative but not in Warburg-phenotype human tumor cells. PLoS ONE 2012, 7, e46571. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, D.; Han, S.; Wang, N.; Mo, F.; Loo, T.Y.; Shen, J.; Huang, H.; Chen, J. Bioactivity-guided identification and cell signaling technology to delineate the lactate dehydrogenase A inhibition effects of Spatholobus suberectus on breast cancer. PLoS ONE 2013, 8, e56631. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, G.; Zhu, H.; Dong, X.; Zhao, D.; Jiang, X.; Li, J.; Qiao, H.; Ni, S.; Sun, X. 2-Methoxyestradiol synergizes with sorafenib to suppress hepatocellular carcinoma by simultaneously dysregulating hypoxia-inducible factor-1 and -2. Cancer Lett. 2014, 355, 96–105. [Google Scholar] [CrossRef]
- Wairagu, P.M.; Phan, A.N.; Kim, M.K.; Han, J.; Kim, H.W.; Choi, J.W.; Kim, K.W.; Cha, S.K.; Park, K.H.; Jeong, Y. Insulin priming effect on estradiol-induced breast cancer metabolism and growth. Cancer Biol. Ther. 2015, 16, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.; Azevedo, A.; Nogueira, M.; Torres, T. Management of psoriasis in pregnancy—A review of the evidence to date. Drugs Context 2020, 9. [Google Scholar] [CrossRef] [Green Version]
- De Simone, C.; Caldarola, G.; Moretta, G.; Piscitelli, L.; Ricceri, F.; Prignano, F. Moderate-to-severe psoriasis and pregnancy: Impact on fertility, pregnancy outcome and treatment perspectives. G. Ital. Dermatol. Venereol. 2019, 154, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Hall, G.; Phillips, T.J. Estrogen and skin: The effects of estrogen, menopause, and hormone replacement therapy on the skin. J. Am. Acad. Dermatol. 2005, 53, 555–568, quiz 569–572. [Google Scholar] [CrossRef]
- Rozenberg, S.; Al-Daghri, N.; Aubertin-Leheudre, M.; Brandi, M.L.; Cano, A.; Collins, P.; Cooper, C.; Genazzani, A.R.; Hillard, T.; Kanis, J.A.; et al. Is there a role for menopausal hormone therapy in the management of postmenopausal osteoporosis? Osteoporos. Int. 2020, 31, 2271–2286. [Google Scholar] [CrossRef] [PubMed]
- Sridevi, V.; Naveen, P.; Karnam, V.S.; Reddy, P.R.; Arifullah, M. Beneficiary and Adverse Effects of Phytoestrogens: A Potential Constituent of Plant-based Diet. Curr. Pharm. Des. 2021, 27, 802–815. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Age | Medical History | ID | Age | Medical History |
|---|---|---|---|---|---|
| LC-MS male patients | qPCR/ELISA patients | ||||
| 1 | 30 | not reported | with menopause | ||
| 2 | 40 | stage 1 arterial hyperten- | 1 | 46 | gastritis, cholecystitis, |
| sion, hyperuricemia, | pancreatitis, | ||||
| obesity, | hypertension, coronary | ||||
| 3 | 68 | psoriatic arthritis, | artery disease, angina | ||
| hypertension, stage 2, | pectoris, urolithiasis | ||||
| bronchitis, dyscirculatory | 2 | 46 | hypertension, stage 2, | ||
| encephalopathy, | kidney cyst | ||||
| hyperuricemia, obesity | 3 | 47 | cholecystitis, | ||
| 1st degree | cholelithiasis | ||||
| 4 | 48 | type II diabetes, obesity | |||
| LC-MS female patients | stage 3, | ||||
| 1 | 48 | stage 2 hypertension | arterial hypertension, no- | ||
| 2 | 60 | stage 2 hypertension | dular Hashimoto’s thy- | ||
| roiditis, uterine fibroids, | |||||
| LC-MS male volunteers | cerebrovascular | ||||
| 1 | 53 | phlebeurysm | disease, discirculatory | ||
| 2 | 77 | arthrosclerosis | encephalopathy arte- | ||
| rial hypertension | |||||
| LC-MS female volunteers | 5 | 51 | tachycardia | ||
| 1 | 39 | white line hernia | nodular Hashimoto’s | ||
| 2 | 60 | incisional ventral hernia | thyroiditis, | ||
| 3 | 79 | arthrosclerosis, abdomi- | osteochondrosis | ||
| nal aortic aneurysm, | dorsopathy | ||||
| chronic pyelonephritis, | 6 | 51 | hypertension, stage 2; | ||
| dyslipidemia, | 7 | 53 | hypertension, stage 2; | ||
| hypertension | depression; | ||||
| 8 | 53 | nodular goiter; | |||
| qPCR/ELISA patients, | 9 | 53 | hypertension, stage 2 | ||
| without menopause | type 2 diabetes, osteo- | ||||
| 1 | 19 | not reported | arthritis; dorsopathy; | ||
| 2 | 21 | erysipelas, obesity stage | 10 | 54 | not reported |
| 4, cholecystitis, uterine | qPCR/ELISA volunteers | ||||
| fibroids | without menopause | ||||
| 3 | 28 | not reported | 1 | 26 | not reported |
| 4 | 30 | goiter, euthyroid sick | 2 | 28 | not reported |
| syndrome, | 3 | 29 | not reported | ||
| 5 | 32 | bilateral otitis media, | 4 | 32 | not reported |
| cholecystitis | 5 | 33 | not reported | ||
| vaginal yeast infection | 6 | 38 | not reported | ||
| 6 | 32 | not reported | |||
| 7 | 32 | diffuse goiter, grade 2; | qPCR/ELISA volunteers | ||
| 8 | 36 | depression | with menopause | ||
| 9 | 42 | gastritis, pyelonephritis; | |||
| insulin resistance, | 1 | 43 | not reported | ||
| hypertension, stage 2, | 2 | 49 | not reported | ||
| cerebrovascular disease, | 3 | 50 | not reported | ||
| cholecystitis | 4 | 55 | not reported | ||
| 10 | 43 | not reported | 5 | 57 | not reported |
| Gene | Reference Sequence | Primer Name | Primer Sequence | Product Size, bp |
|---|---|---|---|---|
| CA2 | NM_000067.3 | CA2 forward | GGCTGGTTGGTGCTTTGTTT | 118 |
| CA2 re-verse | TTGTGAGTGCTCATCACCCT | |||
| HMOX1 | NM_002133.3 | HMOX1 forward | GGCCTAAACTTCAGAGGGGG | 99 |
| HMOX1 reverse | AGACAGCTGCCACATTAGGG | |||
| HSPD1 | NM_002156.5 | HSPD1 forward | CTGGCACGCTCTATAGCCAA | 142 |
| HSPD1 reverse | CAGGGGTGGTCACAGGTTTA | |||
| KRT19 | NM_002276.5 | KRT19 forward | CCACTACTACACGACCATCCA | 89 |
| KRT19 reverse | GTCGATCTGCAGGACAATCC | |||
| LDHA | NM_005566.4 | LDHA forward | TAAGCTGTCATGGGTGGGTC | 100 |
| LDHA reverse | GGGTGCAGAGTCTTCAGAGAG | |||
| MAPK1 | NM_002745.5 | MAPK1 forward | CAGTTCTTGACCCCTGGTCC | 186 |
| MAPK1 reverse | TACATACTGCCGCAGGTCAC | |||
| 18S RNA | NR_003286.2 | 18S RNA forward | CTACCACATCCAAGGAAGCA | 103 |
| 18S RNA reverse | TTTTTCGTCACTACCTCCCCG |
| Group of Participants | N | Estradiol, pM | Progesterone, nM | Testosterone, nM |
|---|---|---|---|---|
| Volunteers without | 6 | 480.63 ± 157.43 | 8.18 ± 2.47 | 0.406 ± 0.078 |
| menopause | ||||
| Volunteers with | 5 | 53.8 ± 15.91 | 0.973 ± 0.206 | 0.733 ± 0.061 |
| menopause | ||||
| p-value | 0.026 | 0.018 | 0.018 | |
| Patients without | 10 | 116.32 ± 45.85 | 0.159 ± 0.013 | 1.708 ± 0.362 |
| menopause | ||||
| Patients with | 10 | 14.51 ± 0.44 | 0.110 ± 0.011 | 3.428 ± 0.368 |
| menopause | ||||
| p-value | 0.0495 | 0.011 | 0.005 |
| Term | Genes | p-Value | FDR |
|---|---|---|---|
| Translational initiation | RPL30, RPL10, RPS7, RPS8, RPL11, RPL13A, RPL23A, RPS15, RPS27, RPS19, RPL14, EIF3C, RPL28, EIF3D, EIF4G1, RPS12 | 8.59 × 10−15 | 6.62 × 10−12 |
| Nuclear-transcribed mRNA catabolic process, nonsense- mediated decay | RPL30, RPL10, RPS7, RPS8, RPL11, RPL13A, RPL23A, RPS15, RPS27, RPS19, RPL14, RPL28, EIF4G1, RPS12 | 6.03 × 10−13 | 1.89 × 10−10 |
| SRP-dependent cotranslational protein targeting to membrane | RPL30, RPL10, RPS7, RPS8, RPL11, RPL13A, RPL23A, RPS15, RPS27, RPS19, RPL14, RPL28, RPS12 | 7.37 × 10−13 | 1.89 × 10−10 |
| Viral transcription | RPL30, RPL10, RPS7, RPS8, RPL11, RPL13A, RPL23A, RPS15, RPS27, RPS19, RPL14, RPL28, RPS12 | 6.20 × 10−12 | 1.19 × 10−9 |
| Translation | RPL30, RPL10, RPS7, RPS8, RPL11, RPL13A, SARS, RPL23A, RPS15, RPS27, RPS19, RPL14, RPL28, EIF4G1, RPS12 | 7.59 × 10−10 | 1.17 × 10−7 |
| rRNA processing | RPL30, RPL10, RPS7, RPS8, RPL11, RPL13A, DDX21, RPL23A, RPS15, RPS27, RPS19, RPL14, RPL28, RPS12 | 1.03 × 10−9 | 1.32 × 10−7 |
| Cell-cell adhesion | LDHA, AHNAK, HSPA5, RPL14, TACSTD2, EFHD2, RPL23A, TAGLN2, ENO1, ALDOA, SPTBN2, EIF4G1 | 1.24 × 10−6 | 1.37 × 10−4 |
| Regulation of mRNA stability | PSMD6, PSMD7, PSMD13, PSMC1, PSMD3, HSPB1, HSPA1B, EIF4G1 | 4.20 × 10−6 | 4.04 × 10−4 |
| Response to estrogen | LDHA, KRT19, CA2, HMOX1, MAPK1, HSPD1 | 5.83 × 10−5 | 4.81 × 10−3 |
| Glycolytic process | GPI, LDHA, PGAM1, ENO1, ALDOA | 6.24 × 10−5 | 4.81 × 10−3 |
| Regulation of cellular amino acid metabolic process | PSMD6, PSMD7, PSMD13, PSMC1, PSMD3 | 3.09 × 10−4 | 0.022 |
| Canonical glycolysis | GPI, PGAM1, ENO1, ALDOA | 5.88 × 10−4 | 0.038 |
| Antigen processing and presentation of exogenous peptide antigen via MHC class I, TAP-dependent | PSMD6, PSMD7, PSMD13, PSMC1, PSMD3 | 6.95 × 10−4 | 0.041 |
| NIK/NF-κB signaling | PSMD6, PSMD7, PSMD13, PSMC1, PSMD3 | 8.29 × 10−4 | 0.046 |
| Term | Genes | p-Value | FDR |
|---|---|---|---|
| SRP-dependent cotran- slational protein targe- ting to membrane | RPS28, RPS16, RPL32, RPL23, RPL37A, RPL35A, FAU, RPL8, SRP14, RPL17, RPL19 | 4.74 × 10−9 | 4.95 × 10−6 |
| Translational initiation | RPS28, RPS16, RPL32, EIF6, RPL23, RPL37A, RPL35A, FAU, RPL8, RPL17, RPL19 | 1.79 × 10−7 | 9.32 × 10−5 |
| Viral transcription | RPS28, RPS16, RPL32, RPL23, RPL37A, RPL35A, FAU, RPL8, RPL17, RPL19 | 3.32 × 10−7 | 1.16 × 10−4 |
| Nuclear-transcribed mRNA catabolic process, nonsense- mediated decay | RPS28, RPS16, RPL32, RPL23, RPL37A, RPL35A, FAU, RPL8, RPL17, RPL19 | 5.57 × 10−7 | 1.45 × 10−4 |
| Translation | RPS28, RPS16, RPL32, RPL23, RPL37A, RPL35A, FAU, RPL8, RPL17, SLC25A6, RPL19 | 4.33 × 10−5 | 0.009 |
| Negative regulation of endopeptidase activity | CSTB, CSTA, ITIH2, SERPIND1, SERPINF1, SERPINH1, SERPING1, COL6A3 | 6.01 × 10−5 | 0.010 |
| rRNA processing | RPS28, RPS16, RPL32, RPL23, RPL37A, RPL35A, FAU, RPL8, RPL17, RPL19 | 6.56 × 10−5 | 0.010 |
| Regulation of comple- ment activation | CFH, C9, C8B, PHB2, C8A | 9.86 × 10−5 | 0.013 |
| Complement activation alternative pathway | CFH, C9, C8B, C8A | 1.43 × 10−4 | 0.017 |
| Cell-cell adhesion | CNN2, PDLIM1, LAD1, DDX3X, ATIC, CTTN, RUVBL1, CHMP4B, PARK7, CNN3 | 3.86 × 10−4 | 0.040 |
| DNA duplex unwinding | DDX3X, XRCC5, DDX1, RUVBL2, RUVBL1 | 4.47 × 10−4 | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobolev, V.; Soboleva, A.; Denisova, E.; Denieva, M.; Dvoryankova, E.; Suleymanov, E.; Zhukova, O.V.; Potekaev, N.; Korsunskaya, I.; Mezentsev, A. Differential Expression of Estrogen-Responsive Genes in Women with Psoriasis. J. Pers. Med. 2021, 11, 925. https://doi.org/10.3390/jpm11090925
Sobolev V, Soboleva A, Denisova E, Denieva M, Dvoryankova E, Suleymanov E, Zhukova OV, Potekaev N, Korsunskaya I, Mezentsev A. Differential Expression of Estrogen-Responsive Genes in Women with Psoriasis. Journal of Personalized Medicine. 2021; 11(9):925. https://doi.org/10.3390/jpm11090925
Chicago/Turabian StyleSobolev, Vladimir, Anna Soboleva, Elena Denisova, Malika Denieva, Eugenia Dvoryankova, Elkhan Suleymanov, Olga V. Zhukova, Nikolay Potekaev, Irina Korsunskaya, and Alexandre Mezentsev. 2021. "Differential Expression of Estrogen-Responsive Genes in Women with Psoriasis" Journal of Personalized Medicine 11, no. 9: 925. https://doi.org/10.3390/jpm11090925
APA StyleSobolev, V., Soboleva, A., Denisova, E., Denieva, M., Dvoryankova, E., Suleymanov, E., Zhukova, O. V., Potekaev, N., Korsunskaya, I., & Mezentsev, A. (2021). Differential Expression of Estrogen-Responsive Genes in Women with Psoriasis. Journal of Personalized Medicine, 11(9), 925. https://doi.org/10.3390/jpm11090925