
Sex-Related Differences of Matrix Metalloproteinases (MMPs): New Perspectives for These Biomarkers in Cardiovascular and Neurological Diseases

 ,
, 
 and
and
Abstract
:1. Introduction
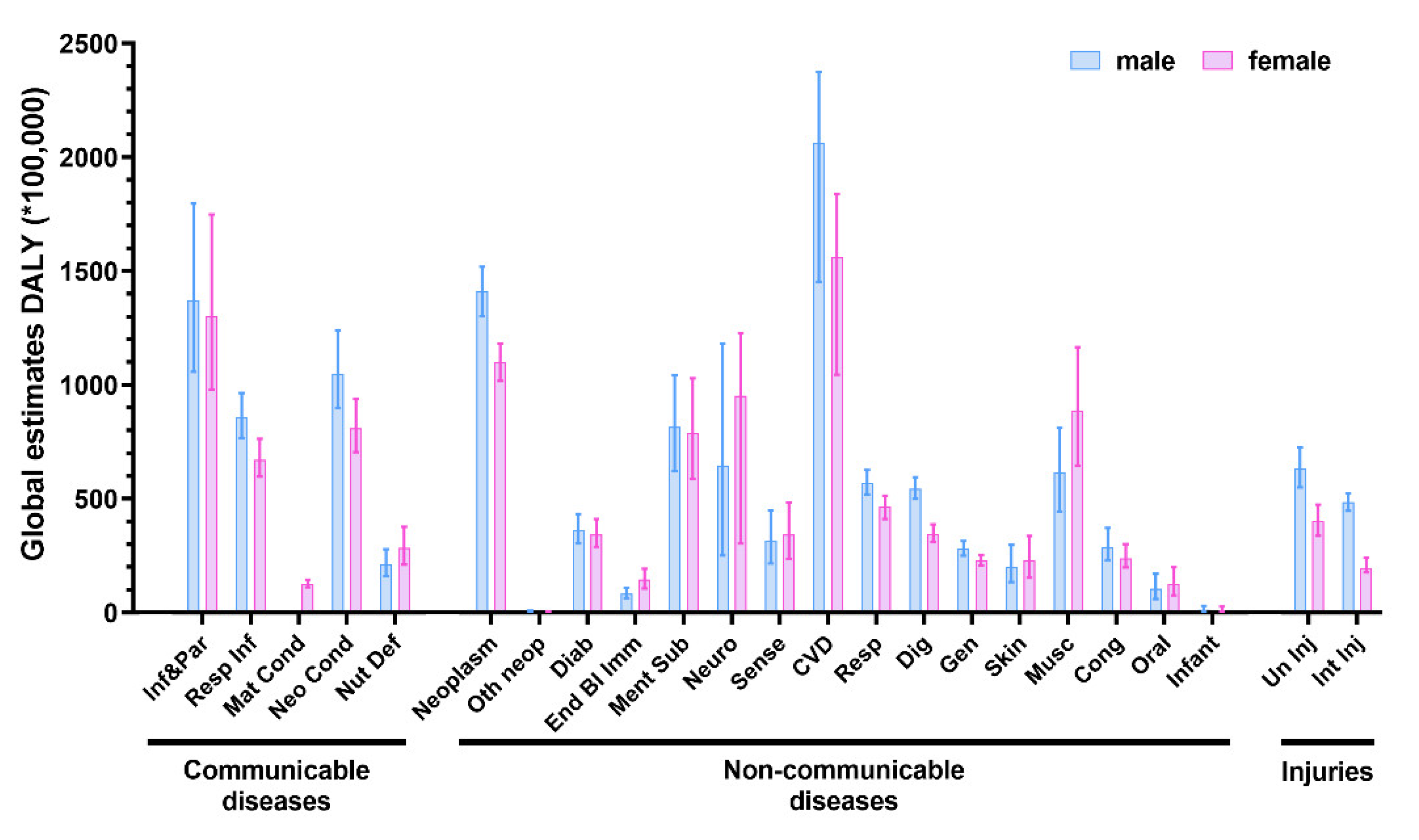
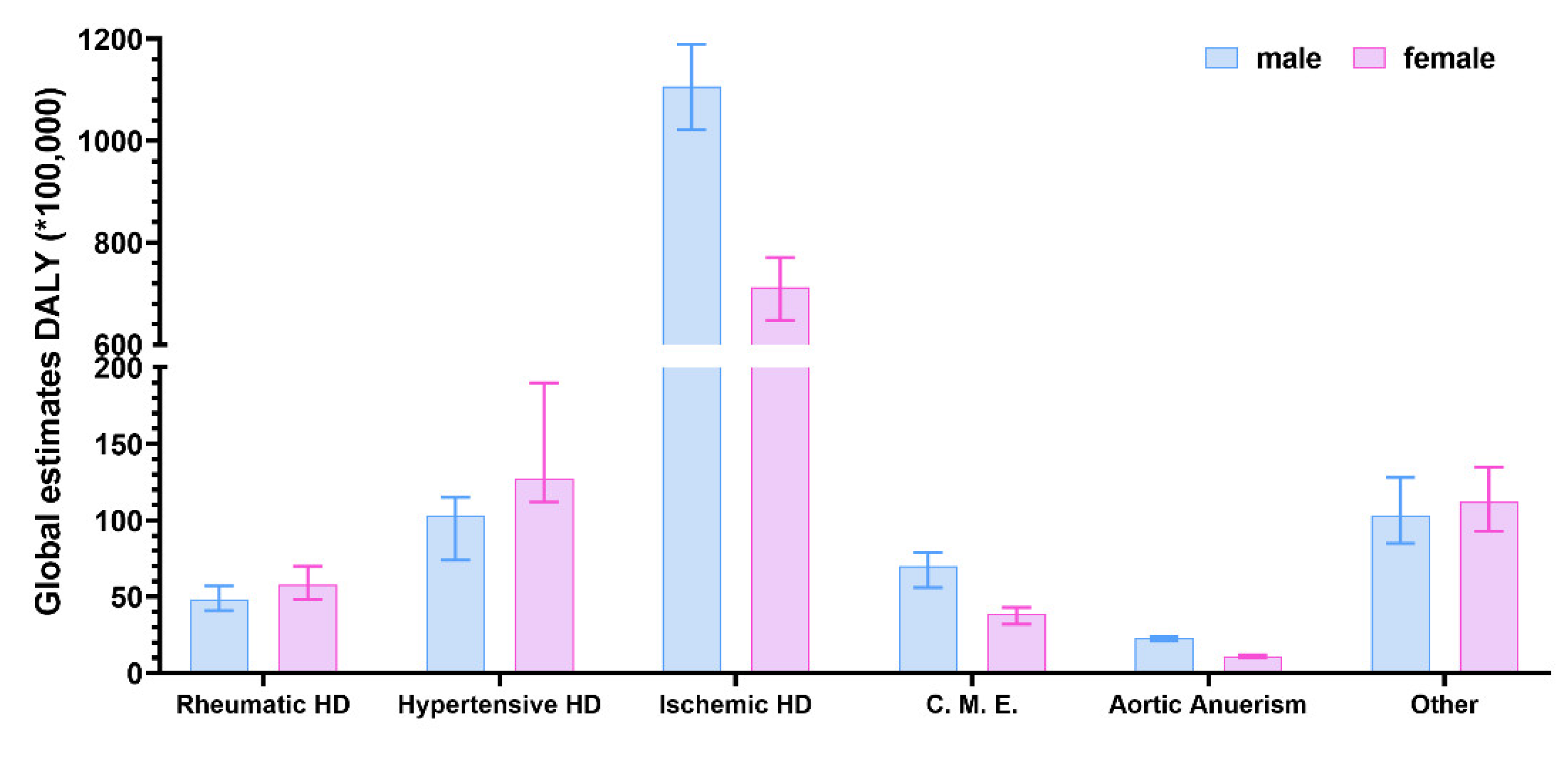
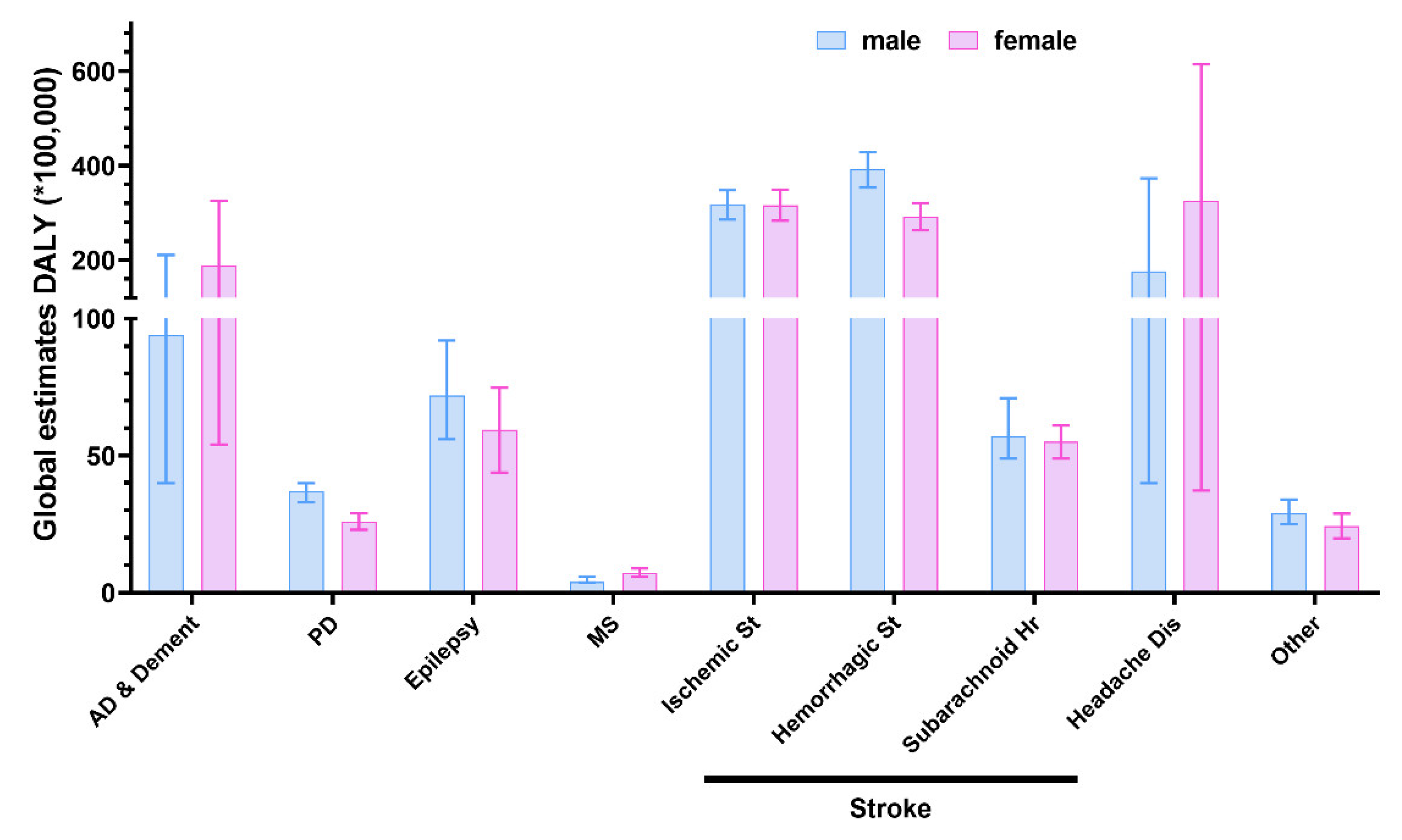
2. Search Strategy and Data
3. Matrix Metalloproteinases
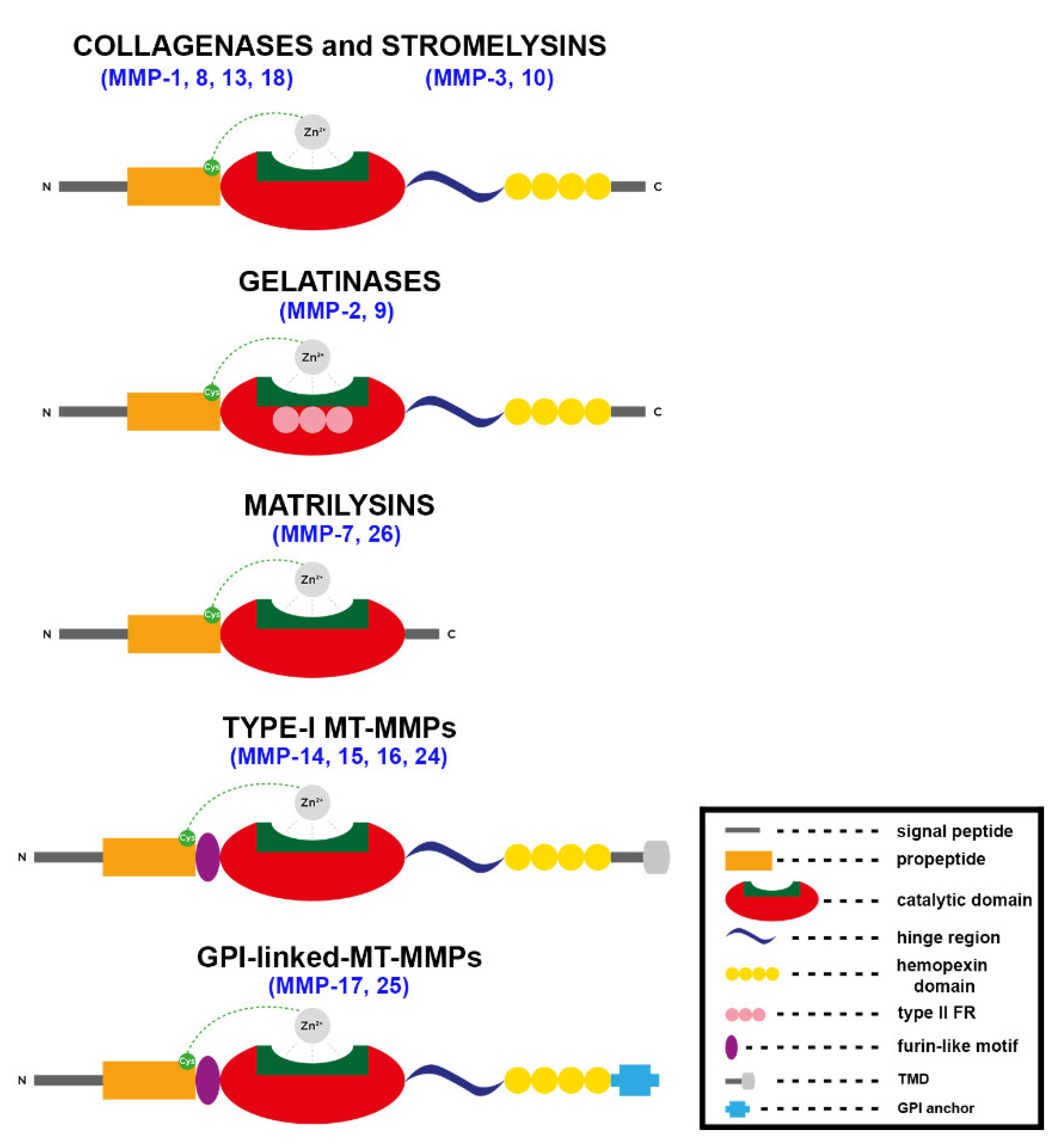
3.1. Structure and Function of MMPs
3.1.1. General Structure and Regulation
3.1.2. Role of MMPs in CVDs
3.1.3. Role of MMPs in Neurological Conditions
4. Sex Differences in MMPs and Pathologies
4.1. Sex Differences in MMPs and Cardiovascular Diseases
4.2. Sex Differences in MMPs and CNS Disorders
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ngun, T.C.; Ghahramani, N.; Sánchez, F.J.; Bocklandt, S.; Vilain, E. The Genetics of Sex Differences in Brain and Behavior. Front. Neuroendocrinol. 2011, 32, 227–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGregor, A.J.; Hasnain, M.; Sandberg, K.; Morrison, M.F.; Berlin, M.; Trott, J. How to Study the Impact of Sex and Gender in Medical Research: A Review of Resources. Biol. Sex Differ. 2016, 7, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oertelt-Prigione, S.; Dalibert, L.; Verdonk, P.; Stutz, E.Z.; Klinge, I. Implementation Strategies for Gender-Sensitive Public Health Practice: A European Workshop. J. Women’s Health 2017, 26, 1255–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemmati, D.; Varani, K.; Bramanti, B.; Piva, R.; Bonaccorsi, G.; Trentini, A.; Manfrinato, M.C.; Tisato, V.; Carè, A.; Bellini, T. “Bridging the Gap” Everything That Could Have Been Avoided If We Had Applied Gender Medicine, Pharmacogenetics and Personalized Medicine in the Gender-Omics and Sex-Omics Era. Int. J. Mol. Sci. 2019, 21, 296. [Google Scholar] [CrossRef] [Green Version]
- Feigin, V.L.; Vos, T. Global Burden of Neurological Disorders: From Global Burden of Disease Estimates to Actions. Neuroepidemiology 2019, 52, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.; Thomas, E.E.; Tan, J.T.M.; Inglis, S.C.; Wu, J.H.Y.; Climie, R.E.; Picone, D.S.; Blekkenhorst, L.C.; Wise, S.G.; Mirabito Colafella, K.M.; et al. A Roadmap of Strategies to Support Cardiovascular Researchers: From Policy to Practice. Nat. Rev. Cardiol. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Ramsey, J.M.; Cooper, J.D.; Penninx, B.W.J.H.; Bahn, S. Variation in Serum Biomarkers with Sex and Female Hormonal Status: Implications for Clinical Tests. Sci. Rep. 2016, 6, 26947. [Google Scholar] [CrossRef]
- Chhabra, A.; Kohli, S.; Rani, V. MMPs in Cardiovascular Diseases: Emerging Pharmacological Targets. In Role of Proteases in Cellular Dysfunction; Springer: New York, NY, USA, 2014; pp. 407–426. [Google Scholar]
- Rosenberg, G. Metalloproteinases and Neurodegenerative Diseases: Pathophysiological and Therapeutic Perspectives. Metalloproteinases Med. 2015, 2, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Rempe, R.G.; Hartz, A.M.S.; Bauer, B. Matrix Metalloproteinases in the Brain and Blood-Brain Barrier: Versatile Breakers and Makers. J. Cereb. Blood Flow Metab. 2016, 36, 1481–1507. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zheng, M.; Betancourt, C.E.; Liu, L.; Sitikov, A.; Sladojevic, N.; Zhao, Q.; Zhang, J.H.; Liao, J.K.; Wu, R. Increase in Blood-Brain Barrier (BBB) Permeability Is Regulated by MMP3 via the ERK Signaling Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 6655122. [Google Scholar] [CrossRef]
- Olejarz, W.; Łacheta, D.; Kubiak-Tomaszewska, G. Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability. Int. J. Mol. Sci. 2020, 21, 3946. [Google Scholar] [CrossRef] [PubMed]
- Thrailkill, K.M.; Moreau, C.S.; Cockrell, G.; Simpson, P.; Goel, R.; North, P.; Fowlkes, J.L.; Bunn, R.C. Physiological Matrix Metalloproteinase Concentrations in Serum during Childhood and Adolescence, Using Luminex Multiplex Technology. Clin. Chem. Lab. Med. 2005, 43, 1392–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, L.G.N.; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. Matrix Metalloproteinases: From Molecular Mechanisms to Physiology, Pathophysiology, and Pharmacology. Pharmacol. Rev. 2022, 74, 712–768. [Google Scholar] [CrossRef]
- Loffek, S.; Schilling, O.; Franzke, C.-W. Biological Role of Matrix Metalloproteinases: A Critical Balance. Eur. Respir. J. 2011, 38, 191–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.B.; Stephens, R.W.; Brünner, N.; Nielsen, H.J.; Engelholm, L.H.; Christensen, I.J.; Stetler-Stevenson, W.G.; Høyer-Hansen, G. Quantification of Tissue Inhibitor of Metalloproteinases 2 in Plasma from Healthy Donors and Cancer Patients. Scand. J. Immunol. 2005, 61, 449–460. [Google Scholar] [CrossRef]
- Burrage, P.S.; Mix, K.S.; Brinckerhoff, C.E. Matrix Metalloproteinases: Role in Arthritis. Front. Biosci. 2006, 11, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Roldán, V.; Marín, F.; Gimeno, J.R.; Ruiz-Espejo, F.; González, J.; Feliu, E.; García-Honrubia, A.; Saura, D.; de la Morena, G.; Valdés, M.; et al. Matrix Metalloproteinases and Tissue Remodeling in Hypertrophic Cardiomyopathy. Am. Heart J. 2008, 156, 85–91. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Winsz-Szczotka, K.; Kuznik-Trocha, K.; Klimek, K.; Olczyk, K. Age- and Gender-Dependent Changes in Connective Tissue Remodeling: Physiological Differences in Circulating MMP-3, MMP-10, TIMP-1 and TIMP-2 Level. Gerontology 2011, 57, 44–52. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, G.; Sehgal, A.; Bhardwaj, S.; Singh, S.; Buhas, C.; Judea-Pusta, C.; Uivarosan, D.; Munteanu, M.A.; Bungau, S. Multifaceted Role of Matrix Metalloproteinases in Neurodegenerative Diseases: Pathophysiological and Therapeutic Perspectives. Int. J. Mol. Sci. 2021, 22, 1413. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and Function of Matrix Metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.P.; Hansch, C. Matrix Metalloproteinases (MMPs): Chemical-Biological Functions and (Q)SARs. Bioorg. Med. Chem. 2007, 15, 2223–2268. [Google Scholar] [CrossRef] [PubMed]
- Paiva, K.B.S.; Granjeiro, J.M. Bone Tissue Remodeling and Development: Focus on Matrix Metalloproteinase Functions. Arch. Biochem. Biophys. 2014, 561, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, C.; Vaidya, S.; Wadhwan, V.; Hitesh; Kaur, G.; Pathak, A. Seesaw of Matrix Metalloproteinases (MMPs). J. Cancer Res. Ther. 2016, 12, 28–35. [Google Scholar] [CrossRef]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix Metalloproteinases (MMPs), the Main Extracellular Matrix (ECM) Enzymes in Collagen Degradation, as a Target for Anticancer Drugs. J. Enzyme. Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef] [Green Version]
- van Wart, H.E.; Birkedal-Hansen, H. The Cysteine Switch: A Principle of Regulation of Metalloproteinase Activity with Potential Applicability to the Entire Matrix Metalloproteinase Gene Family. Proc. Natl. Acad. Sci. USA 1990, 87, 5578–5582. [Google Scholar] [CrossRef] [Green Version]
- Bode, W.; Gomis-Rüth, F.X.; Stöckler, W. Astacins, Serralysins, Snake Venom and Matrix Metalloproteinases Exhibit Identical Zinc-Binding Environments (HEXXHXXGXXH and Met-Turn) and Topologies and Should Be Grouped into a Common Family, the “Metzincins”. FEBS Lett. 1993, 331, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [Green Version]
- Jaoude, J.; Koh, Y. Matrix Metalloproteinases in Exercise and Obesity. Vasc Health Risk Manag 2016, 12, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhard, U.; Huesgen, P.F.; Schilling, O.; Bellac, C.L.; Butler, G.S.; Cox, J.H.; Dufour, A.; Goebeler, V.; Kappelhoff, R.; Keller, U.A.; et al. Active Site Specificity Profiling of the Matrix Metalloproteinase Family: Proteomic Identification of 4300 Cleavage Sites by Nine MMPs Explored with Structural and Synthetic Peptide Cleavage Analyses. Matrix Biol. 2016, 49, 37–60. [Google Scholar] [CrossRef]
- Ra, H.-J.; Parks, W.C. Control of Matrix Metalloproteinase Catalytic Activity. Matrix Biol. 2007, 26, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Cauwe, B.; Opdenakker, G. Intracellular Substrate Cleavage: A Novel Dimension in the Biochemistry, Biology and Pathology of Matrix Metalloproteinases. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 351–423. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Boyd, D.D. Regulation of Matrix Metalloproteinase Gene Expression. J. Cell. Physiol. 2007, 211, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Arpino, V.; Brock, M.; Gill, S.E. The Role of TIMPs in Regulation of Extracellular Matrix Proteolysis. Matrix Biol. 2015, 44, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Roderfeld, M.; Graf, J.; Giese, B.; Salguero-Palacios, R.; Tschuschner, A.; Müller-Newen, G.; Roeb, E. Latent MMP-9 Is Bound to TIMP-1 before Secretion. BCHM 2007, 388, 1227–1234. [Google Scholar] [CrossRef]
- Ardi, V.C.; Kupriyanova, T.A.; Deryugina, E.I.; Quigley, J.P. Human Neutrophils Uniquely Release TIMP-Free MMP-9 to Provide a Potent Catalytic Stimulator of Angiogenesis. Proc. Natl Acad. Sci. USA 2007, 104, 20262–20267. [Google Scholar] [CrossRef] [Green Version]
- Bellini, T.; Trentini, A.; Manfrinato, M.C.; Tamborino, C.; Volta, C.A.; di Foggia, V.; Fainardi, E.; Dallocchio, F.; Castellazzi, M. Matrix Metalloproteinase-9 Activity Detected in Body Fluids Is the Result of Two Different Enzyme Forms. J. Biochem. 2012, 151, 493–499. [Google Scholar] [CrossRef]
- Trentini, A.; Manfrinato, M.C.; Castellazzi, M.; Tamborino, C.; Roversi, G.; Volta, C.A.; Baldi, E.; Tola, M.R.; Granieri, E.; Dallocchio, F.; et al. TIMP-1 Resistant Matrix Metalloproteinase-9 Is the Predominant Serum Active Isoform Associated with MRI Activity in Patients with Multiple Sclerosis. Mult. Scler. 2015, 21, 1121–1130. [Google Scholar] [CrossRef]
- Alfaddagh, A.; Martin, S.S.; Leucker, T.M.; Michos, E.D.; Blaha, M.J.; Lowenstein, C.J.; Jones, S.R.; Toth, P.P. Inflammation and Cardiovascular Disease: From Mechanisms to Therapeutics. Am. J. Prev. Cardiol. 2020, 4, 100130. [Google Scholar] [CrossRef]
- Shah, P.K.; Falk, E.; Badimon, J.J.; Fernandez-Ortiz, A.; Mailhac, A.; Villareal-Levy, G.; Fallon, J.T.; Regnstrom, J.; Fuster, V. Human Monocyte-Derived Macrophages Induce Collagen Breakdown in Fibrous Caps of Atherosclerotic Plaques. Potential Role of Matrix-Degrading Metalloproteinases and Implications for Plaque Rupture. Circulation 1995, 92, 1565–1569. [Google Scholar]
- Hopps, E.; lo Presti, R.; Caimi, G. Matrix Metalloproteases in Arterial Hypertension and Their Trend after Antihypertensive Treatment. Kidney Blood Press. Res. 2017, 42, 347–357. [Google Scholar] [CrossRef]
- Rizzi, E.; Castro, M.M.; Prado, C.M.; Silva, C.A.; Fazan, R.; Rossi, M.A.; Tanus-Santos, J.E.; Gerlach, R.F. Matrix Metalloproteinase Inhibition Improves Cardiac Dysfunction and Remodeling in 2-Kidney, 1-Clip Hypertension. J. Card. Fail. 2010, 16, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.F.; Tran, E.D.; Fortes, Z.B.; Schmid-Schönbein, G.W. Matrix Metalloproteinases Cleave the β 2 -Adrenergic Receptor in Spontaneously Hypertensive Rats. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H25–H35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Patron, C.; Stewart, K.G.; Zhang, Y.; Koivunen, E.; Radomski, M.W.; Davidge, S.T. Vascular Matrix Metalloproteinase-2-Dependent Cleavage of Calcitonin Gene-Related Peptide Promotes Vasoconstriction. Circ. Res. 2000, 87, 670–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, A.; Oh, H.-R.; Unsworth, E.J.; Bregonzio, C.; Saavedra, J.M.; Stetler-Stevenson, W.G.; Cuttitta, F. Matrix Metalloproteinase-2 Cleavage of Adrenomedullin Produces a Vasoconstrictor out of a Vasodilator. Biochem. J. 2004, 383, 413–418. [Google Scholar] [CrossRef]
- Zhou, H.-Z.; Ma, X.; Gray, M.O.; Zhu, B.; Nguyen, A.P.; Baker, A.J.; Simonis, U.; Cecchini, G.; Lovett, D.H.; Karliner, J.S. Transgenic MMP-2 Expression Induces Latent Cardiac Mitochondrial Dysfunction. Biochem. Biophys. Res. Commun. 2007, 358, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Sawicki, G.; Leon, H.; Sawicka, J.; Sariahmetoglu, M.; Schulze, C.J.; Scott, P.G.; Szczesna-Cordary, D.; Schulz, R. Degradation of Myosin Light Chain in Isolated Rat Hearts Subjected to Ischemia-Reperfusion Injury: A New Intracellular Target for Matrix Metalloproteinase-2. Circulation 2005, 112, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Schulze, C.J.; Suarez-Pinzon, W.L.; Dyck, J.R.B.; Sawicki, G.; Schulz, R. Intracellular Action of Matrix Metalloproteinase-2 Accounts for Acute Myocardial Ischemia and Reperfusion Injury. Circulation 2002, 106, 1543–1549. [Google Scholar] [CrossRef] [Green Version]
- Mata, K.M.; Prudente, P.S.; Rocha, F.S.; Prado, C.M.; Floriano, E.M.; Elias, J.; Rizzi, E.; Gerlach, R.F.; Rossi, M.A.; Ramos, S.G. Combining Two Potential Causes of Metalloproteinase Secretion Causes Abdominal Aortic Aneurysms in Rats: A New Experimental Model. Int. J. Exp. Pathol. 2011, 92, 26–39. [Google Scholar] [CrossRef]
- Knox, J.B.; Sukhova, G.K.; Whittemore, A.D.; Libby, P. Evidence for Altered Balance between Matrix Metalloproteinases and Their Inhibitors in Human Aortic Diseases. Circulation 1997, 95, 205–212. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Regulation of the Inflammatory Response in Cardiac Repair. Circ. Res. 2012, 110, 159–173. [Google Scholar] [CrossRef]
- Kai, H.; Ikeda, H.; Yasukawa, H.; Kai, M.; Seki, Y.; Kuwahara, F.; Ueno, T.; Sugi, K.; Imaizumi, T. Peripheral Blood Levels of Matrix Metalloproteases-2 and -9 Are Elevated in Patients with Acute Coronary Syndromes. J. Am. Coll. Cardiol. 1998, 32, 368–372. [Google Scholar] [CrossRef] [Green Version]
- Falk, E.; Shah, P.K.; Fuster, V. Coronary Plaque Disruption. Circulation 1995, 92, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.C.; Leu, H.B.; Lin, W.T.; Lin, C.P.; Lin, S.J.; Chen, J.W. Plasma Matrix Metalloproteinase-3 Level Is an Independent Prognostic Factor in Stable Coronary Artery Disease. Eur. J. Clin. Invest. 2005, 35, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Kremastiotis, G.; Handa, I.; Jackson, C.; George, S.; Johnson, J. Disparate Effects of MMP and TIMP Modulation on Coronary Atherosclerosis and Associated Myocardial Fibrosis. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Lenglet, S.; Mach, F.; Montecucco, F. Role of Matrix Metalloproteinase-8 in Atherosclerosis. Mediat. Inflamm. 2013, 2013, 659282. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Mingoia, J.T.; Bruce, J.A.; Austin, J.S.; Stroud, R.E.; Escobar, G.P.; McClister, D.M.; Allen, C.M.; Alfonso-Jaume, M.A.; Fini, M.E.; et al. Selective Spatiotemporal Induction of Matrix Metalloproteinase-2 and Matrix Metalloproteinase-9 Transcription after Myocardial Infarction. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H2216–H2228. [Google Scholar] [CrossRef] [Green Version]
- Ducharme, A.; Frantz, S.; Aikawa, M.; Rabkin, E.; Lindsey, M.; Rohde, L.E.; Schoen, F.J.; Kelly, R.A.; Werb, Z.; Libby, P.; et al. Targeted Deletion of Matrix Metalloproteinase-9 Attenuates Left Ventricular Enlargement and Collagen Accumulation after Experimental Myocardial Infarction. J. Clin. Invest. 2000, 106, 55–62. [Google Scholar] [CrossRef]
- Nilsson, L.; Hallén, J.; Atar, D.; Jonasson, L.; Swahn, E. Early Measurements of Plasma Matrix Metalloproteinase-2 Predict Infarct Size and Ventricular Dysfunction in ST-Elevation Myocardial Infarction. Heart 2012, 98, 31–36. [Google Scholar] [CrossRef]
- Kelly, D.; Cockerill, G.; Ng, L.L.; Thompson, M.; Khan, S.; Samani, N.J.; Squire, I.B. Plasma Matrix Metalloproteinase-9 and Left Ventricular Remodelling after Acute Myocardial Infarction in Man: A Prospective Cohort Study. Eur. Heart J. 2007, 28, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Brinsa, T.A.; Dowdy, K.B.; Scott, A.A.; Baskin, J.M.; Deschamps, A.M.; Lowry, A.S.; Escobar, G.P.; Lucas, D.G.; Yarbrough, W.M.; et al. Myocardial Infarct Expansion and Matrix Metalloproteinase Inhibition. Circulation 2003, 107, 618–625. [Google Scholar] [CrossRef] [Green Version]
- Spinale, F.G.; Janicki, J.S.; Zile, M.R. Membrane-Associated Matrix Proteolysis and Heart Failure. Circ. Res. 2013, 112, 195–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lijnen, H.R.; van Hoef, B.; Vanlinthout, I.; Verstreken, M.; Rio, M.C.; Collen, D. Accelerated Neointima Formation after Vascular Injury in Mice with Stromelysin-3 (MMP-11) Gene Inactivation. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2863–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söder, P.-O.; Meurman, J.H.; Jogestrand, T.; Nowak, J.; Söder, B. Matrix Metalloproteinase-9 and Tissue Inhibitor of Matrix Metalloproteinase-1 in Blood as Markers for Early Atherosclerosis in Subjects with Chronic Periodontitis. J. Periodontal Res. 2009, 44, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.M.; Rizzi, E.; Rodrigues, G.J.; Ceron, C.S.; Bendhack, L.M.; Gerlach, R.F.; Tanus-Santos, J.E. Antioxidant Treatment Reduces Matrix Metalloproteinase-2-Induced Vascular Changes in Renovascular Hypertension. Free Radic. Biol. Med. 2009, 46, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Ceron, C.S.; Rizzi, E.; Guimaraes, D.A.; Martins-Oliveira, A.; Cau, S.B.; Ramos, J.; Gerlach, R.F.; Tanus-Santos, J.E. Time Course Involvement of Matrix Metalloproteinases in the Vascular Alterations of Renovascular Hypertension. Matrix Biol. 2012, 31, 261–270. [Google Scholar] [CrossRef]
- Fernandez-Patron, C.; Radomski, M.W.; Davidge, S.T. Vascular Matrix Metalloproteinase-2 Cleaves Big Endothelin-1 Yielding a Novel Vasoconstrictor. Circ. Res. 1999, 85, 906–911. [Google Scholar] [CrossRef]
- Matsusaka, H.; Ide, T.; Matsushima, S.; Ikeuchi, M.; Kubota, T.; Sunagawa, K.; Kinugawa, S.; Tsutsui, H. Targeted Deletion of Matrix Metalloproteinase 2 Ameliorates Myocardial Remodeling in Mice with Chronic Pressure Overload. Hypertension 2006, 47, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.M.; Lau, L.; Yong, V.W. MMPs in the Central Nervous System: Where the Good Guys Go Bad. Semin. Cell Dev. Biol. 2008, 19, 42–51. [Google Scholar] [CrossRef]
- Lopez-Navarro, E.R.; Gutierrez, J. Metalloproteinases and Their Inhibitors in Neurological Disease. Naunyn Schmiedebergs Arch. Pharmacol. 2022, 395, 27–38. [Google Scholar] [CrossRef]
- Gasche, Y.; Soccal, P.M.; Kanemitsu, M.; Copin, J.-C. Matrix Metalloproteinases and Diseases of the Central Nervous System with a Special Emphasis on Ischemic Brain. Front. Biosci. 2006, 11, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Newby, A.C. Dual Role of Matrix Metalloproteinases (Matrixins) in Intimal Thickening and Atherosclerotic Plaque Rupture. Physiol. Rev. 2005, 85, 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arend, W.P.; Dayer, J.M. Inhibition of the Production and Effects of Interleukin-1 and Tumor Necrosis Factor Alpha in Rheumatoid Arthritis. Arthritis Rheum. 1995, 38, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A. Matrix Metalloproteinases and Their Multiple Roles in Neurodegenerative Diseases. Lancet Neurol. 2009, 8, 205–216. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Yang, Y.; Rosenberg, G.A. Diverse Roles of Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases in Neuroinflammation and Cerebral Ischemia. Neuroscience 2009, 158, 983–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-X.; Tan, M.-S.; Yu, J.-T.; Tan, L. Matrix Metalloproteinases and Their Multiple Roles in Alzheimer’s Disease. Biomed. Res. Int. 2014, 2014, 908636. [Google Scholar] [CrossRef]
- Fainardi, E.; Castellazzi, M.; Tamborino, C.; Trentini, A.; Manfrinato, M.C.; Baldi, E.; Tola, M.R.; Dallocchio, F.; Granieri, E.; Bellini, T. Potential Relevance of Cerebrospinal Fluid and Serum Levels and Intrathecal Synthesis of Active Matrix Metalloproteinase-2 (MMP-2) as Markers of Disease Remission in Patients with Multiple Sclerosis. Mult. Scler. 2009, 15, 547–554. [Google Scholar] [CrossRef]
- Choi, D.H.; Kim, E.-M.; Son, H.J.; Joh, T.H.; Kim, Y.S.; Kim, D.; Flint Beal, M.; Hwang, O. A Novel Intracellular Role of Matrix Metalloproteinase-3 during Apoptosis of Dopaminergic Cells. J. Neurochem. 2008, 106, 405–415. [Google Scholar] [CrossRef]
- Powell, B.S.; Dhaher, Y.Y.; Szleifer, I.G. Review of the Multiscale Effects of Female Sex Hormones on Matrix Metalloproteinase-Mediated Collagen Degradation. Crit. Rev. Biomed. Eng. 2015, 43, 401–428. [Google Scholar] [CrossRef]
- Fainardi, E.; Castellazzi, M.; Bellini, T.; Manfrinato, M.C.; Baldi, E.; Casetta, I.; Paolino, E.; Granieri, E.; Dallocchio, F. Cerebrospinal Fluid and Serum Levels and Intrathecal Production of Active Matrix Metalloproteinase-9 (MMP-9) as Markers of Disease Activity in Patients with Multiple Sclerosis. Mult. Scler. 2006, 12, 294–301. [Google Scholar] [CrossRef]
- Bjerke, M.; Zetterberg, H.; Edman, Å.; Blennow, K.; Wallin, A.; Andreasson, U. Cerebrospinal Fluid Matrix Metalloproteinases and Tissue Inhibitor of Metalloproteinases in Combination with Subcortical and Cortical Biomarkers in Vascular Dementia and Alzheimer’s Disease. J. Alzheimers Dis. 2011, 27, 665–676. [Google Scholar] [CrossRef]
- Gutierrez, J.; Menshawy, K.; Goldman, J.; Dwork, A.J.; Elkind, M.S.V.; Marshall, R.S.; Morgello, S. Metalloproteinases and Brain Arterial Remodeling Among Individuals with and Those Without HIV Infection. J. Infect. Dis. 2016, 214, 1329–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, J.; Sacco, R.L.; Wright, C.B. Dolichoectasia-an Evolving Arterial Disease. Nat. Rev. Neurol. 2011, 7, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Kiaei, M.; Kipiani, K.; Calingasan, N.Y.; Wille, E.; Chen, J.; Heissig, B.; Rafii, S.; Lorenzl, S.; Beal, M.F. Matrix Metalloproteinase-9 Regulates TNF-Alpha and FasL Expression in Neuronal, Glial Cells and Its Absence Extends Life in a Transgenic Mouse Model of Amyotrophic Lateral Sclerosis. Exp. Neurol. 2007, 205, 74–81. [Google Scholar] [CrossRef]
- Kaplan, A.; Spiller, K.J.; Towne, C.; Kanning, K.C.; Choe, G.T.; Geber, A.; Akay, T.; Aebischer, P.; Henderson, C.E. Neuronal Matrix Metalloproteinase-9 Is a Determinant of Selective Neurodegeneration. Neuron 2014, 81, 333–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.P.; Holcomb, J.; Al-Ramahi, I.; de Haro, M.; Gafni, J.; Zhang, N.; Kim, E.; Sanhueza, M.; Torcassi, C.; Kwak, S.; et al. Matrix Metalloproteinases Are Modifiers of Huntingtin Proteolysis and Toxicity in Huntington’s Disease. Neuron 2010, 67, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, G.A. Extracellular Matrix Inflammation in Vascular Cognitive Impairment and Dementia. Clin. Sci. 2017, 131, 425–437. [Google Scholar] [CrossRef]
- Lech, A.M.; Wiera, G.; Mozrzymas, J.W. Matrix Metalloproteinase-3 in Brain Physiology and Neurodegeneration. Adv. Clin. Exp. Med. 2019, 28, 1717–1722. [Google Scholar] [CrossRef]
- Mikkola, T.S.; Gissler, M.; Merikukka, M.; Tuomikoski, P.; Ylikorkala, O. Sex Differences in Age-Related Cardiovascular Mortality. PLoS ONE 2013, 8, e63347. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, A.; Hertig, V.; Villeneuve, L.; Chauvette, V.; El-Hamamsy, I.; Calderone, A. The Ascending Aorta of Male Hypertensive Bicuspid Aortic Valve Patients Preferentially Associated with a Cellular Aneurysmal Phenotype. Physiol. Rep. 2022, 10, e15251. [Google Scholar] [CrossRef]
- DiMusto, P.D.; Lu, G.; Ghosh, A.; Roelofs, K.J.; Sadiq, O.; McEvoy, B.; Su, G.; Laser, A.; Bhamidipati, C.M.; Ailawadi, G.; et al. Increased JNK in Males Compared with Females in a Rodent Model of Abdominal Aortic Aneurysm. J. Surg. Res. 2012, 176, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Ehrlichman, L.K.; Ford, J.W.; Roelofs, K.J.; Tedeschi-Filho, W.; Futchko, J.S.; Ramacciotti, E.; Eliason, J.L.; Henke, P.K.; Upchurch, G.R. Gender-Dependent Differential Phosphorylation in the ERK Signaling Pathway Is Associated with Increased MMP2 Activity in Rat Aortic Smooth Muscle Cells. J. Surg. Res. 2010, 160, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Sokolis, D.P.; Iliopoulos, D.C. Impaired Mechanics and Matrix Metalloproteinases/Inhibitors Expression in Female Ascending Thoracic Aortic Aneurysms. J. Mech. Behav. Biomed. Mater. 2014, 34, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shen, M.; Parajuli, N.; Oudit, G.Y.; McMurtry, M.S.; Kassiri, Z. Gender-Dependent Aortic Remodelling in Patients with Bicuspid Aortic Valve-Associated Thoracic Aortic Aneurysm. J. Mol. Med. (Berl.) 2014, 92, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.W.; Maslen, C.L.; Kindem, M.; Kroner, B.L.; Song, H.K.; Ravekes, W.; Dietz, H.C.; Weinsaft, J.W.; Roman, M.J.; Devereux, R.B.; et al. GenTAC Registry Report: Gender Differences among Individuals with Genetically Triggered Thoracic Aortic Aneurysm and Dissection. Am. J. Med. Genet. 2013, 161, 779–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, E.M.; Pearce, S.W.A.; Xiao, R.; Oo, A.Y.; Xiao, Q. Matrix Metalloproteinase in Abdominal Aortic Aneurysm and Aortic Dissection. Pharmaceuticals 2019, 12, 118. [Google Scholar] [CrossRef] [Green Version]
- Cavasin, M.A.; Tao, Z.; Menon, S.; Yang, X.-P. Gender Differences in Cardiac Function during Early Remodeling after Acute Myocardial Infarction in Mice. Life Sci. 2004, 75, 2181–2192. [Google Scholar] [CrossRef]
- Münch, J.; Avanesov, M.; Bannas, P.; Säring, D.; Krämer, E.; Mearini, G.; Carrier, L.; Suling, A.; Lund, G.; Patten, M. Serum Matrix Metalloproteinases as Quantitative Biomarkers for Myocardial Fibrosis and Sudden Cardiac Death Risk Stratification in Patients With Hypertrophic Cardiomyopathy. J. Card. Fail. 2016, 22, 845–850. [Google Scholar] [CrossRef]
- Samnegård, A.; Silveira, A.; Tornvall, P.; Hamsten, A.; Ericsson, C.-G.; Eriksson, P. Lower Serum Concentration of Matrix Metalloproteinase-3 in the Acute Stage of Myocardial Infarction. J. Intern. Med. 2006, 259, 530–536. [Google Scholar] [CrossRef]
- Samnegård, A.; Silveira, A.; Lundman, P.; Boquist, S.; Odeberg, J.; Hulthe, J.; McPheat, W.; Tornvall, P.; Bergstrand, L.; Ericsson, C.-G.; et al. Serum Matrix Metalloproteinase-3 Concentration Is Influenced by MMP-3 -1612 5A/6A Promoter Genotype and Associated with Myocardial Infarction. J. Intern. Med. 2005, 258, 411–419. [Google Scholar] [CrossRef]
- Samnegård, A.; Hulthe, J.; Silveira, A.; Ericsson, C.-G.; Hamsten, A.; Eriksson, P. Gender Specific Associations between Matrix Metalloproteinases and Inflammatory Markers in Post Myocardial Infarction Patients. Atherosclerosis 2009, 202, 550–556. [Google Scholar] [CrossRef]
- Hellings, W.E.; Pasterkamp, G.; Verhoeven, B.A.N.; de Kleijn, D.P.V.; de Vries, J.-P.P.M.; Seldenrijk, K.A.; van den Broek, T.; Moll, F.L. Gender-Associated Differences in Plaque Phenotype of Patients Undergoing Carotid Endarterectomy. J. Vasc. Surg. 2007, 45, 289–296, discussion 296-7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djurić, T.; Zivković, M.; Stanković, A.; Kolaković, A.; Jekić, D.; Selaković, V.; Alavantić, D. Plasma Levels of Matrix Metalloproteinase-8 in Patients with Carotid Atherosclerosis. J. Clin. Lab. Anal. 2010, 24, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, L.D.S.; Bown, M.J.; Tomaszewski, M. Sexual Dimorphism of Abdominal Aortic Aneurysms: A Striking Example of “Male Disadvantage” in Cardiovascular Disease. Atherosclerosis 2012, 225, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix Metalloproteinase-9: Many Shades of Function in Cardiovascular Disease. Physiology 2013, 28, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ailawadi, G.; Eliason, J.L.; Roelofs, K.J.; Sinha, I.; Hannawa, K.K.; Kaldjian, E.P.; Lu, G.; Henke, P.K.; Stanley, J.C.; Weiss, S.J.; et al. Gender Differences in Experimental Aortic Aneurysm Formation. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2116–2122. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Keimig, T.; He, Q.; Ding, J.; Zhang, Z.; Pourabdollah-Nejad, S.; Yang, X.-P. Augmented Healing Process in Female Mice with Acute Myocardial Infarction. Gend. Med. 2007, 4, 230–247. [Google Scholar] [CrossRef]
- Fang, L.; Gao, X.-M.; Moore, X.-L.; Kiriazis, H.; Su, Y.; Ming, Z.; Lim, Y.L.; Dart, A.M.; Du, X.-J. Differences in Inflammation, MMP Activation and Collagen Damage Account for Gender Difference in Murine Cardiac Rupture Following Myocardial Infarction. J. Mol. Cell Cardiol. 2007, 43, 535–544. [Google Scholar] [CrossRef]
- Garvin, P.; Jonasson, L.; Nilsson, L.; Falk, M.; Kristenson, M. Plasma Matrix Metalloproteinase-9 Levels Predict First-Time Coronary Heart Disease: An 8-Year Follow-Up of a Community-Based Middle Aged Population. PLoS ONE 2015, 10, e0138290. [Google Scholar] [CrossRef]
- Kobayashi, N.; Hata, N.; Kume, N.; Yokoyama, S.; Shinada, T.; Tomita, K.; Kitamura, M.; Shirakabe, A.; Inami, T.; Yamamoto, M.; et al. Matrix Metalloproteinase-9 for the Earliest Stage Acute Coronary Syndrome. Circ. J. 2011, 75, 2853–2861. [Google Scholar] [CrossRef] [Green Version]
- Tayebjee, M.H.; Lip, G.Y.H.; Tan, K.T.; Patel, J.V.; Hughes, E.A.; MacFadyen, R.J. Plasma Matrix Metalloproteinase-9, Tissue Inhibitor of Metalloproteinase-2, and CD40 Ligand Levels in Patients with Stable Coronary Artery Disease. Am. J. Cardiol. 2005, 96, 339–345. [Google Scholar] [CrossRef]
- Sullivan, S.; Hammadah, M.; Wilmot, K.; Ramadan, R.; Pearce, B.D.; Shah, A.; Kaseer, B.; Gafeer, M.M.; Lima, B.B.; Kim, J.H.; et al. Young Women With Coronary Artery Disease Exhibit Higher Concentrations of Interleukin-6 at Baseline and in Response to Mental Stress. J. Am. Heart Assoc. 2018, 7, e010329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, B.S.; Roelofs, K.J.; Ford, J.W.; Henke, P.K.; Upchurch, G.R. Decreased Collagen and Increased Matrix Metalloproteinase-13 in Experimental Abdominal Aortic Aneurysms in Males Compared with Females. Surgery 2010, 147, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronado, M.J.; Brandt, J.E.; Kim, E.; Bucek, A.; Bedja, D.; Abston, E.D.; Shin, J.; Gabrielson, K.L.; Mitzner, W.; Fairweather, D. Testosterone and Interleukin-1β Increase Cardiac Remodeling during Coxsackievirus B3 Myocarditis via Serpin A 3n. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1726–H1736. [Google Scholar] [CrossRef] [PubMed]
- Passos, L.S.A.; Becker-Greene, D.; Braulio, R.; Le, T.-D.; Gelape, C.L.; de Almeida, L.F.R.; Rocha, D.P.A.; Gomes, C.A.P.; Esteves, W.A.M.; Passaglia, L.G.; et al. Proinflammatory Matrix Metalloproteinase-1 Associates with Mitral Valve Leaflet Disruption Following Percutaneous Mitral Valvuloplasty. Front. Cardiovasc. Med. 2021, 8, 804111. [Google Scholar] [CrossRef]
- Banerjee, T.; Mukherjee, S.; Ghosh, S.; Biswas, M.; Dutta, S.; Pattari, S.; Chatterjee, S.; Bandyopadhyay, A. Clinical Significance of Markers of Collagen Metabolism in Rheumatic Mitral Valve Disease. PLoS ONE 2014, 9, e90527. [Google Scholar] [CrossRef]
- Elhewala, A.A.; Sanad, M.; Soliman, A.M.; Sami, M.M.; Ahmed, A.A. Matrix Metalloproteinase-9 in Pediatric Rheumatic Heart Disease with and without Heart Failure. Biomed. Rep. 2021, 14, 4. [Google Scholar] [CrossRef]
- Lee, S.-D.; Chen, L.-M.; Kuo, W.-W.; Shu, W.-T.; Kuo, W.-H.; Huang, E.-J.; Tsai, C.-C.; Li, P.-C.; Liu, J.-Y.; Chen, T.-H.; et al. Serum Insulin-like Growth Factor-Axis and Matrix Metalloproteinases in Patients with Rheumatic Arthritis or Rheumatic Heart Disease. Clin. Chim. Acta. 2006, 367, 62–68. [Google Scholar] [CrossRef]
- Giannakos, E.; Vardali, E.; Bartekova, M.; Fogarassyova, M.; Barancik, M.; Radosinska, J. Changes in Activities of Circulating MMP-2 and MMP-9 in Patients Suffering from Heart Failure in Relation to Gender, Hypertension and Treatment: A Cross-Sectional Study. Physiol. Res. 2016, 65, S149. [Google Scholar] [CrossRef]
- Mendes, P.R.; Kiyota, T.A.; Cipolli, J.A.; Schreiber, R.; Paim, L.R.; Bellinazzi, V.R.; Matos-Souza, J.R.; Sposito, A.C.; Nadruz, W. Gender Influences the Relationship between Lung Function and Cardiac Remodeling in Hypertensive Subjects. Hypertens. Res. 2015, 38, 264–268. [Google Scholar] [CrossRef]
- Ramani, R.; Mathier, M.; Wang, P.; Gibson, G.; Tögel, S.; Dawson, J.; Bauer, A.; Alber, S.; Watkins, S.C.; McTiernan, C.F.; et al. Inhibition of Tumor Necrosis Factor Receptor-1-Mediated Pathways Has Beneficial Effects in a Murine Model of Postischemic Remodeling. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1369–H1377. [Google Scholar] [CrossRef]
- Gu, C.; Wang, F.; Hou, Z.; Lv, B.; Wang, Y.; Cong, X.; Chen, X. Sex-Related Differences in Serum Matrix Metalloproteinase-9 Screening Non-Calcified and Mixed Coronary Atherosclerotic Plaques in Outpatients with Chest Pain. Heart Vessel. 2017, 32, 1424–1431. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.M. Normal Sexual Dimorphism of the Adult Human Brain Assessed by In Vivo Magnetic Resonance Imaging. Cereb. Cortex 2001, 11, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, S.; Benkelfat, C.; Young, S.N.; Leyton, M.; Mzengeza, S.; de Montigny, C.; Blier, P.; Diksic, M. Differences between Males and Females in Rates of Serotonin Synthesis in Human Brain. Proc. Natl. Acad. Sci. USA 1997, 94, 5308–5313. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Cao, J.; Yu, Y.; Yang, H.; Jiang, Y.; Liu, Y.; Wang, R.; Zhao, Q. Gender-Related Differences in Regional Cerebral Glucose Metabolism in Normal Aging Brain. Front. Aging Neurosci. 2022, 14, 809767. [Google Scholar] [CrossRef] [PubMed]
- Gur, R.C.; Gur, R.E. Complementarity of Sex Differences in Brain and Behavior: From Laterality to Multimodal Neuroimaging. J. Neurosci. Res. 2017, 95, 189–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Lancet Neurology A Spotlight on Sex Differences in Neurological Disorders. Lancet Neurol. 2019, 18, 319. [CrossRef]
- Wooten, G.F. Are Men at Greater Risk for Parkinson’s Disease than Women? J. Neurol. Neurosurg. Psychiatry 2004, 75, 637–639. [Google Scholar] [CrossRef]
- McHugh, J.C.; Delanty, N. Epidemiology and Classification of Epilepsy: Gender Comparisons. Int. Rev. Neurobiol. 2008, 83, 11–26. [Google Scholar] [CrossRef]
- Vyas, M.V.; Silver, F.L.; Austin, P.C.; Yu, A.Y.X.; Pequeno, P.; Fang, J.; Laupacis, A.; Kapral, M.K. Stroke Incidence by Sex Across the Lifespan. Stroke 2021, 52, 447–451. [Google Scholar] [CrossRef]
- Manjaly, Z.R.; Scott, K.M.; Abhinav, K.; Wijesekera, L.; Ganesalingam, J.; Goldstein, L.H.; Janssen, A.; Dougherty, A.; Willey, E.; Stanton, B.R.; et al. The Sex Ratio in Amyotrophic Lateral Sclerosis: A Population Based Study. Amyotroph. Lateral Scler. 2010, 11, 439–442. [Google Scholar] [CrossRef]
- Na, W.; Lee, J.Y.; Kim, W.-S.; Yune, T.Y.; Ju, B.-G. 17β-Estradiol Ameliorates Tight Junction Disruption via Repression of MMP Transcription. Mol. Endocrinol. 2015, 29, 1347–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellazzi, M.; Pizzicotti, S.; Lombardo, I.; Alfiero, S.; Morotti, A.; Pellegatti, P.; Negri, G.; Natali, L.; Ferri, C.; Fainardi, E.; et al. Sexual Dimorphism in the Cerebrospinal Fluid Total Protein Content. Clin. Chem. Lab. Med. 2020, 58, 1885–1890. [Google Scholar] [CrossRef] [PubMed]
- McCudden, C.R.; Brooks, J.; Figurado, P.; Bourque, P.R. Cerebrospinal Fluid Total Protein Reference Intervals Derived from 20 Years of Patient Data. Clin. Chem. 2017, 63, 1856–1865. [Google Scholar] [CrossRef] [PubMed]
- Castellazzi, M.; Morotti, A.; Tamborino, C.; Alessi, F.; Pilotto, S.; Baldi, E.; Caniatti, L.M.; Trentini, A.; Casetta, I.; Granieri, E.; et al. Increased Age and Male Sex Are Independently Associated with Higher Frequency of Blood-Cerebrospinal Fluid Barrier Dysfunction Using the Albumin Quotient. Fluids Barriers CNS 2020, 17, 14. [Google Scholar] [CrossRef]
- Meixensberger, S.; Bechter, K.; Dersch, R.; Feige, B.; Maier, S.; Schiele, M.A.; Runge, K.; Denzel, D.; Nickel, K.; Spieler, D.; et al. Sex Difference in Cerebrospinal Fluid/Blood Albumin Quotients in Patients with Schizophreniform and Affective Psychosis. Fluids Barriers CNS 2020, 17, 67. [Google Scholar] [CrossRef]
- Parrado-Fernández, C.; Blennow, K.; Hansson, M.; Leoni, V.; Cedazo-Minguez, A.; Björkhem, I. Evidence for Sex Difference in the CSF/Plasma Albumin Ratio in ~20 000 Patients and 335 Healthy Volunteers. J. Cell. Mol. Med. 2018, 22, 5151–5154. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.; Lim, C.; Kim, Y.; Moon, W.-J. Sex-Related Differences in Regional Blood-Brain Barrier Integrity in Non-Demented Elderly Subjects. Int. J. Mol. Sci. 2021, 22, 2860. [Google Scholar] [CrossRef]
- Gupta, V.; Singh, M.K.; Garg, R.K.; Pant, K.K.; Khattri, S. Evaluation of Peripheral Matrix Metalloproteinase-1 in Parkinson’s Disease: A Case-Control Study. Int. J. Neurosci. 2014, 124, 88–92. [Google Scholar] [CrossRef]
- Castellazzi, M.; Ligi, D.; Contaldi, E.; Quartana, D.; Fonderico, M.; Borgatti, L.; Bellini, T.; Trentini, A.; Granieri, E.; Fainardi, E.; et al. Multiplex Matrix Metalloproteinases Analysis in the Cerebrospinal Fluid Reveals Potential Specific Patterns in Multiple Sclerosis Patients. Front. Neurol. 2018, 9, 1080. [Google Scholar] [CrossRef]
- Iulita, M.F.; Ganesh, A.; Pentz, R.; Flores Aguilar, L.; Gubert, P.; Ducatenzeiler, A.; Christie, S.; Wilcock, G.K.; Cuello, A.C. Identification and Preliminary Validation of a Plasma Profile Associated with Cognitive Decline in Dementia and At-Risk Individuals: A Retrospective Cohort Analysis. J. Alzheimers Dis. 2019, 67, 327–341. [Google Scholar] [CrossRef]
- Pentz, R.; Iulita, M.F.; Mikutra-Cencora, M.; Ducatenzeiler, A.; Bennett, D.A.; Cuello, A.C. A New Role for Matrix Metalloproteinase-3 in the NGF Metabolic Pathway: Proteolysis of Mature NGF and Sex-Specific Differences in the Continuum of Alzheimer’s Pathology. Neurobiol. Dis. 2021, 148, 105150. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zeng, G.Q.; Liu, X.; Tong, R.Z.; Zhou, D.; Hong, Z. Evaluation of Serum Matrix Metalloproteinase-3 as a Biomarker for Diagnosis of Epilepsy. J. Neurol. Sci. 2016, 367, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Yang, J.; Xu, T.; Xu, T.; Peng, Y.; Wang, A.; Wang, J.; Peng, H.; Li, Q.; Ju, Z.; et al. Serum Matrix Metalloproteinase-9 Levels and Prognosis of Acute Ischemic Stroke. Neurology 2017, 89, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Howe, M.D.; Furr, J.W.; Zhu, L.; Edwards, N.J.; McCullough, L.D.; Gonzales, N.R. Sex-Specific Association of Matrix Metalloproteinases with Secondary Injury and Outcomes after Intracerebral Hemorrhage. J. Stroke Cerebrovasc. Dis. 2019, 28, 1718–1725. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MMPs | CVDs | Role | References |
|---|---|---|---|
| MMP-2 | Hypertension | Causes vasoconstriction | [41,42,43] |
| Limits vasodilation | [44,45] | ||
| Cardiomyopathies | Increases infarct areas | [46] | |
| Process several cardiac proteins | [47,48] | ||
| MMP-1, -2, -3, -8, and -9 | Aneurism | Weakens the ECM structure | [49,50] |
| MMP-2 and -9 | Myocardial infarction | Degrade ECM/process cytokines and chemokines | [51] |
| MMP-2, -3, -8, -9, and 12 | atherosclerosis | connected to plaque development | [52,53,54,55,56] |
| MMP-7 and -9 | hypertension | cause vasoconstriction/increase blood pressure | [43] |
| MMPs | CNS Conditions | Role | References |
|---|---|---|---|
| MMP-2 | AD | Protective | [76] |
| MS | Associated to remission | [77] | |
| MMP-3 | AD and PD | Detrimental | [78,79] |
| MMP-9 | MS | Associated to demyelination | [80] |
| AD | Associated to neuronal cell death | [81] | |
| PD and dolichoectasia | Associated to neuroinflammation | [20,82,83] | |
| ALS | Associated to neuroinflammation | [84,85] | |
| MMP-10 | HD | Detrimental | [86] |
| MMP-2, -3, -9 | HI lesions | Increase BBB permeability | [74] |
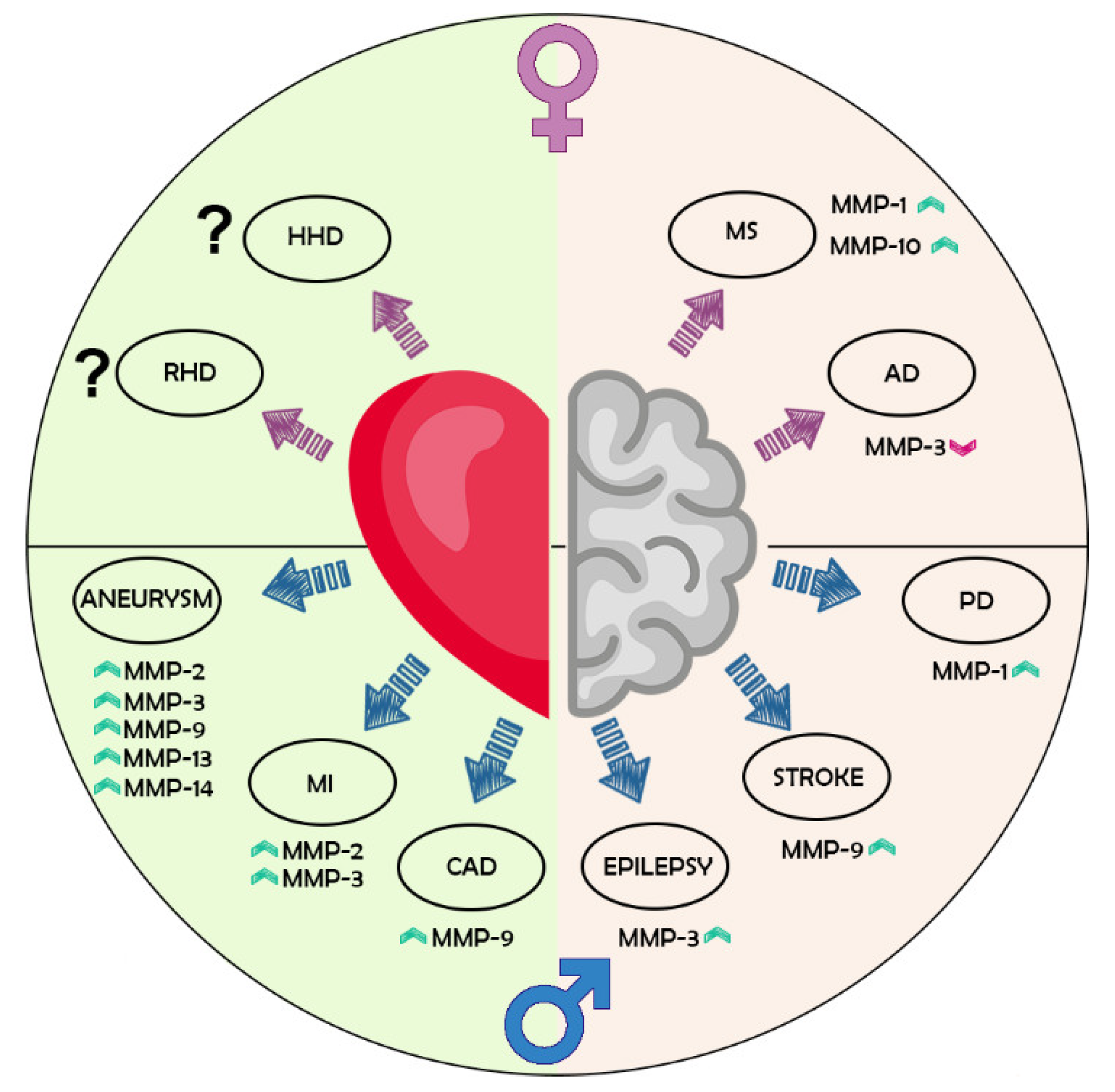
| MMPs | CVD | Organism | Male | Female | References |
|---|---|---|---|---|---|
| MMP-2 | Aneurism | Human/Mouse | ↑ in aorta | [90,91] | |
| Thoracic aortic aneurysm | Human | ↑ in aorta | [93,94] | ||
| Acute MI | Mouse | ↑ in affected heart | [97] | ||
| Heart failure | Human | ↓ in serum | [119] | ||
| Hypertension | Human | No change | No change | [120] | |
| Myocardial fibrosis | Human | ↑ serum concentration associated with LOWER fibrosis | [98] | ||
| MMP-3 | MI | Human | ↓ in serum; positively correlated with inflammatory markers | [99,100,101] | |
| MMP-8 | Thoracic aortic aneurysm | Human | ↓ in aorta | [94] | |
| Carotid atherosclerosis | Human | No difference | No difference; ↓ in plaques. The difference disappeared in multivariate correction | [102,103] | |
| MMP-9 | Aneurism/Ascending thoracic aortic aneurysms | Human/Mouse | ↑ in the ascending aorta | ↑ in aorta | [90,91,93,106] |
| MI | Mouse/Rat | ↑ in heart | ↓ in hearth | [107,97] | |
| post MI | Human/Mouse | No change/Positively correlated with inflammatory markers; ↑ in Heart | No change | [121,101] | |
| Cardiac rupture following MI | Mouse | ↑ in left ventricle | [108] | ||
| Myocardial fibrosis | Human | ↑ serum concentration associated with HIGHER fibrosis | [98] | ||
| CAD/CHD | Human | ↑ in serum | ↓ in serum | [109,110,111,112] | |
| Chest Pain | Human | ↑ in serum (patients with non-calcified and mixed plaques) | [122] | ||
| Hypertension | Human | No change | No change | [120] | |
| MMP-13 | Thoracic aortic aneurysm | Human/Rat | ↓ in aorta | [94,113] | |
| MMP-14 | Thoracic aortic aneurysm | Human | ↑ in aorta | [94] |
| MMPs | CNS Conditions | Male | Female | References |
|---|---|---|---|---|
| MMP-1 | PD | ↓ in serum | [139] | |
| MS and neurological controls | ↑ CSF levels | [140] | ||
| MMP-3 | AD, MCI, and cognitively normal individuals | ↑ frontal cortex protein levels | [142] | |
| ↑ plasma levels | ↑ levels associated to greater cognitive decline | [141] | ||
| Epilepsy | ↑ serum levels | [143] | ||
| ICH | associated to long-term functional outcomes | [145] | ||
| MMP-9 | Ischemic stroke | ↑ serum levels in acute phase | [144] | |
| MMP-10 | ICH | Associated to long-term functional outcomes | [145] | |
| MS | ↑ CSF levels | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trentini, A.; Manfrinato, M.C.; Castellazzi, M.; Bellini, T. Sex-Related Differences of Matrix Metalloproteinases (MMPs): New Perspectives for These Biomarkers in Cardiovascular and Neurological Diseases. J. Pers. Med. 2022, 12, 1196. https://doi.org/10.3390/jpm12081196
Trentini A, Manfrinato MC, Castellazzi M, Bellini T. Sex-Related Differences of Matrix Metalloproteinases (MMPs): New Perspectives for These Biomarkers in Cardiovascular and Neurological Diseases. Journal of Personalized Medicine. 2022; 12(8):1196. https://doi.org/10.3390/jpm12081196
Chicago/Turabian StyleTrentini, Alessandro, Maria Cristina Manfrinato, Massimiliano Castellazzi, and Tiziana Bellini. 2022. "Sex-Related Differences of Matrix Metalloproteinases (MMPs): New Perspectives for These Biomarkers in Cardiovascular and Neurological Diseases" Journal of Personalized Medicine 12, no. 8: 1196. https://doi.org/10.3390/jpm12081196
APA StyleTrentini, A., Manfrinato, M. C., Castellazzi, M., & Bellini, T. (2022). Sex-Related Differences of Matrix Metalloproteinases (MMPs): New Perspectives for These Biomarkers in Cardiovascular and Neurological Diseases. Journal of Personalized Medicine, 12(8), 1196. https://doi.org/10.3390/jpm12081196