Biological Adaptations Associated with Dehydration in Mosquitoes


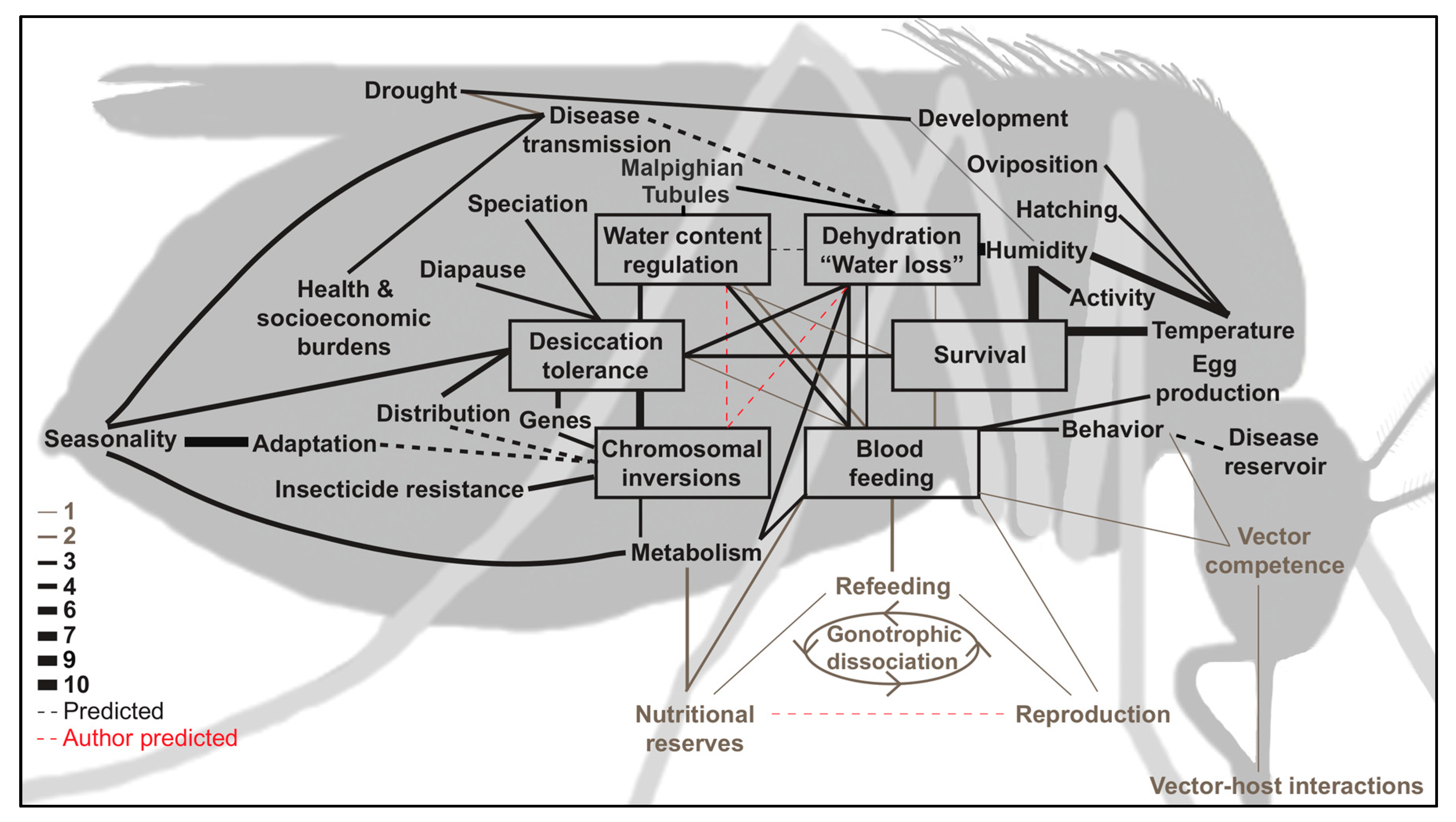{kind=link}
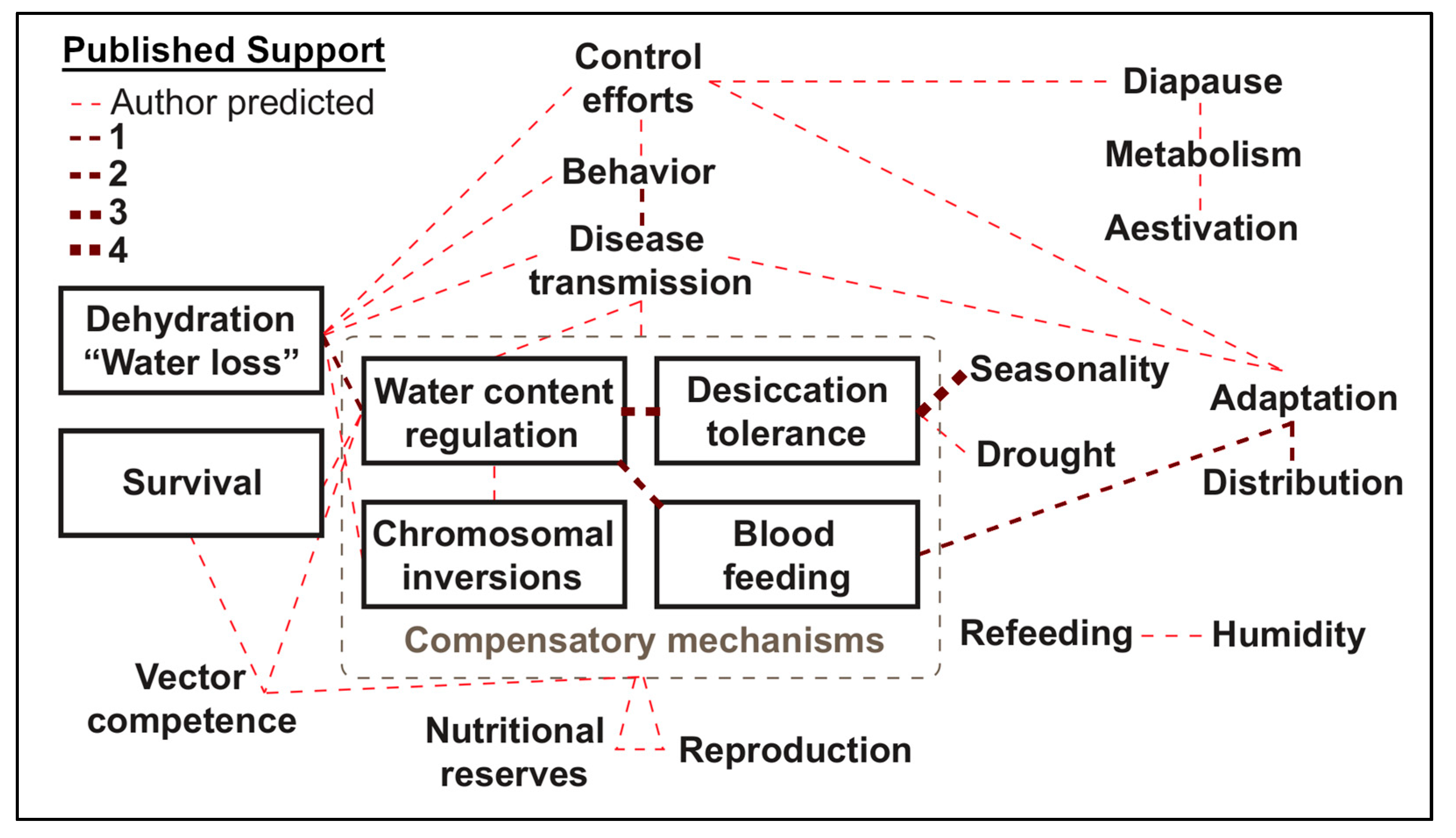{kind=link}
Abstract
:1. Basics of Desiccation Resistance in Mosquitoes
1.1. Water Homeostasis Is Paramount for Mosquito Survival
1.2. Current (Mis)Understandings of Mosquito Dehydration and Resulting Consequences in the Field
1.3. Behavioral Plasticity Is Relevant and Important in Response to Mosquito Dehydration Stress
1.4. A Plethora of Factors Influence Mosquito Biology and Interactions between Stressors Should Be Scrutinized
2. Adaptations to and Influences of Mosquito Dehydration Dynamics
2.1. Wet and Dry Season Dynamics
2.2. Long-term Adaptations for Survival During Unfavorable Periods
2.2.1. Diapause, Migration, Aestivation, and Likely Anthropophilia, Promote Survival and May Establish Disease Reservoirs During Adverse Seasons
2.2.2. Examination of the Genetic Components in Desiccation Resistance Offers a Glimpse into the Mechanistic Underpinnings
2.2.3. Chromosomal Inversion Polymorphisms Underlie Climatic Adaptations
Climatic Adaptation and Distribution
Gene Regulation and Metabolism Related to Chromosomal Inversions
2.3. Adaptations to Dehydration Confer Biological Advantages
2.3.1. Spiracule and Cuticular Hydrocarbon Adaptations Impact Adult Desiccation Resistance
2.3.2. Metabolic and Transcriptomic Alterations Caused by Dehydration Affect Processes Integral to Mosquito Biology
2.3.3. Nutritional Reserve Dynamics Underlie Many Behavioral Adaptations
2.3.4. Reproductive Plasticity is Inherently Related to Maternal Condition and is Likely Influenced by Environmental Conditions
2.3.5. Desiccation Tolerance in Mosquito Eggs Fosters Adaptation to Dry Environments
3. Interactions Between Dehydrating Conditions and Vectorial Capacity
3.1. Contributions of Behavior to Vectorial Capacity
3.1.1. Blood Feeding and Water Content Regulation Influence Disease Transmission Dynamics
3.1.2. Mosquito Humidity Sensing is Influenced by Hydration Status and Likely Contributes to altered Pathogen Transmission
3.2. Alternative Perspectives: Utilizing Environmental Stressors as a Proxy for Behavioral and Biological Adaptation to Insecticide Resistance
3.3. Incorporating the Underlying Mechanisms of Desiccation Dynamics into Disease Transmission Models Would Promote Accuracy and Efficacy
4. Caveats and Future Directions in Desiccation-disease Transmission Research
4.1. Utilizing Vapor Pressure Deficit and Increasing Data Transparency Would Promote Interstudy Interpretability
4.2. Desiccation Resistance Dynamics are Likely Applicable in Other Arthropod Systems of Interest and May Be Utilized to Foster Comparative Interspecies Analyses
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Caraballo, H.; King, K. Emergency Department Management Of Mosquito- Borne Illness: Malaria, Dengue, And West Nile Virus. Emerg. Med. Pract. 2014, 16, 1–24. [Google Scholar] [PubMed]
- Torres, J.R.; Castro, J. The Health and Economic Impact of Dengue in Latin America. Cad. Saude Publica 2007, 23, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Dash, A.P.; Bhatia, R.; Sunyoto, T.; Mourya, D.T. Emerging and Re-Emerging Arboviral Diseases in Southeast Asia. J. Vector Borne Dis. 2013, 50, 77–84. [Google Scholar] [PubMed]
- Gubler, D.J. Resurgent Vector-Borne Diseases as a Global Health Problem. Emerg. Infect. Dis. 1998, 4, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Epidemic Dengue/Dengue Hemorrhagic Fever as a Public Health, Social and Economic Problem in the 21st Century. Trends Microbiol. 2002, 10, 100–103. [Google Scholar] [CrossRef]
- Hagan, R.W.; Didion, E.M.; Rosselot, A.E.; Holmes, C.J.; Siler, S.C.; Rosendale, A.J.; Hendershot, J.M.; Elliot, K.S.B.; Jennings, E.C.; Nine, G.A.; et al. Dehydration Prompts Increased Activity and Blood Feeding by Mosquitoes. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Canyon, D.V.; Hii, J.L.K.; Müller, R. Adaptation of Aedes aegypti (Diptera: Culicidae) Oviposition Behavior in Response to Humidity and Diet. J. Insect Physiol. 1999, 45, 959–964. [Google Scholar] [CrossRef]
- Benoit, J.B.; Patrick, K.R.; Desai, K.; Hardesty, J.J.; Krause, T.B.; Denlinger, D.L. Repeated Bouts of Dehydration Deplete Nutrient Reserves and Reduce Egg Production in the Mosquito Culex pipiens. J. Exp. Biol. 2010, 213, 2763–2769. [Google Scholar] [CrossRef]
- Reidenbach, K.R.; Cheng, C.; Liu, F.; Liu, C.; Besansky, N.J.; Syed, Z. Cuticular Differences Associated with Aridity Acclimation in African Malaria Vectors Carrying Alternative Arrangements of Inversion 2La. Parasit. Vectors 2014, 7, 1–13. [Google Scholar] [CrossRef]
- Piermarini, P.M. Renal Excretory Processes in Mosquitoes, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; Volume 51. [Google Scholar]
- Benoit, J.B.; Denlinger, D.L. Suppression of Water Loss during Adult Diapause in the Northern House Mosquito, Culex pipiens. J. Exp. Biol. 2007, 210, 217–226. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Robich, R.M.; Denlinger, D.L. Enhanced Cold and Desiccation Tolerance in Diapausing Adults of Culex pipiens, and a Role for Hsp70 in Response to Cold Shock but Not as a Component of the Diapause Program. J. Med. Entomol. 2006, 43, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Denlinger, D.L.; Piermarini, P.M. The Diapause Program Impacts Renal Excretion and Molecular Expression of Aquaporins in the Northern House Mosquito, Culex pipiens. J. Insect Physiol. 2017, 98, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Dao, A.; Yaro, A.S.; Diallo, M.; Timbine, S.; Huestis, D.L.; Kassogue, Y.; Traore, A.I.; Sanogo, Z.L.; Samake, D.; Lehmann, T. Signatures of Aestivation and Migration in Sahelian Malaria Mosquito Populations. Nature 2014, 516, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Mamai, W.; Simard, F.; Couret, D.; Ouedraogo, G.A.; Renault, D.; Dabiré, K.R.; Mouline, K. Monitoring Dry Season Persistence of Anopheles gambiae s.l. Populations in a Contained Semi-Field System in Southwestern Burkina Faso, West Africa. J. Med. Entomol. 2015, 53, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Benoit, J.B.; Denlinger, D.L. Meeting the Challenges of On-Host and Off-Host Water Balance in Blood-Feeding Arthropods. J. Insect Physiol. 2010, 56, 1366–1376. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, B.J.; Gibbs, A.G.; Roberts, S.P. Gene Transcription During Exposure to, and Recovery from, Cold and Desiccation Stress in Drosophila melanogaster. Insect Mol. Biol. 2007, 16, 435–443. [Google Scholar] [CrossRef]
- Cheng, C.; Tan, J.C.; Hahn, M.W.; Besansky, N.J. Systems Genetic Analysis of Inversion Polymorphisms in the Malaria Mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2018, 115, 7005–7014. [Google Scholar] [CrossRef]
- Drake, L.L.; Boudko, D.Y.; Marinotti, O.; Carpenter, V.K.; Dawe, A.L.; Hansen, I.A. The Aquaporin Gene Family of the Yellow Fever Mosquito, Aedes aegypti. PLoS ONE 2010, 5, e15578. [Google Scholar] [CrossRef]
- Vaidyanathan, R.; Fleisher, A.E.; Minnick, S.L.; Simmons, K.A.; Scott, T.W. Nutritional Stress Affects Mosquito Survival and Vector Competence for West Nile Virus. Vector Borne Zoonotic Dis. 2008, 8, 727–732. [Google Scholar] [CrossRef]
- Gates, B. The Deadliest Animal in the World|Bill Gates. Blog Bill Gates 2014, 1. [Google Scholar]
- Ling, L.; Kokoza, V.A.; Zhang, C.; Aksoy, E.; Raikhel, A.S. MicroRNA-277 Targets Insulin-like Peptides 7 and 8 to Control Lipid Metabolism and Reproduction in Aedes aegypti Mosquitoes. Proc. Natl. Acad. Sci. USA 2017, 114, 8017–8024. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, K.; Mouline, K.; Mamai, W.; Foucreau, N.; Dabiré, K.R.; Bouchereau, A.; Simard, F.; Renault, D. Combining Two-Dimensional Gel Electrophoresis and Metabolomic Data in Support of Dry-Season Survival in the Two Main Species of the Malarial Mosquito Anopheles gambiae. Data Br. 2015, 5, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.M.; Ehrlich, H.; Bransfield, A.; Warren, J.L.; Pitzer, V.E.; Brackney, D.E. Successive Bloodmeals Enhance Virus Dissemination Within. bioRxiv 2018, 1–26. [Google Scholar]
- Kilpatrick, A.M.; Meola, M.A.; Moudy, R.M.; Kramer, L.D. Temperature, Viral Genetics, and the Transmission of West Nile Virus by Culex pipiens Mosquitoes. PLoS Pathog. 2008, 4, e1000092. [Google Scholar] [CrossRef]
- Ogden, N.H. Climate Change and Vector-Borne Diseases of Public Health Significance. FEMS Microbiol. Lett. 2017, 364, 1–8. [Google Scholar] [CrossRef]
- Hawley, D.M.; Altizer, S.M. Disease Ecology Meets Ecological Immunology: Understanding the Links between Organismal Immunity and Infection Dynamics in Natural Populations. Funct. Ecol. 2011, 25, 48–60. [Google Scholar] [CrossRef]
- Ragan, I.K.; Blizzard, E.L.; Gordy, P.; Bowen, R.A. Investigating the Potential Role of North American Animals as Hosts for Zika Virus. Vector Borne Zoonotic Dis. 2017, 17, 161–164. [Google Scholar] [CrossRef]
- Carissimo, G.; Pondeville, E.; McFarlane, M.; Dietrich, I.; Mitri, C.; Bischoff, E.; Antoniewski, C.; Bourgouin, C.; Failloux, A.-B.; Kohl, A.; et al. Antiviral Immunity of Anopheles gambiae Is Highly Compartmentalized, with Distinct Roles for RNA Interference and Gut Microbiota. Proc. Natl. Acad. Sci. USA 2015, 112, 176–185. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 3741–3767. [Google Scholar] [CrossRef]
- Hillyer, J.F. Mosquito Immunity. Adv. Exp. Med. Biol. 2010, 708, 218–238. [Google Scholar]
- Liu, K.; Dong, Y.; Huang, Y.; Rasgon, J.L.; Agre, P. Impact of Trehalose Transporter Knockdown on Anopheles gambiae Stress Adaptation and Susceptibility to Plasmodium falciparum Infection. Proc. Natl. Acad. Sci. USA 2013, 110, 17504–17509. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Kim, C.H.; Alto, B.W.; Berenbaum, M.R.; Schuler, M.A. Larval Environmental Stress Alters Aedes aegypti Competence for Sindbis Virus. Trop. Med. Int. Heal. 2011, 16, 955–964. [Google Scholar] [CrossRef]
- Marcantonio, M.; Rizzoli, A.; Metz, M.; Rosa, R.; Marini, G.; Chadwick, E.; Neteler, M. Identifying the Environmental Conditions Favouring West Nile Virus Outbreaks in Europe. PLoS ONE 2015, 10, e0121158. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Reiter, P.; Ebi, K.L.; Yap, W.; Nasci, R.; Patz, J.A. Climate Variability and Change in the United States: Potential Impacts on Vector- and Rodent-Borne Diseases. Environ. Health Perspect. 2001, 109, 223–233. [Google Scholar] [PubMed]
- Canyon, D.V.; Muller, R.; Hii, J.L.K. Aedes aegypti Disregard Humidity-Related Conditions with Adequate Nutrition. Trop. Biomed. 2013, 30, 1–8. [Google Scholar]
- Schmidt, C.A.; Comeau, G.; Monaghan, A.J.; Williamson, D.J.; Ernst, K.C. Effects of Desiccation Stress on Adult Female Longevity in Aedes aegypti and Ae. albopictus (Diptera: Culicidae): Results of a Systematic Review and Pooled Survival Analysis. Parasit. Vectors 2018, 11, 267. [Google Scholar] [CrossRef]
- Sternberg, E.D.; Thomas, M.B. Local Adaptation to Temperature and the Implications for Vector-Borne Diseases. Trends Parasitol. 2014, 30, 115–122. [Google Scholar] [CrossRef]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-Dependent Effects on the Replication and Transmission of Arthropod-Borne Viruses in Their Insect Hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef]
- Yaro, A.S.; Huestis, D.L.; Adamou, A.; Diallo, M.; Dao, A.; Lehmann, T.; Traoré, A.I.; Huestis, D.L.; Adamou, A.; Timbiné, S.; et al. Dry Season Reproductive Depression of Anopheles gambiae in the Sahel. J. Insect Physiol. 2012, 58, 1050–1059. [Google Scholar] [CrossRef]
- Dohm, D.J.; O’Guinn, M.L.; Turell, M.J. Effect of Environmental Temperature on the Ability of Culex pipiens (Diptera: Culicidae) to Transmit West Nile Virus. J. Med. Entomol. 2002, 39, 221–225. [Google Scholar] [CrossRef]
- Reisen, W.K.; Fang, Y.; Martinez, V.M.; Martinez, V.M. Effects of Temperature on the Transmission of West Nile Virus by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 2006, 43, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, E.; Paslaru, A.; Silaghi, C.; Tobler, K.; Glavinic, U.; Torgerson, P.; Mathis, A. Experimental Evaluation of Infection, Dissemination, and Transmission Rates for Two West Nile Virus Strains in European Aedes japonicus under a Fluctuating Temperature Regime. Parasitol. Res. 2018, 117, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Belova, A.; Mills, D.; Hall, R.; St. Julina, A.; Crimmins, A.; Barker, C.; Jones, R. Impacts of Increasing Temperature on the Future Incidence of West Nile Neuroinvasive Disease in the United States. Am. J. Clim. Chang. 2017, 6, 166–216. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, C.J.; Reiskind, M.H.; Pesko, K.N.; Greene, K.E.; Lounibos, L.P. Larval Environmental Temperature and the Susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya Virus. Vector Borne Zoonotic Dis. 2010, 10, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Murdock, C.C.; Blanford, S.; Hughes, G.L.; Rasgon, J.L.; Thomas, M.B. Temperature Alters Plasmodium Blocking by Wolbachia. Sci. Rep. 2014, 4, 3932. [Google Scholar] [CrossRef]
- Wimberly, M.C.; Lamsal, A.; Giacomo, P.; Chuang, T.W. Regional Variation of Climatic Influences on West Nile Virus Outbreaks in the United States. Am. J. Trop. Med. Hyg. 2014, 91, 677–684. [Google Scholar] [CrossRef]
- Landesman, W.J.; Allan, B.F.; Langerhans, R.B.; Knight, T.M.; Chase, J.M. Inter-Annual Associations Between Precipitation and Human Incidence of West Nile Virus in the United States. Vector Borne Zoonotic Dis. 2007, 7, 337–343. [Google Scholar] [CrossRef]
- Shaman, J.; Day, J.F.; Stieglitz, M. Drought-Induced Amplification and Epidemic Transmission of West Nile Virus in Southern Florida. J. Med. Entomol. 2005, 42, 134–141. [Google Scholar] [CrossRef]
- Day, J.F.; Shaman, J. Using Hydrologic Conditions to Forecast the Risk of Focal and Epidemic Arboviral Transmission in Peninsular Florida. J. Med. Entomol. 2008, 45, 458–465. [Google Scholar] [CrossRef]
- Shaman, J.; Harding, K.; Campbell, S.R. Meteorological and Hydrological Influences on the Spatial and Temporal Prevalence of West Nile Virus in Culex Mosquitoes, Suffolk County, New York. J. Med. Entomol. 2011, 48, 867–875. [Google Scholar] [CrossRef]
- Shaman, J.; Day, J.F.; Stieglitz, M. Drought-Induced Amplification of Saint Louis Encephalitis Virus, Florida. Emerg. Infect. Dis. 2002, 8, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P. Climate Change and Mosquito-Borne Disease. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar] [PubMed]
- Paz, S. Climate Change Impacts on West Nile Virus Transmission in a Global Context. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed]
- Paull, S.H.S.; Horton, D.E.; Ashfaq, M.; Rastogi, D.; Kramer, L.D.L.; Diffenbaugh, N.S.N.; Kilpatrick, A.M.A. Drought and Immunity Determine the Intensity of West Nile Virus Epidemics and Climate Change Impacts. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162078. [Google Scholar] [CrossRef]
- Rosà, R.; Marini, G.; Bolzoni, L.; Neteler, M.; Metz, M.; Delucchi, L.; Chadwick, E.A.; Balbo, L.; Mosca, A.; Giacobini, M.; et al. Early Warning of West Nile Virus Mosquito Vector: Climate and Land Use Models Successfully Explain Phenology and Abundance of Culex pipiens Mosquitoes in North-Western Italy. Parasit. Vectors 2014, 7, 1–12. [Google Scholar] [CrossRef]
- Davis, J.K.; Vincent, G.P.; Hildreth, M.B.; Kightlinger, L.; Carlson, C.; Wimberly, M.C.; Davis, A.J.K.; Vincent, G.P.; Michael, B.; Kightlinger, L.; et al. Improving the Prediction of Arbovirus Outbreaks: A Comparison of Climate-Driven Models for West Nile Virus in an Endemic Region of the United States. Acta Trop. 2018, 185, 242–250. [Google Scholar] [CrossRef]
- Paz, S. The West Nile Virus Outbreak in Israel (2000) from a New Perspective: The Regional Impact of Climate Change. Int. J. Environ. Health Res. 2006, 16, 1–13. [Google Scholar] [CrossRef]
- Paz, S.; Malkinson, D.; Green, M.S.; Tsioni, G.; Papa, A.; Danis, K.; Sirbu, A.; Ceianu, C.; Katalin, K.; Ferenczi, E.; et al. Permissive Summer Temperatures of the 2010 European West Nile Fever Upsurge. PLoS ONE 2013, 8, e56398. [Google Scholar] [CrossRef]
- Dodson, B.L.; Kramer, L.D.; Rasgon, J.L. Larval Nutritional Stress Does Not Affect Vector Competence for West Nile Virus (WNV) in Culex tarsalis. Vector Borne Zoonotic Dis. 2011, 11, 1493–1497. [Google Scholar] [CrossRef]
- Alto, B.W.; Lounibos, L.P.; Mores, C.N.; Reiskind, M.H. Larval Competition Alters Susceptibility of Adult Aedes Mosquitoes to Dengue Infection. Proc. R. Soc. B 2008, 275, 463–471. [Google Scholar] [CrossRef]
- Geoghegan, J.L.; Walker, P.J.; Duchemin, J.B.; Jeanne, I.; Holmes, E.C. Seasonal Drivers of the Epidemiology of Arthropod-Borne Viruses in Australia. PLoS Negl. Trop. Dis. 2014, 8, e3325. [Google Scholar] [CrossRef] [PubMed]
- Mamai, W.; Mouline, K.; Parvy, J.P.; Le Lannic, J.; Dabiré, K.R.; Ouédraogo, G.A.; Renault, D.; Simard, F. Morphological Changes in the Spiracles of Anopheles gambiae s.l. (Diptera) as a Response to the Dry Season Conditions in Burkina Faso (West Africa). Parasit. Vectors 2016, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dai, A. Increasing Drought under Global Warming in Observations and Models. Nat. Clim. Chang. 2012, 3, 52–58. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; Van Der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global Warming and Changes in Drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Brackney, D.E.; Foy, B.D.; Olson, K.E. The Effects of Midgut Serine Proteases on Dengue Virus Type 2 Infectivity of Aedes aegypti. Am. J. Trop. Med. Hyg. 2008, 79, 267–274. [Google Scholar] [CrossRef]
- Molina-Cruz, A.; Gupta, L.; Richardson, J.; Bennett, K.; Black IV, W.; Barillas-Mury, C. Effect of Mosquito Midgut Trypsin Activity on Dengue-2 Virus Infection and Dissemination in Aedes Aegypti. Am. J. Trop. Med. Hyg. 2005, 72, 631–637. [Google Scholar] [CrossRef]
- Scott, J.C.; Brackney, D.E.; Campbell, C.L.; Bondu-Hawkins, V.; Hjelle, B.; Ebel, G.D.; Olson, K.E.; Blair, C.D. Comparison of Dengue Virus Type 2-Specific Small RNAs from RNA Interference-Competent and -Incompetent Mosquito Cells. PLoS Negl. Trop. Dis. 2010, 4, e848. [Google Scholar] [CrossRef]
- Fitzpatrick, K.A.; Deardorff, E.R.; Pesko, K.; Brackney, D.E.; Zhang, B.; Bedrick, E.; Shi, P.Y.; Ebel, G.D. Population Variation of West Nile Virus Confers a Host-Specific Fitness Benefit in Mosquitoes. Virology 2010, 404, 89–95. [Google Scholar] [CrossRef]
- Alpert, P. Constraints of Tolerance: Why Are Desiccation-Tolerant Organisms so Small or Rare? J. Exp. Biol. 2006, 209, 1575–1584. [Google Scholar] [CrossRef]
- McCluney, K.E. Implications of Animal Water Balance for Terrestrial Food Webs. Curr. Opin. Insect Sci. 2017, 23, 13–21. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Sylvestre, G.; Gandini, M.; Koella, J.C. The Influence of Dengue Virus Serotype-2 Infection on Aedes aegypti (Diptera: Culicidae) Motivation and Avidity to Blood Feed. PLoS ONE 2013, 8, e65252. [Google Scholar] [CrossRef] [PubMed]
- Van der Linde, T.C.D.; Hewitt, P.H.; Nel, A.; Van der Westhuizen, M.C. The Influence of Different Constant Temperatures and Saturation Deficits on the Survival of Adult Culex (Culex) theileri Theobald (Diptera: Culicidae) in the Laboratory. J. Entomol. Soc. South. Afr. 1990, 53, 57–63. [Google Scholar]
- Gardner, A.M.; Muturi, E.J.; Overmier, L.D.; Allan, B.F. Large-Scale Removal of Invasive Honeysuckle Decreases Mosquito and Avian Host Abundance. Ecohealth 2017, 14, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Chowdhury, V.; Faria, S.; Huda, M.M.; Laila, R.; Maheswary, N.P.; Dash, A.P. How Dengue Vector Aedes albopictus (Diptera: Culicidae) Survive during the Dry Season in Dhaka City, Bangladesh? J. Vector Borne Dis. 2014, 51, 179–187. [Google Scholar] [PubMed]
- Lalubin, F.; Delédevant, A.; Glaizot, O.; Christe, P. Natural Malaria Infection Reduces Starvation Resistance of Nutritionally Stressed Mosquitoes. J. Anim. Ecol. 2014, 83, 850–857. [Google Scholar] [CrossRef]
- Ross, P.A.; Endersby, N.M.; Hoffmann, A.A. Costs of Three Wolbachia Infections on the Survival of Aedes aegypti Larvae under Starvation Conditions. PLoS Negl. Trop. Dis. 2016, 10, e0004320. [Google Scholar] [CrossRef]
- Sawabe, K.; Mogi, M. Differences in Energy Metabolism and Adult Desiccation Resistance among Three Aedes (Stegomyia) Species (Diptera: Culicidae) from South Sulawesi, Indonesia. J. Med. Entomol. 1999, 36, 101–107. [Google Scholar] [CrossRef]
- Reisen, W.K.; Meyer, R.P.; Milby, M.M. Patterns of Fructose Feeding by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 1986, 23, 366–373. [Google Scholar] [CrossRef]
- Foster, W.A. Mosquito Sugar Feeding and Reproductive Energetics. Annu. Rev. Entomol. 1995. [Google Scholar] [CrossRef]
- Rodriguez, M.H.; Yuval, B.; Martinez-Ibarra, J.A.; Arredondo-Jimenez, J.I. Influence of Plant Abundance on Nectar Feeding by Aedes aegypti (Diptera: Culicidae) in Southern Mexico. J. Med. Entomol. 1997, 34, 589–593. [Google Scholar]
- Edman, J.D.; Strickman, D.; Kittayapong, P.; Scott, T.W. Female Aedes aegypti (Diptera: Culicidae) in Thailand Rarely Feed on Sugar. J. Med. Entomol. 1992, 29, 1035–1038. [Google Scholar] [CrossRef] [PubMed]
- Briegel, H. Metabolic Relationship between Female Body Size, Reserves, and Fecundity of Aedes aegypti. J. Insect Physiol. 1990, 1990. 36, 165–172. [Google Scholar] [CrossRef]
- Jones, M.D.R.; Gubbins, S.J. Changes in the Circadian Flight Activity of the Mosquito Anopheles gambiae in Relation to Insemination, Feeding and Oviposition. Physiol. Entomol. 1978, 3, 213–220. [Google Scholar] [CrossRef]
- Aboagye-Antwi, F.; Tripet, F. Effects of Larval Growth Condition and Water Availability on Desiccation Resistance and Its Physiological Basis in Adult Anopheles gambiae sensu stricto. Malar. J. 2010, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.C.; Edman, J.D.; Scott, T.W. Why Do Female Aedes aegypti (Diptera: Culicidae) Feed Preferentially and Frequently on Human Blood? J. Med. Entomol. 2001, 38, 411–422. [Google Scholar] [CrossRef]
- Huestis, D.L.; Yaro, A.S.; Traore, A.I.; Adamou, A.; Kassogue, Y.; Diallo, M.; Timbine, S.; Dao, A.; Lehmann, T. Variation in Metabolic Rate of Anopheles gambiae and A. arabiensis in a Sahelian Village. J. Exp. Biol. 2011, 214, 2345–2353. [Google Scholar] [CrossRef]
- Dow, R.P.; Gerrish, G.M. Day-to-Day Change in Relative Humidity and the Activity of Culex nigripalpus (Diptera: Culicidae). Ann. Entomol. Soc. Am. 1970, 63, 995–999. [Google Scholar] [CrossRef]
- Ritchie, S.A.; Townsend, M.; Paton, C.J.; Callahan, A.G.; Hoffmann, A.A. Application of Wmelpop Wolbachia Strain to Crash Local Populations of Aedes aegypti. PLoS Negl. Trop. Dis. 2015, 9, e0003930. [Google Scholar] [CrossRef]
- Reisen, W.K.; Mahmood, F.; Parveen, T. Seasonal Trends in Population Size and Survivorship of Anopheles culicifacies, An. stephensi, and An. subpictus (Diptera: Culicidae) in Rural Punjab Province, Pakistan. J. Med. Entomol. 1982, 19, 86–97. [Google Scholar] [CrossRef]
- Lyons, C.L.; Coetzee, M.; Terblanche, J.S.; Chown, S.L. Desiccation Tolerance as a Function of Age, Sex, Humidity and Temperature in Adults of the African Malaria Vectors Anopheles arabiensis and Anopheles funestus. J. Exp. Biol. 2014, 217, 3823–3833. [Google Scholar] [CrossRef]
- Kumar, M. Effect of Temperature and Humidity on Life Cycle Duration of Culex quinquefasciatus Say (Diptera: Culicidae) at Muzaffarpur (Bihar), India. Adv. Biores. 2015, 6, 103–105. [Google Scholar]
- Leeson, H.S. Longevity of Anopheles maculipennis Race atroparvus, Van Thiel, at Controlled Temperature and Humidity after One Blood Meal. Bull. Entomol. Res. 1939, 30, 295–301. [Google Scholar] [CrossRef]
- Martin, L.J.; Adams, R.I.; Bateman, A.; Bik, H.M.; Hawks, J.; Hird, S.M.; Hughes, D.; Kembel, S.W.; Kinney, K.; Kolokotronis, S.; et al. Evolution of the Indoor Biome. Trends Ecol. Evol. 2015, 30, 223–232. [Google Scholar] [CrossRef]
- Chase, J.M.; Knight, T.M. Drought-Induced Mosquito Outbreaks in Wetlands. Ecol. Lett. 2003, 6, 1017–1024. [Google Scholar] [CrossRef]
- Ruiz, M.O.; Chaves, L.F.; Hamer, G.L.; Sun, T.; Brown, W.M.; Walker, E.D.; Haramis, L.; Goldberg, T.L.; Kitron, U.D. Local Impact of Temperature and Precipitation on West Nile Virus Infection in Culex Species Mosquitoes in Northeast Illinois, USA. Parasit. Vectors 2010, 3, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.F.; Kitron, U.D. Weather Variability Impacts on Oviposition Dynamics of the Southern House Mosquito at Intermediate Time Scales. Bull. Entomol. Res. 2010, 101, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Mbepera, S.; Nkwengulila, G.; Peter, R.; Mausa, E.A.; Mahande, A.M.; Coetzee, M.; Kweka, E.J. The Influence of Age on Insecticide Susceptibility of Anopheles arabiensis during Dry and Rainy Seasons in Rice Irrigation Schemes of Northern Tanzania. Malar. J. 2017, 16, 1–9. [Google Scholar] [CrossRef]
- Besansky, N.J.B. Malaria: How Vector Mosquitoes Beat the Heat. Nature 2014, 516, 334–336. [Google Scholar] [CrossRef]
- Slaff, M.; Crans, W.J. The Activity and Physiological Status of Pre- and Posthibernating Culex salinarius (Diptera: Culicidae) Populations. J. Med. Entomol 1981, 18, 65–68. [Google Scholar] [CrossRef]
- Hidalgo, K.; Montazeau, C.; Siaussat, D.; Braman, V.; Trabalon, M.; Simard, F.; Renault, D.; Mouline, K. Distinct Physiological, Biochemical and Morphometric Adjustments in the Malaria Vectors Anopheles gambiae and An. coluzzii as Means to Survive to Dry Season Conditions in Burkina Faso. J. Exp. Biol. 2018, 221, jeb.174433. [Google Scholar] [CrossRef]
- Huestis, D.L.; Dao, A.; Diallo, M.; Sanogo, Z.L.; Samake, D.; Yaro, A.S.; Ousman, Y.; Linton, Y.-M.; Krishna, A.; Veru, L.; et al. Windborne Long-Distance Migration of Malaria Mosquitoes in the Sahel. Nature 2019, 574, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Arcaz, A.C.; Huestis, D.L.; Dao, A.; Yaro, A.S.; Diallo, M.; Andersen, J.; Blomquist, G.J.; Lehmann, T. Desiccation Tolerance in Anopheles coluzzii: The Effects of Spiracle Size and Cuticular Hydrocarbons. J. Exp. Biol. 2016, 219, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Machado-Allison, C.E.; Craig, G.B. Geographic Variation in Resistance to Desiccation in Aedes aegypti and A. atropalpus (Diptera: Culicidae). Ann. Entomol. Soc. Am. 1972, 65, 542–547. [Google Scholar] [CrossRef]
- Carrieri, M.; Fariselli, P.; Maccagnani, B.; Angelini, P.; Calzolari, M.; Bellini, R. Weather Factors Influencing the Population Dynamics of Culex pipiens (Diptera: Culicidae) in the Po Plain Valley, Italy (1997–2011). Environ. Entomol. 2014, 43, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Miyagi, I.; Abadi, K.; Syafruddin. Inter- and Intraspecific Variation in Resistance to Desiccation by Adult aedes (Stegomyia) spp. (Diptera: Culicidae) from Indonesia. J Med Entomol. 1996, 33, 53–57. [Google Scholar] [CrossRef]
- Hidalgo, K.; Siaussat, D.; Braman, V.; Dabiré, K.R.; Simard, F.; Mouline, K.; Renault, D. Comparative Physiological Plasticity to Desiccation in Distinct Populations of the Malarial Mosquito Anopheles coluzzii. Parasit. Vectors 2016, 9, 1–13. [Google Scholar] [CrossRef]
- Gibbs, A.G.; Chippindale, A.K.; Rose, M.R. Physiological Mechanisms of Evolved Desiccation Resistance in Drosophila melanogaster. J. Exp. Biol. 1997, 200, 1821–1832. [Google Scholar]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Niehuis, O. Phylogenomics Resolves the Timing and Pattern of Insect Evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; et al. Episodic Radiations in the Fly Tree of Life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef]
- Yeates, D.K.; Wiegmann, B.M. Congruence and Controversy: Toward a Higher-Level Phylogeny of Diptera. Annu. Rev. Entomol. 1999, 44, 397–428. [Google Scholar] [CrossRef]
- Wiegmann, B.M.; Richards, S. Genomes of Diptera. Curr. Opin. Insect Sci. 2018, 25, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, J.; Grushko, O.G.; Besansky, N.J. Analysis of the Complete Mitochondrial DNA from Anopheles funestus: An Improved Dipteran Mitochondrial Genome Annotation and a Temporal Dimension of Mosquito Evolution. Mol. Phylogenet. Evol. 2006, 39, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Samuel, G.H.; Wiley, M.R.; Badawi, A.; Adelman, Z.N.; Myles, K.M. Yellow Fever Virus Capsid Protein Is a Potent Suppressor of RNA Silencing That Binds Double-Stranded RNA. Proc. Natl. Acad. Sci. USA 2016, 113, 13863–13868. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Guerbois, M.; Seymour, R.L.; Spratt, H.; Weaver, S.C. Vector-Borne Transmission Imposes a Severe Bottleneck on an RNA Virus Population. PLoS Pathog. 2012, 8, e1002897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Aksoy, E.; Girke, T.; Raikhel, A.S.; Karginov, F.V. Transcriptome-Wide MicroRNA and Target Dynamics in the Fat Body during the Gonadotrophic Cycle of Aedes aegypti. Proc. Natl. Acad. Sci. USA 2017, 114, 1895–1903. [Google Scholar] [CrossRef] [PubMed]
- Fouet, C.; Gray, E.; Besansky, N.J.; Costantini, C. Adaptation to Aridity in the Malaria Mosquito Anopheles gambiae: Chromosomal Inversion Polymorphism and Body Size Influence Resistance to Desiccation. PLoS ONE 2012, 7, e34841. [Google Scholar] [CrossRef]
- Ayala, D.; Zhang, S.; Chateau, M.; Fouet, C.; Morlais, I.; Costantini, C.; Hahn, M.W.; Besansky, N. Association Mapping Desiccation Resistance within Chromosomal Inversions in the African Malaria Vector Anopheles gambiae. Mol. Ecol. 2018, 28. [Google Scholar] [CrossRef]
- Rocca, K.A.; Gray, E.M.; Costantini, C.; Besansky, N.J. 2La Chromosomal Inversion Enhances Thermal Tolerance of Anopheles gambiae Larvae. Malar. J. 2009, 8, 1–7. [Google Scholar] [CrossRef]
- Gray, E.M.; Rocca, K.A.; Costantini, C.; Besansky, N.J. Inversion 2La Is Associated with Enhanced Desiccation Resistance in Anopheles gambiae. Malar. J. 2009, 8, 215. [Google Scholar] [CrossRef]
- White, B.J.; Collins, F.H.; Besansky, N.J. Evolution of Anopheles gambiae in Relation to Humans and Malaria. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 111–132. [Google Scholar] [CrossRef]
- Ayala, D.; Acevedo, P.; Pombi, M.; Dia, I.; Boccolini, D.; Costantini, C.; Simard, F.; Fontenille, D. Chromosome Inversions and Ecological Plasticity in the Main African Malaria Mosquitoes. Evolution 2017, 71, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Sayono, S.; Nurullita, U.; Sumanto, D.; Handoyo, W. Altitudinal Distribution of Aedes Indices during Dry Season in the Dengue Endemic Area of Central Java, Indonesia. Ann. Parasitol. 2017, 63, 213–221. [Google Scholar] [PubMed]
- Touré, Y.; Petrarca, V.; Traoré, S.; Coulibaly, A.; Maiga, H.; Sankaré, O.; Sow, M.; Di Deco, M.; Coluzzi, M. The Distribution and Inversion Polymorphism of Chromosomally Recognized Taxa of the Anopheles gambiae Complex in Mali, West Africa. Parassitologia 1999, 40, 477–511. [Google Scholar]
- Hidalgo, K.; Mouline, K.; Mamai, W.; Foucreau, N.; Dabiré, K.R.; Bouchereau, A.; Simard, F.; Renault, D. Novel Insights into the Metabolic and Biochemical Underpinnings Assisting Dry-Season Survival in Female Malaria Mosquitoes of the Anopheles gambiae Complex. J. Insect Physiol. 2014, 70, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.M.; Bradley, T.J. Physiology of Desiccation Resistance in Anopheles gambiae and Anopheles arabiensis. Am. J. Trop. Med. Hyg. 2005, 73, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, B.N.; Srivastava, A.; Kalra, N.L.; Subbarao, S.K. Spiracular Indices in Anopheles stephensi: A Taxonomic Tool to Identify Ecological Variants. J. Med. Entomol. 2003, 40, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-H.; Marinotti, O.; Vardo-Zalik, A.; Boparai, R.; Yan, G. Genome-Wide Transcriptional Analysis of Genes Associated with Acute Desiccation Stress in Anopheles gambiae. PLoS ONE 2011, 6, e26011. [Google Scholar] [CrossRef]
- Mamai, W.; Mouline, K.; Blais, C.; Larvor, V.; Dabiré, K.R.; Ouedraogo, G.A.; Simard, F.; Renault, D. Metabolomic and Ecdysteroid Variations in Anopheles gambiae s.l. Mosquitoes Exposed to the Stressful Conditions of the Dry Season in Burkina Faso, West Africa. Physiol. Biochem. Zool. 2014, 87, 486–497. [Google Scholar] [CrossRef]
- Colpitts, T.M.; Cox, J.; Vanlandingham, D.L.; Feitosa, F.M.; Cheng, G.; Kurscheid, S.; Wang, P.; Krishnan, M.N.; Higgs, S.; Fikrig, E. Alterations in the Aedes aegypti Transcriptome during Infection with West Nile, Dengue and Yellow Fever Viruses. PLoS Pathog. 2011, 7, e1002189. [Google Scholar] [CrossRef]
- Briegel, H.; Hörler, E. Multiple Blood Meals as a Reproductive Strategy in Anopheles (Diptera: Culicidae). J. Med. Entomol. 1993, 30, 975–985. [Google Scholar] [CrossRef]
- Lea, A.O.; Briegel, H.; Lea, H.M. Arrest, Resorption, or Maturation of Oocytes in Aedes aegypti: Dependence on the Quantity of Blood and the Internval between Blood Meals. Physiol. Entomol. 1978, 3, 309–316. [Google Scholar] [CrossRef]
- Nayar, J.K.; Sauerman, D.M. The Effects of Nutrition on Survival and Fecundity on Florida Mosquitoes. J. Med. Ent. 1975, 12, 99–103. [Google Scholar] [CrossRef]
- Lehane, M.J. The Biology of Blood-Sucking in Insects, 2nd ed.; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Sanders, T.; Scanlon, V. Essentials of Anatomy and Physiology Scanlon; FA Davis Co.: Philadelphia, PA, USA, 2007. [Google Scholar]
- Williams, J.C.; Hagedorn, H.H.; Beyenbach, K.W. Dynamic Changes in Flow Rate and Composition of Urine during the Post-Bloodmeal Diuresis in Aedes aegypti (L.). J. Comp. Physiol. B 1983, 153, 257–265. [Google Scholar] [CrossRef]
- Li, Y.; Piermarini, P.M.; Esquivel, C.J.; Drumm, H.E.; Schilkey, F.D.; Hansen, I.A. RNA-Seq Comparison of Larval and Adult Malpighian Tubules of the Yellow Fever Mosquito Aedes aegypti Reveals Life Stage-Specific Changes in Renal Function. Front. Physiol. 2017, 8, 1–13. [Google Scholar]
- Madeira, N.G.; Macharelli, C.A.; Carvalho, L.R. Variation of the Oviposition Preferences of Aedes aegypti in Function of Substratum and Humidity. Mem. Inst. Oswaldo Cruz 2002, 97, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.J.; Younger, M.A.; Vosshall, L.B. The Ion Channel ppk301 Controls Freshwater Egg-Laying in the Mosquito Aedes aegypti. Elife 2019, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Mayne, B. Notes on the Influence of Temperature and Humidity on Oviposition and Early Life of Anopheles. Public Health Rep. 1926, 41, 986–990. [Google Scholar] [CrossRef]
- Detinova, T.S.; Beklemishev, W.N.; Bertram, D.S. Age-Grouping Metohds in Diptera of Medical Importance with Special Reference to Some Vectors of Malaria. World Heal. Organ. Monogr. Ser. 1962, 47, 1–213. [Google Scholar]
- Costa, E.A.P.D.A.; Santos, E.M.D.M.; Correia, J.C.; Albuquerque, C.M.R.D. Impact of Small Variations in Temperature and Humidity on the Reproductive Activity and Survival of Aedes aegypti (Diptera, Culicidae). Rev. Bras. Entomol. 2010, 54, 488–493. [Google Scholar] [CrossRef]
- Tunggul Satoto, T.B.; R Umniyati, S.; Suardipa, A.; Sintorini, M. Effects of Temperature, Relative Humidity, and DEN-2 Virus Transovarial Infection on Viability of Aedes aegypti. J. Kesehat. Masy. Nas. 2013, 7, 331–336. [Google Scholar] [CrossRef]
- Swellengrebel, N.H. La Dissociation Des Fonctions Sexuelles et Nutritives (Dissociation Gono-Trophique) d’Anopheles maculipennis Comme Cause Du Paludisme Dans Les Pays-Bas et Ses Rapports Avec “l’infection Domiciliaire”. Ann. Inst. Pasteur (Paris) 1929, 43, 1370–1389. [Google Scholar]
- Mitchell, C.J.; Briegel, H. Fate of the Blood Meal in Force-Fed, Diapausing Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 1989, 26, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Sota, T.; Mogi, M. Survival Time and Resistance to Desiccation of Diapause and Non-diapause Eggs of Temperate Aedes (Stegomyia) Mosquitoes. Entomol. Exp. Appl. 1992, 63, 155–161. [Google Scholar] [CrossRef]
- Mala, A.O.; Irungu, L.W.; Mitaki, E.K.; Shililu, J.I.; Mbogo, C.M.; Njagi, J.K.; Githure, J.I. Gonotrophic Cycle Duration, Fecundity and Parity of Anopheles gambiae Complex Mosquitoes during an Extended Period of Dry Weather in a Semi Arid Area in Baringo County, Kenya. Int. J. Mosq. Res. IJMR 2014, 28, 28–34. [Google Scholar]
- Farnesi, L.C.; Menna-Barreto, R.F.S.; Martins, A.J.; Valle, D.; Rezende, G.L. Physical Features and Chitin Content of Eggs from the Mosquito Vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus: Connection with Distinct Levels of Resistance to Desiccation. J. Insect Physiol. 2015, 83, 43–52. [Google Scholar] [CrossRef]
- Farnesi, L.C.; Vargas, H.C.M.; Valle, D.; Rezende, G.L. Darker Eggs of Mosquitoes Resist More to Dry Conditions: Melanin Enhances Serosal Cuticle Contribution in Egg Resistance to Desiccation in Aedes, Anopheles and Culex Vectors. PLoS Negl. Trop. Dis. 2017, 11, e0006063. [Google Scholar] [CrossRef]
- Vargas, H.C.M.; Farnesi, L.C.; Martins, A.J.; Valle, D.; Rezende, G.L. Serosal Cuticle Formation and Distinct Degrees of Desiccation Resistance in Embryos of the Mosquito Vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus. J. Insect Physiol. 2014, 62, 54–60. [Google Scholar] [CrossRef]
- Faull, K.J.; Webb, C.; Williams, C.R. Desiccation Survival Time for Eggs of a Widespread and Invasive Australian Mosquito Species, Aedes (Finlaya) notoscriptus (Skuse). J. Vector Ecol. 2016, 41, 55–62. [Google Scholar] [CrossRef]
- Urbanski, J.M.; Benoit, J.B.; Michaud, M.R.; Denlinger, D.L.; Armbruster, P. The Molecular Physiology of Increased Egg Desiccation Resistance during Diapause in the Invasive Mosquito, Aedes albopictus. Proc. R. Soc. B 2010, 2683–2692. [Google Scholar] [CrossRef]
- Faull, K.J.; Williams, C.R. Intraspecific Variation in Desiccation Survival Time of Aedes aegypti (L.) Mosquito Eggs of Australian Origin. J. Vector Ecol. 2015, 40, 292–300. [Google Scholar] [CrossRef]
- Ewing, D.A.; Cobbold, C.A.; Purse, B.V.; Nunn, M.A.; White, S.M. Modelling the Effect of Temperature on the Seasonal Population Dynamics of Temperate Mosquitoes. J. Theor. Biol. 2016, 400, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Bolzoni, L.; Chadwick, E.A.; Capelli, G.; Montarsi, F.; Grisenti, M.; De La Puente, J.M.; Muñoz, J.; Figuerola, J.; Soriguer, R.; et al. Understanding West Nile Virus Ecology in Europe: Culex pipiens Host Feeding Preference in a Hotspot of Virus Emergence. Parasit. Vectors 2015, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Teets, N.M.; Kawarasaki, Y.; Lee, R.E.; Denlinger, D.L. Expression of Genes Involved in Energy Mobilization and Osmoprotectant Synthesis during Thermal and Dehydration Stress in the Antarctic Midge, Belgica antarctica. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2013, 183, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.R.; Dethier, V.G. The Regulation of Taste Thresholds for Sugars in the Blowfly. J. Insect Physiol. 1957. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Crowe, L.M. Anhydrobiosis. Annu. Rev. Physiol. 1992, 54, 579–599. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Maibach, H.I. A Study of the Probing Response of Aedes aegypti. 4. Effect of Dry and Moist Heat on Probing. J. Econ. Entomol. 1971, 64, 442–443. [Google Scholar] [CrossRef]
- Khan, A.A.; Maibach, H.I. Quantitation of Effect of Several Stimuli on Landing and Probing by Aedes aegypti. J. Econ. Entomol. 1966, 59, 902–905. [Google Scholar] [CrossRef]
- Khan, A.A.; Maibach, H.I. A Study of the Probing Response of Aedes aegypti 1. Effect of Nutrition on Probing. J. Econ. Entomol. 1970, 63, 974–976. [Google Scholar] [CrossRef]
- Grossman, G.L.; Pappas, L.G. Human Skin Temperature and Mosquito (Diptera: Culicidae) Blood Feeding Rate. J. Med. Entomol. 1991, 28, 456–460. [Google Scholar] [CrossRef]
- Eiras, A.E.; Jepson, P.C. Responses of Female Aedes aegypti (Diptera: Culicidae) to Host Odours and Convection Currents Using an Olfactometer Bioassay. Bull. Entomol. Res. 1994, 84, 207–211. [Google Scholar] [CrossRef]
- Van Breugel, F.; Riffell, J.; Fairhall, A.; Dickinson, M.H. Mosquitoes Use Vision to Associate Odor Plumes with Thermal Targets. Curr. Biol. 2015, 25, 2123–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Benton, R. Functional Architecture of Olfactory Ionotropic Glutamate Receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enjin, A.; Zaharieva, E.E.; Frank, D.D.; Mansourian, S.; Suh, G.S.B.; Gallio, M.; Stensmyr, M.C. Humidity Sensing in Drosophila. Curr. Biol. 2016, 55, 9557–9561. [Google Scholar] [CrossRef] [PubMed]
- Benoit, J.B.; Lopez-Martinez, G.; Phillips, Z.P.; Patrick, K.R.; Denlinger, D.L. Heat Shock Proteins Contribute to Mosquito Dehydration Tolerance. J. Insect Physiol. 2010, 56, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.S.; Larter, N.K.; Chahda, J.S.; Rioux, D.; Haven, N.; States, U. Humidity Response Depends on the Small Soluble Protein Obp59a in Drosophila. eLife 2018, 7, e39249. [Google Scholar] [CrossRef] [PubMed]
- Qualls, W.A.; Day, J.F.; Xue, R.-D.; Bowers, D.F. Sindbis Virus Infection Alters Blood Feeding Responses and DEET Repellency in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 418–423. [Google Scholar] [CrossRef]
- Ranson, H.; Lissenden, N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation That Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef]
- Toé, K.H.; Jones, C.M.; N’fale, S.; Ismai, H.M.; Dabiré, R.K.; Ranson, H. Increased Pyrethroid Resistance in Malaria Vectors and Decreased Bed Net Effectiveness Burkina Faso. Emerg. Infect. Dis. 2014, 20, 1691–1696. [Google Scholar] [CrossRef]
- Wang, G.; Minnis, R.B.; Belant, J.L.; Wax, C.L. Dry Weather Induces Outbreaks of Human West Nile Virus Infections. BMC Infect. Dis. 2010, 10, 38. [Google Scholar] [CrossRef]
- Eikenberry, S.E.; Gumel, A.B. Mathematical Modeling of Climate Change and Malaria Transmission Dynamics: A Historical Review. J. Math. Biol. 2018, 77, 1–77. [Google Scholar] [CrossRef]
- Tompkins, A.M.; Ermert, V. A Regional-Scale, High Resolution Dynamical Malaria Model That Accounts for Population Density, Climate and Surface Hydrology. Malar. J. 2013, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Christiansen-Jucht, C.; Parham, P.E.; Saddler, A.; Koella, J.C.; Basáñez, M.G. Temperature during Larval Development and Adult Maintenance Influences the Survival of Anopheles gambiae s.s. Parasit. Vectors 2014, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lega, J.; Brown, H.E.; Barrera, R. Aedes aegypti (Diptera: Culicidae) Abundance Model Improved with Relative Humidity and Precipitation-Driven Egg Hatching. J. Med. Entomol. 2017, 54, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, A.E.; Schmitz, O.J. Climate Change, Nutrition, and Bottom-Up and Top-Down Food Web Processes. Trends Ecol. Evol. 2016, 31, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.; Mercier, A.; Lepers, C.; Hoy, D.; Duituturaga, S.; Benyon, E.; Guillaumot, L.; Souarès, Y. Concurrent Outbreaks of Dengue, Chikungunya and Zika Virus Infections–An Unprecedented Epidemic Wave of Mosquito-Borne Viruses in the Pacific 2012–2014. Eurosurveillance 2014, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Thu, H.M.; Aye, K.M.; Thein, S. The Effect of Temperature and Humidity on Dengue Virus Propagation in Aedes aegypti Mosquitos. Southeast Asian J. Trop. Med. Public Health 1998, 29, 280–284. [Google Scholar] [PubMed]
- Slatyer, R.O. Plant-Water Relationships; Academic Press: London, UK, 1967. [Google Scholar]
- Rowley, W.A.; Graham, C.L. The Effect of Temperature and Relative Humidity on the Flight Performance of Female Aedes aegypti. J. Insect Physiol. 1968, 14, 1251–1257. [Google Scholar] [CrossRef]
- Anderson, D.B. Relative Humidity or Vapor Pressure Deficit. Ecology 1936, 17, 277–282. [Google Scholar] [CrossRef]
- Alduchov, O.A.; Eskridge, R.E. Improved Magnus Form Approximation of Saturation Vapor Pressure. J. Appl. Meteorol. 1996, 35, 601–609. [Google Scholar] [CrossRef]
- Buck, A.L. Model Cr-1a Hygrometer with Autofill Operating Manual; Buck Research Instruments LLC: Aurora, CO, USA, 1996. [Google Scholar]
- Buck, A.L. New Equations for Computing Vapor Pressure and Enhancement Factor. J. Appl. Meteorol. 1981, 20, 1527–1532. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.M.; Koop, T. Review of the Vapour Pressures of Ice and Supercooled Water for Atmospheric Applications. Q. J. R. Meteorol. Soc. 2005, 131, 1539–1565. [Google Scholar] [CrossRef]
- Tetens, O. Uber Einige Meteorologische Begriffe. Z. Geophys. 1930, 6, 297–309. [Google Scholar]
- Principles of Environmental Physics. In Principles of Environmental Physics (Fourth Edition); Monteith, J.L.; Unsworth, M.H. (Eds.) Academic Press: Boston, MA, USA, 2013; pp. 1–401. [Google Scholar]
- Murray, F.W. On the Computation of Saturation Vapor Pressure. J. Appl. Meteorol. 1967, 6, 203–204. [Google Scholar] [CrossRef]
- Shipp, J.L.; Grace, B.W.; Schaalje, G.B. Effects of Microclimate on Daily Flight Activity of Simulium arcticum Malloch (Diptera: Simuliidae). Int. J. Biometeorol. 1987, 31, 9–20. [Google Scholar] [CrossRef]
- Wharton, G.W. Water Balance of Insects: In Comprehensive Insect Physiology Biochemistry and Pharmacology, 4th ed.; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985. [Google Scholar]
- de la Vega, G.J.; Schilman, P.E. Using Eco-Physiological Traits to Understand the Realized Niche: The Role of Desiccation Tolerance in Chagas Disease Vectors. Oecologia 2017, 185, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Islam, M.S. Life-History Traits of Two Medically Important Insects Culex quinquefasciatus Say and Musca domestica L. Influenced by Temperature and Humidity. Int. J. Sci. Eng. Appl. Sci. 2017, 3, 135–142. [Google Scholar]
- Schmalhausen, I.I. Factors of Evolution: The Theory of Stabilizing Selection; Blakiston: Oxford, UK, 1949. [Google Scholar]
- Wharton, G.W.; Richards, G.R. Water Vapor Exchange. Annu. Rev. Entomol. 1978, 23, 309–328. [Google Scholar] [CrossRef]
- Carrington, L.B.; Armijos, M.V.; Lambrechts, L.; Barker, C.M.; Scott, T.W. Effects of Fluctuating Daily Temperatures at Critical Thermal Extremes on Aedes aegypti Life-History Traits. PLoS ONE 2013, 8, e58824. [Google Scholar] [CrossRef]
- Parker, A.H. The Effect of a Difference in Temperature and Humidity on Certain Reactions of Female Aëdes aegypti (L.). Bull. Entomol. Res. 1952, 43, 221–229. [Google Scholar] [CrossRef]
- Rosendale, A.J.; Romick-Rosendale, L.E.; Watanabe, M.; Dunlevy, M.E.; Benoit, J.B. Mechanistic Underpinnings of Dehydration Stress in the American Dog Tick Revealed through RNA-Seq and Metabolomics. J. Exp. Biol. 2016, 219, 1808–1819. [Google Scholar] [CrossRef]
- Hercus, M.J.; Hoffmann, A.A. Desiccation Resistance in Interspecific Drosophila Crosses: Genetic Interactions and Trait Correlations. Genetics 1999, 151, 1493–1502. [Google Scholar] [PubMed]
- How, Y.-F.; Lee, C.-Y. Effects of Temperature and Humidity on the Survival and Water Loss of Cimex hemipterus (Hemiptera: Cimicidae). J. Med. Entomol. 2010, 47, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Shipp, J.L.; Gillespie, T.J. Influence of Temperature and Water Vapor Pressure Deficit on Survival of Frankliniella occidentalis (Thysanoptera, Thripidae). Environ. Entomol. 1993, 22, 726–732. [Google Scholar] [CrossRef]
- Boudreaux, H.B. The Effect of Relative Humidity on Egg-Laying, Hatching, and Survival in Various Spider Mites. J. Insect Physiol. 1958, 2, 65–72. [Google Scholar] [CrossRef]
- Bezerra Da Silva, C.S.; Price, B.E.; Walton, V.M. Water-Deprived Parasitic Wasps (Pachycrepoideus vindemmiae) Kill More Pupae of a Pest (Drosophila suzukii) as a Water-Intake Strategy. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chown, S.L.; Nicolson, S. Metabolism and Gas Exchange: In Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, UK; New York, NY, USA, 2004. [Google Scholar]
- Marques, J.T.; Imler, J.L. The Diversity of Insect Antiviral Immunity: Insights from Viruses. Curr. Opin. Microbiol. 2016, 32, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Gatton, M.L.; Chitnis, N.; Churcher, T.; Donnelly, M.J.; Ghani, A.C.; Godfray, H.C.J.; Gould, F.; Hastings, I.; Marshall, J.; Ranson, H.; et al. The Importance of Mosquito Behavioural Adaptations to Malaria Control in Africa. Evolution 2013, 67, 1218–1230. [Google Scholar] [CrossRef]
- Riehle, M.M.; Bukhari, T.; Gneme, A.; Guelbeogo, W.M.; Coulibaly, B.; Fofana, A.; Pain, A.; Bischoff, E.; Renaud, F.; Beavogui, A.H.; et al. The Anopheles gambiae 2La Chromosome Inversion Is Associated with Susceptibility to Plasmodium falciparum in Africa. Elife 2017, 6, 1–24. [Google Scholar] [CrossRef]
- Brooke, B.D.; Hunt, R.H.; Chandre, F.; Carnevale, P.; Coetzee, M. Stable Chromosomal Inversion Polymorphisms and Insecticide Resistance in the Malaria Vector Mosquito Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 568–573. [Google Scholar] [CrossRef]
- Koekemoer, L.L.; Spillings, B.L.; Christian, R.N.; Lo, T.-C.M.; Kaiser, M.L.; Norton, R.A.I.; Oliver, S.V.; Choi, K.S.; Brooke, B.D.; Hunt, R.H.; et al. Multiple Insecticide Resistance in Anopheles gambiae (Diptera: Culicidae) from Pointe Noire, Republic of the Congo. Vector Borne Zoonotic Dis. 2011, 11, 1193–1200. [Google Scholar] [CrossRef]
- Suleman, M. Intraspecific Variation in the Reproductive Capacity of Anopheles stephensi (Diptera: Culicidae). J. Med. Entomol. 1990, 27, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Naksathit, A.T.; Edman, J.D.; Scott, T.W. Utilization of Human Blood and Sugar as Nutrients by Female Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 1999, 36, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Qualls, W.A.; Xue, R.; Beier, J.C.; Debboun, M. Alterations of Blood-Feeding Behavior and Repellent Response of Pathogen-Infected Biting Flies, 2nd ed.; Debboun, M., Frances, S.P., Strickman, D.A., Eds.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Wekesa, J.W.; Copeland, R.S.; Mwangi, R.W. Effect of Plasmodium falciparum on Blood Feeding Behavior of Naturally Infected Anopheles Mosquitoes in Western Kenya. Am. J. Trop. Med. Hyg. 1992, 47, 484–488. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holmes, C.J.; Benoit, J.B. Biological Adaptations Associated with Dehydration in Mosquitoes. Insects 2019, 10, 375. https://doi.org/10.3390/insects10110375
Holmes CJ, Benoit JB. Biological Adaptations Associated with Dehydration in Mosquitoes. Insects. 2019; 10(11):375. https://doi.org/10.3390/insects10110375
Chicago/Turabian StyleHolmes, Christopher J., and Joshua B. Benoit. 2019. "Biological Adaptations Associated with Dehydration in Mosquitoes" Insects 10, no. 11: 375. https://doi.org/10.3390/insects10110375
APA StyleHolmes, C. J., & Benoit, J. B. (2019). Biological Adaptations Associated with Dehydration in Mosquitoes. Insects, 10(11), 375. https://doi.org/10.3390/insects10110375