Gall-Colonizing Ants and Their Role as Plant Defenders: From ’Bad Job’ to ’Useful Service’

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Gall Selection
2.2. Gall Colonisation
2.2.1. Field Survey
2.2.2. Experiment 1
2.3. Nest Architecture
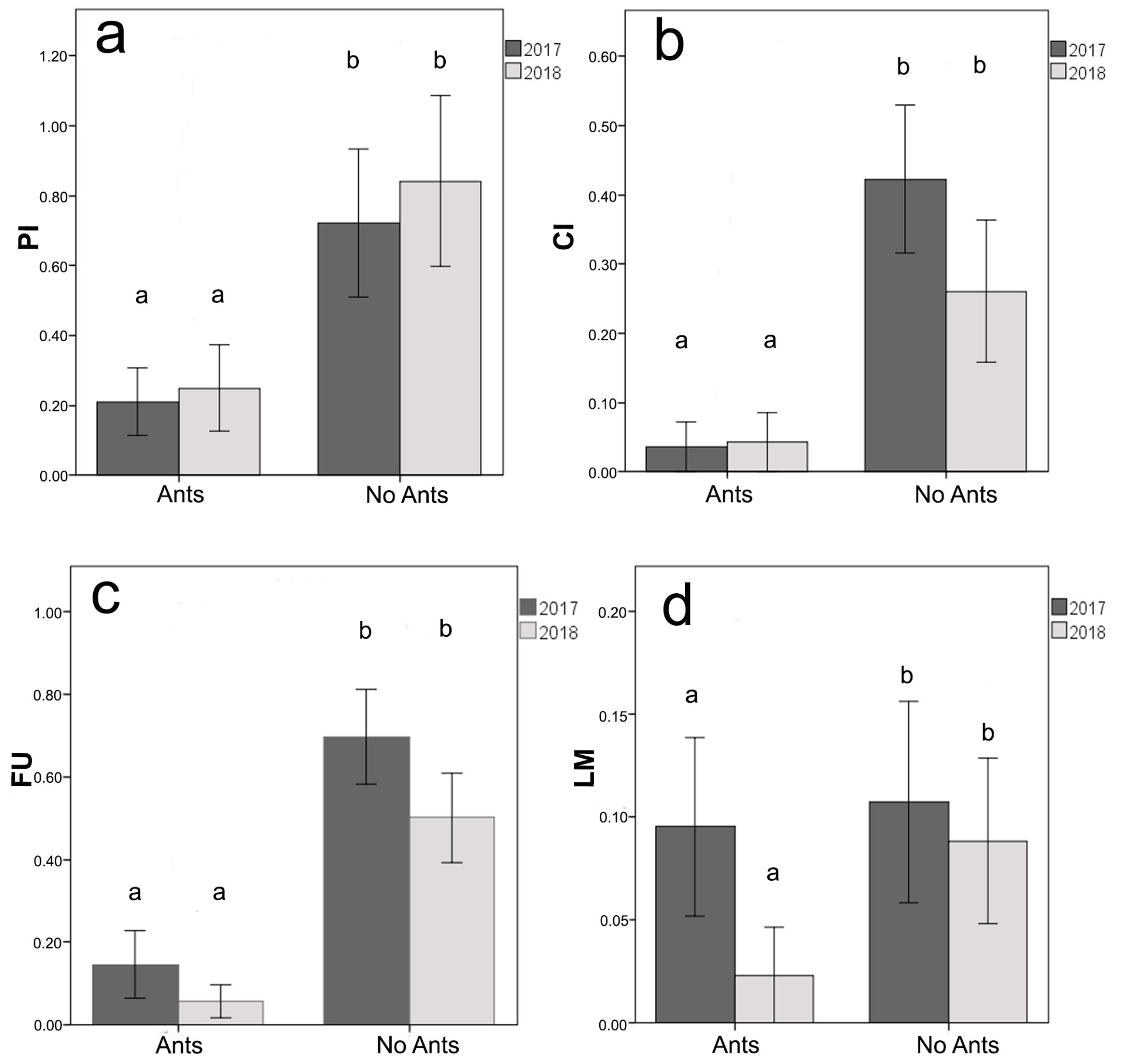
2.4. Defensive Effect of Ants on Plants: Experiment 2
- PI: Number of phytophagous insects counted in all the 24 samplings (only caterpillars of Lepidoptera and adult of Coleoptera were present)
- CI: the proportion of leaves attacked by chewing insects on the total leaves in each plant at the end of the experiment.
- FU: the proportion of leaves attacked by fungi on the total leaves in each plant at the end of the experiment.
- LM: the proportion of leaves attacked by leaf miners on the total leaves in each plant at the end of the experiment.
3. Results
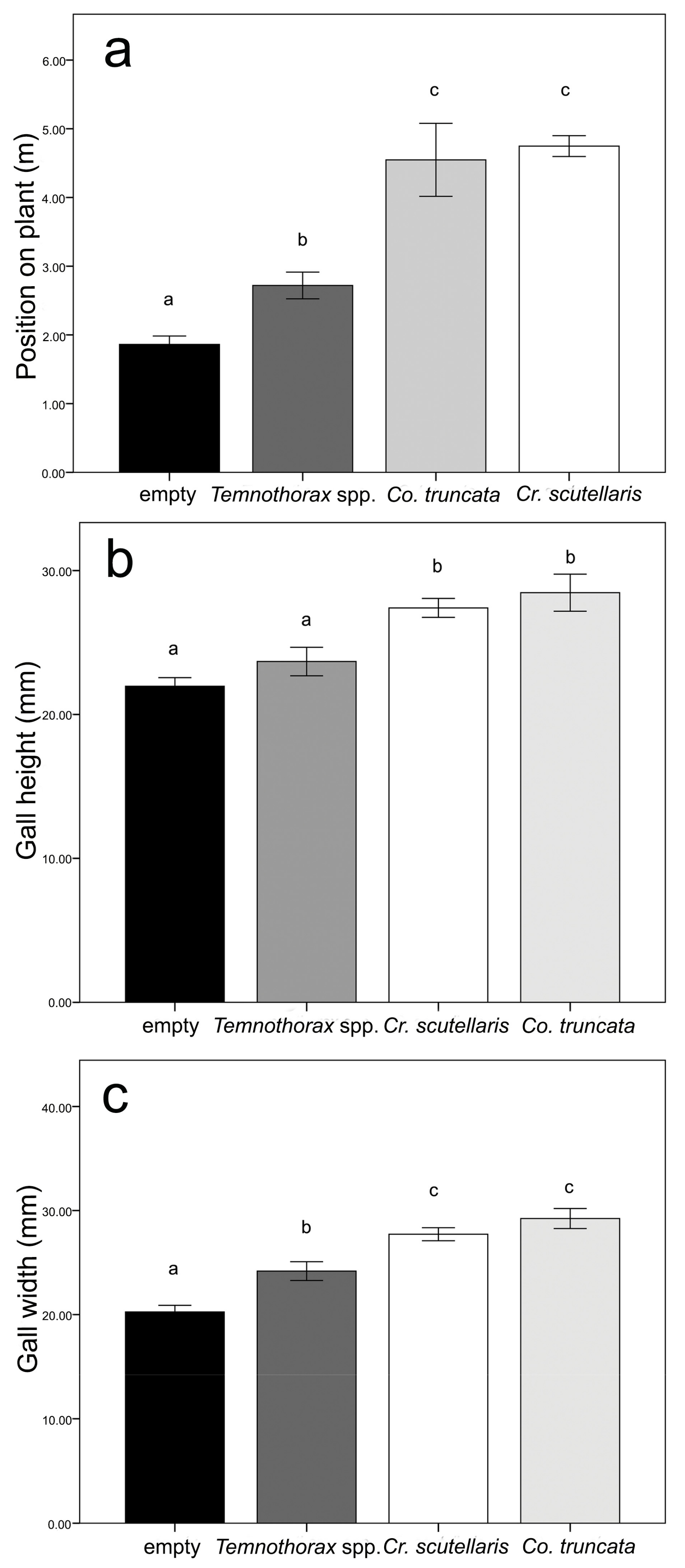
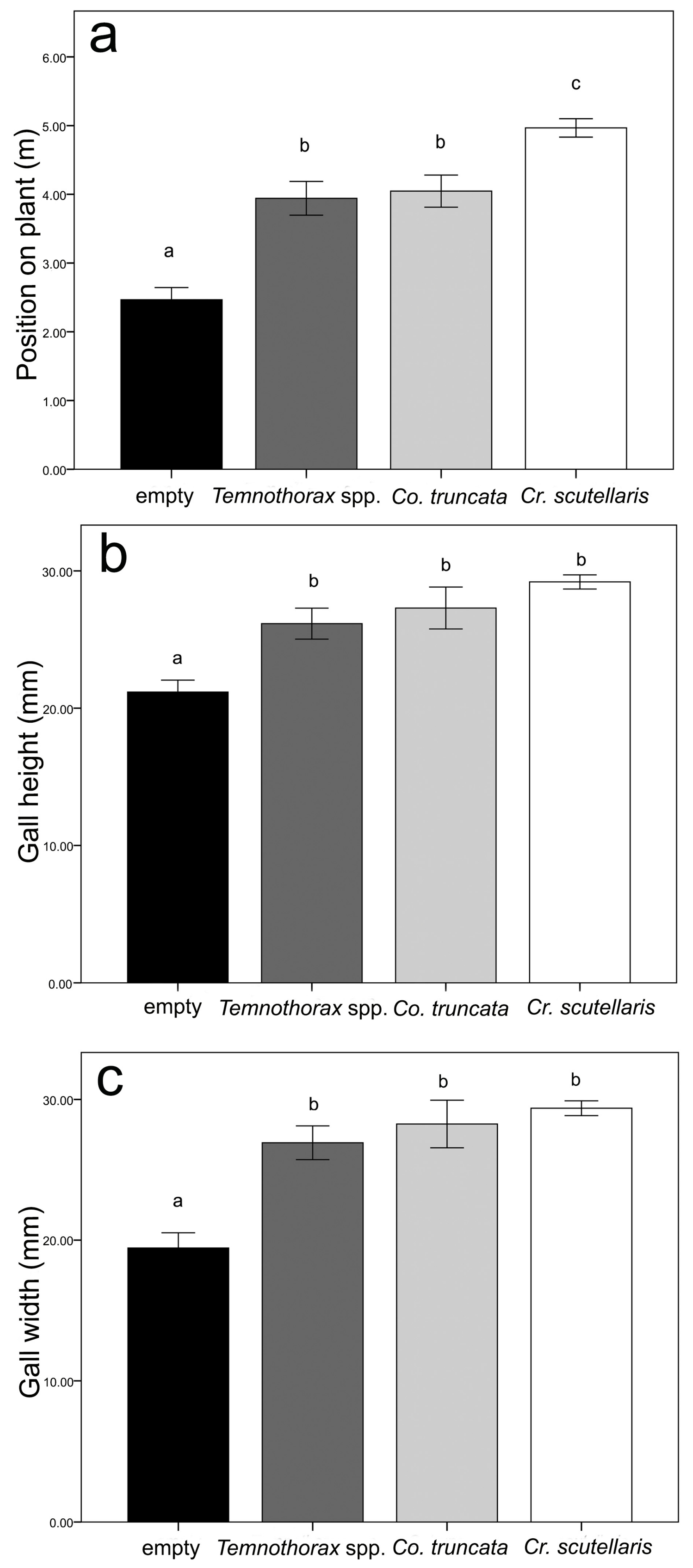
3.1. Gall Colonisation: Field Survey and Experiment 1
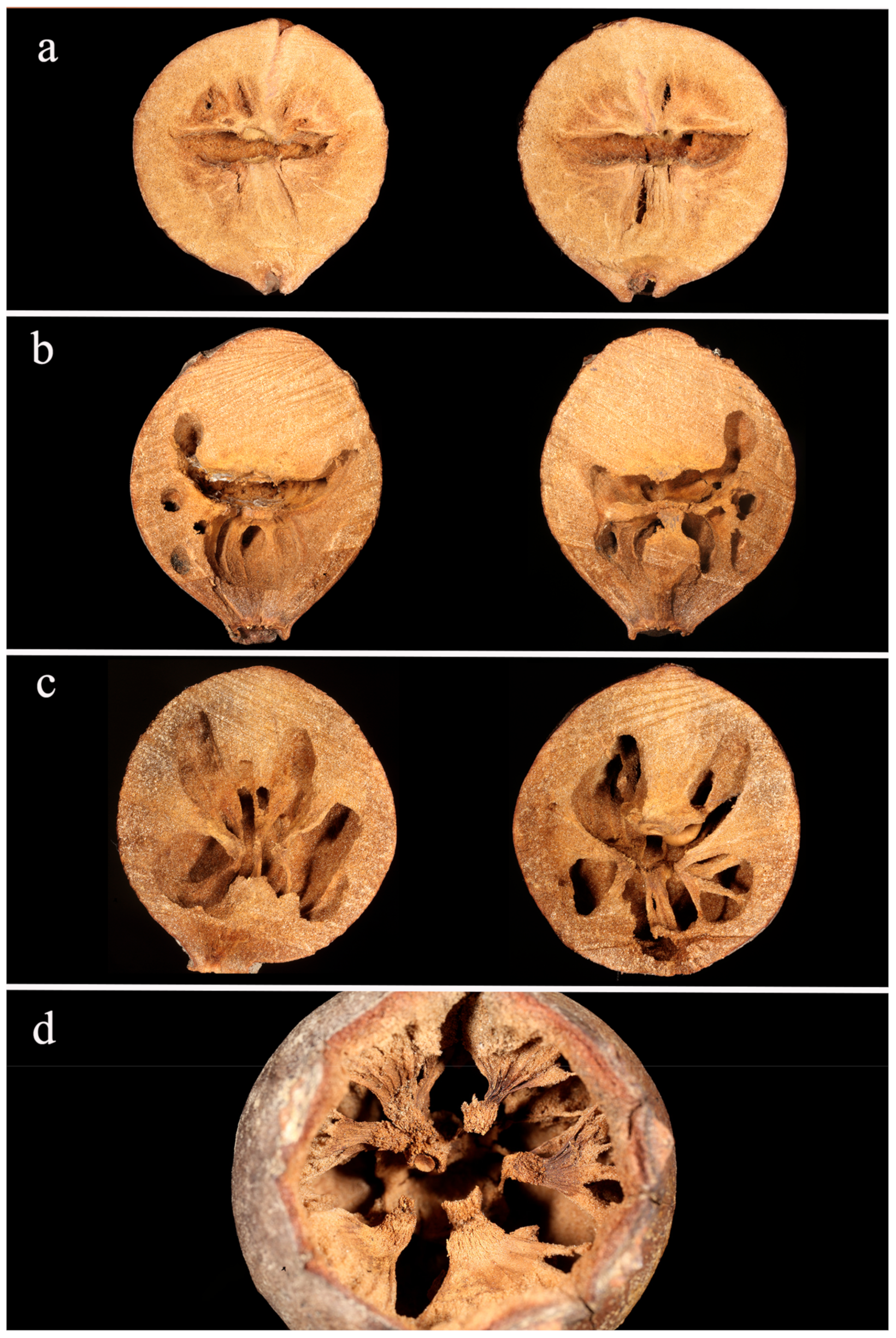
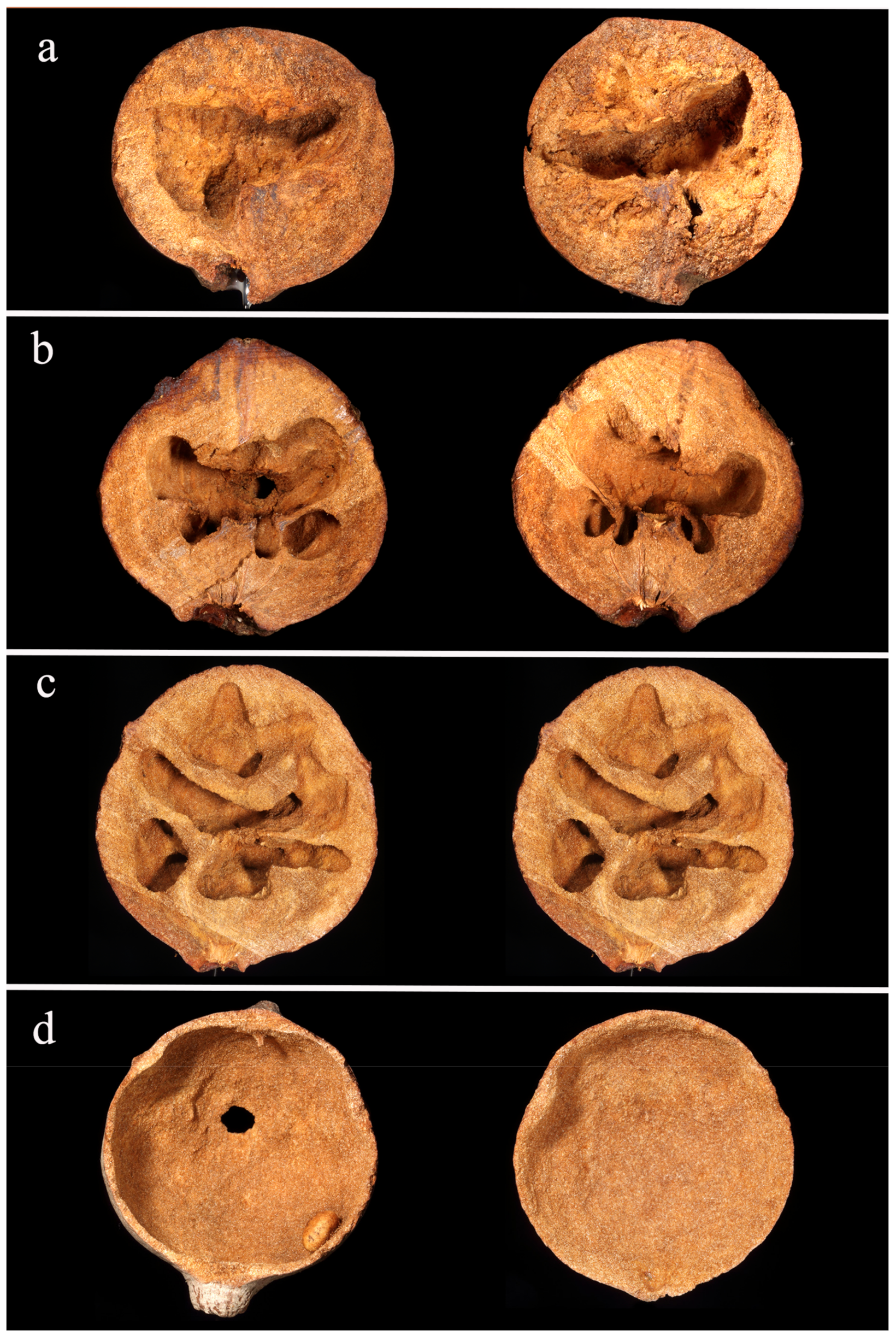
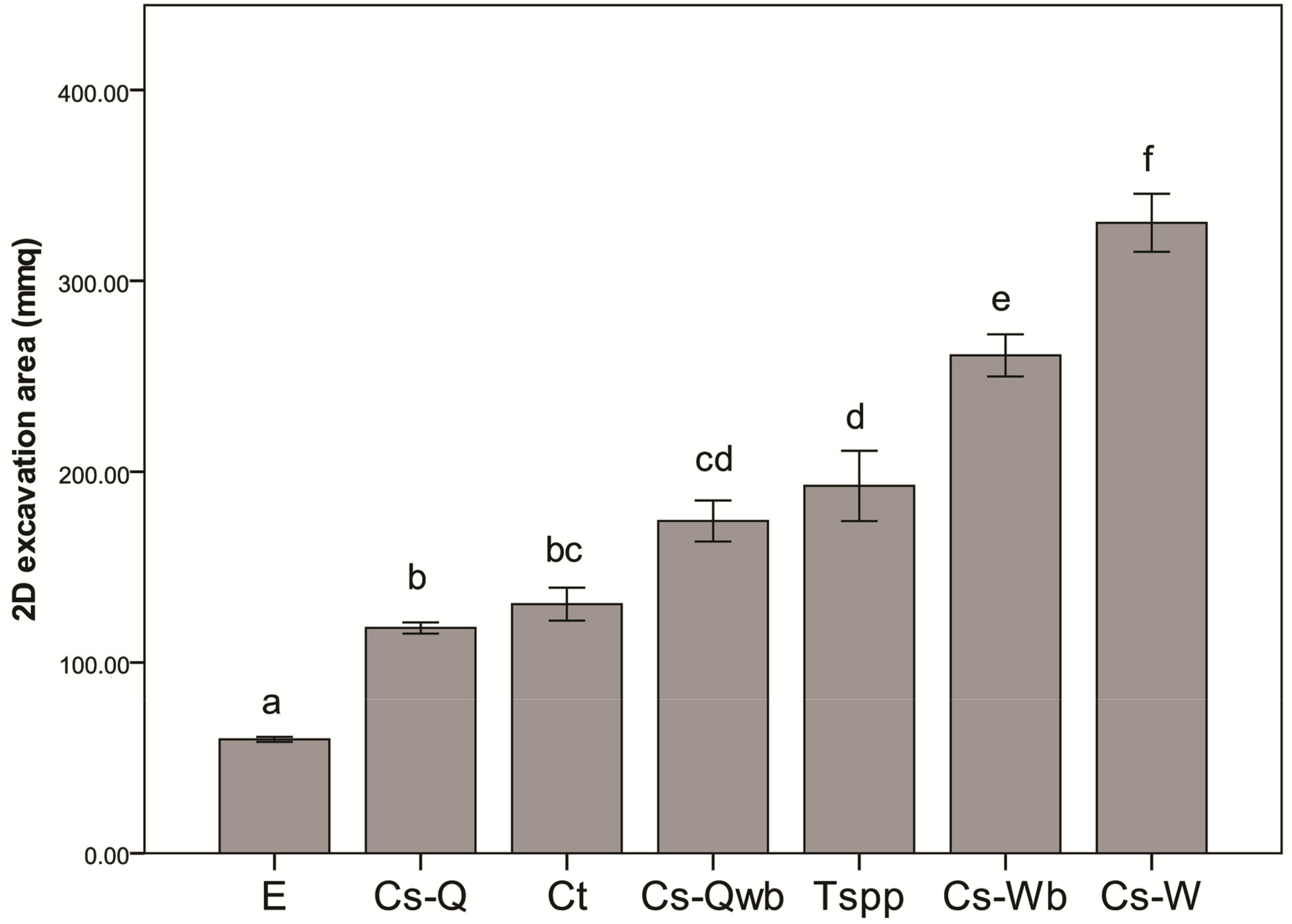
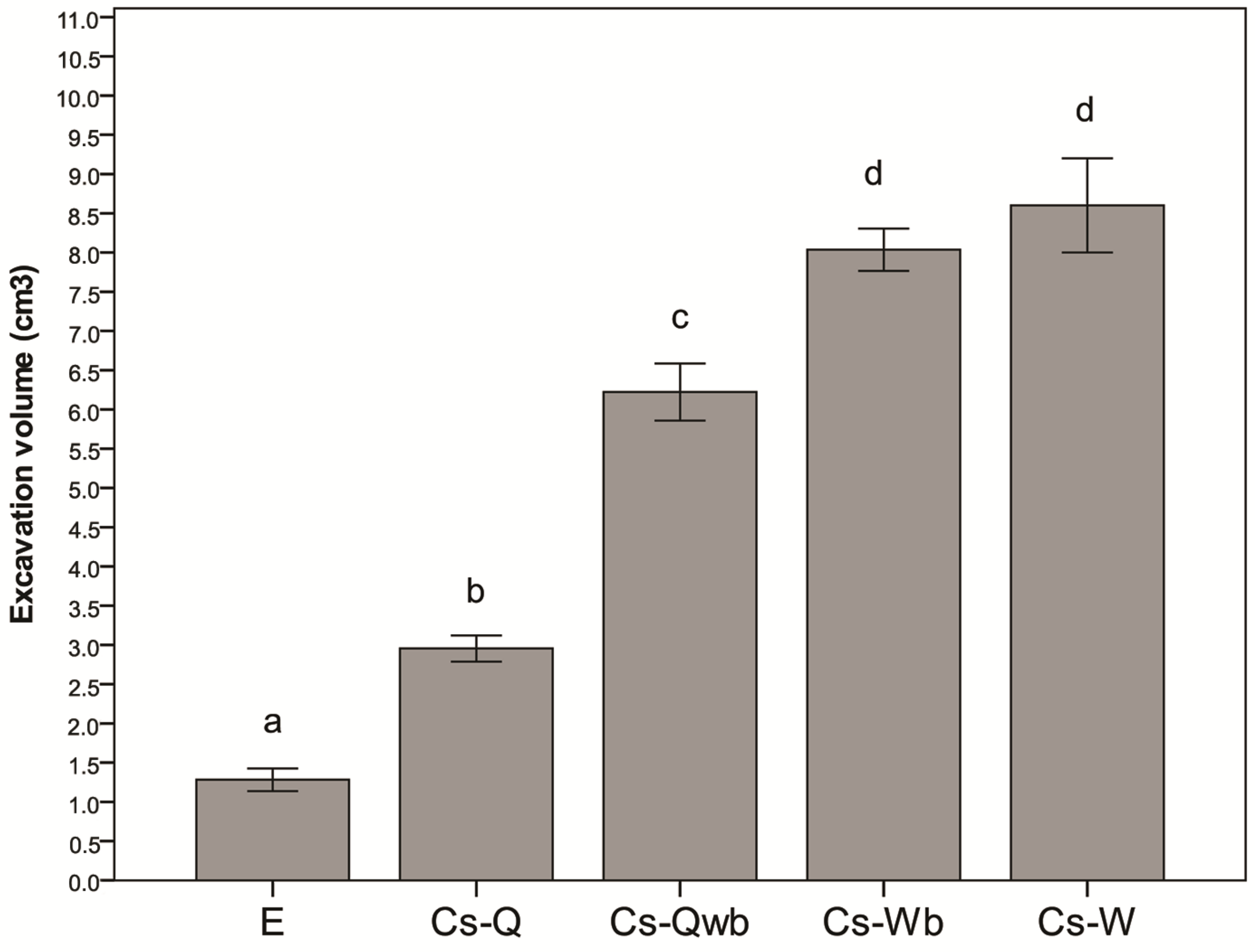
3.2. Nest Architecture
3.3. Defensive Effect of Ants on Plants: Experiment 2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Redfern, M.; Shirley, P.; Bloxham, M. British Plant Galls: Identification of Galls on Plants and Fungi, 1st ed.; FSC Publications: Shrewsbury, UK, 2002; p. 325. [Google Scholar]
- Price, W.P. Adaptive radiation of gall-inducing insects. Basic Appl. Ecol. 2005, 6, 413–421. [Google Scholar] [CrossRef]
- Shorthouse, J.D.; Wool, D.; Raman, A. Gall-inducing insects—Nature’s most sophisticated herbivores. Basic Appl. Ecol. 2005, 6, 407–411. [Google Scholar] [CrossRef]
- Bomfim, P.M.; Cardoso, J.C.; Rezende, U.C.; Martini, V.C.; Oliveira, D.C. Red galls: The different stories of two gall types on the same host. Plant Biol. 2019, 21, 284–291. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Santos, J.C. Neotropical Insect Galls; Springer: Dordrecht, The Netherlands, 2014; Volume 550. [Google Scholar]
- Ronquist, F.; Nieves-Aldrey, J.L.; Buffington, M.L.; Liu, Z.; Liljeblad, J.; Nylander, J.A.A. Phylogeny, evolution and classification of gall wasps: The plot thickens. PLoS ONE 2015, 10, e0123301. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K.; Atkinson, R.A.; Bellido, D.; Pujade-Villar, J. The population biology of oak gall wasps (Hymenoptrera: Cynipidae). Annu. Rev. Entomol. 2002, 47, 633–668. [Google Scholar] [CrossRef]
- Mani, M.S. The Ecology of Plant Galls, 1st ed.; Dr. W. Junk: The Hague, The Netherlands, 1964; p. 334. [Google Scholar]
- Espírito-Santo, M.M.; Fernandes, G.W. How many species of gall-inducing insects are there on Earth, and where are they? Ann. Entomol. Soc. Am. 2007, 100, 95–99. [Google Scholar]
- Ronquist, F.; Liljeblad, J. Evolution of the gall wasp host plant association. Evolution 2001, 55, 2503–2522. [Google Scholar] [PubMed]
- Mutun, S.; Dinç, S. Contributions to the Gallwasp (Hymenoptera: Cynipidae) Fauna of Turkey with one new record. J. Appl. Sci. 2011, 5, 83–85. [Google Scholar]
- Sanver, D.; Hawkins, B.A. Galls as habitats: The inquiline communities of insect galls. Basic Appl. Ecol. 2000, 1, 3–11. [Google Scholar] [CrossRef]
- Crawford, K.M.; Crutsinger, G.M.; Sanders, N.J. Host-plant genotypic diversity mediates the distribution of an ecosystem engineer. Ecology 2007, 88, 2114–2120. [Google Scholar] [CrossRef]
- Maruyama, P.K.; Nahas, L.; Moura-Neto, C.; Santos, J.C. Gall-inducing nematodes as ecosystem engineers for arthropods associated with its host plant in the Cerrado of Brazil. Stud. Neotrop. Fauna. Environ. 2012, 47, 131–138. [Google Scholar] [CrossRef]
- Wetzel, W.C.; Screen, R.M.; Li, I.; McKenzie, J.; Phillips, K.A.; Cruz, M.; Zhang, W.; Greene, A.; Lee, E.; Singh, N.; et al. Ecosystem engineering by a gall-forming wasp indirectly suppresses diversity and density of herbivores on oak trees. Ecology 2016, 97, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Tòrossian, P.C. Faune secondaire des galles de Cynipidae: I. Étude systématique des fourmis et des principaux arthropodes récoltes dans les galles. Insectes Sociaux 1971, 18, 135–154. [Google Scholar] [CrossRef]
- Craig, T.P.; Araújo, L.M.; Itami, J.K.; Fernandes, G.W. Development of the insect community centered on a leaf-bud gall formed by a weevil (Coleoptera: Curculionidae) on Xylopia aromatica (Annonaceae). Rev. Bras. Entomol. 1991, 35, 311–317. [Google Scholar]
- Wheeler, J.; Longino, J.T. Arthropods in live oak galls in Texas. Entomol. News 1988, 99, 25–29. [Google Scholar]
- Cornelissen, T.; Cintra, F.; Santos, J.C. Shelter-building insects and their role as ecosystem engineers. Neotrop. Entomol. 2016, 45, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tòrossian, P.C. Étude biologique des fourmis forestières peuplant les galles de Cynipidae des chênes. Insectes Sociaux 1971, 18, 193–202. [Google Scholar] [CrossRef]
- Tòrossian, P.C. Étude biologique des fourmis forestières peuplant les galles de Cynipidae des chênes: III. Rôle et importance numérique des femelles fondatrices. Insectes Sociaux 1972, 19, 25–38. [Google Scholar] [CrossRef]
- Espadaler, X.; Nieves, J.L. Hormigas (Hymenoptera; Formicidae) pobladoras de las gallas abandonadas de Cinipedos (Hymenoptera Cynipidae) sobre “Quercus sp.” en la penisola Iberica. Bol. Estac. Cent. Ecol. 1983, 12, 89–93. [Google Scholar]
- Santos, L.R.; Feitosa, R.M.; Carneiro, M.A.A. The role of senescent stem-galls over arboreal ant communities structure in Eremanthus erythropappus (DC.) MacLeish (Asteraceae) Trees. Sociobiology 2017, 64, 7–13. [Google Scholar] [CrossRef]
- Heil, M.; McKey, D. Protective ant-plant interactions as model systems in ecological and evolutionary research. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 425–453. [Google Scholar] [CrossRef]
- Rosumek, F.B.; Silveira, F.A.O.; Neves, F.S.; Barbosa, N.P.; Diniz, I.; Oki, Y.; Pezzini, F.; Fernandes, G.W.; Cornelissen, T. Ants on plants: A meta-analysis of the role of ants as plant biotic defenses. Oecologia 2009, 160, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Nepi, M.; Grasso, D.A.; Mancuso, S. Nectar in plant-insect mutualistic relationships: From food reward to partner manipulation. Front. Plant Sci. 2018, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Rico-Grey, V.; Oliveira, P.S. The Ecology and Evolution of Ant-Plant Interactions, 1st ed.; The University of Chicago Press: Chicago, IL, USA, 2007; p. 320. [Google Scholar]
- Grasso, D.A.; Pandolfi, C.; Bazihizina, N.; Nocentini, D.; Nepi, M.; Mancuso, S. Extrafloral-nectar-based partner manipulation in plant-ant relationships. AoB Plants 2015, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Heil, M. Extrafloral nectar at the plant-insect interface: A spotlight on chemical ecology, phenotypic plasticity, and food webs. Annu. Rev. Entomol. 2015, 60, 213–232. [Google Scholar] [CrossRef]
- Del-Claro, K.; Rico-Gray, V.; Torezan-Silingardi, H.M.; Alves-Silva, E.; Fagundes, R.; Lange, D.; Lange, D.; Dáttilo, W.; Vilela, A.A.; Aguirre, A.; et al. Loss and gains in ant–plant interactions mediated by extrafloral nectar: Fidelity, cheats, and lies. Insectes Sociaux 2016, 63, 207–221. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Barbosa, P. Ants, stem borers, and pubescence in Endospermum in Papua New Guinea. Biotropica 1999, 31, 295–302. [Google Scholar] [CrossRef]
- Crutsinger, G.M.; Sanders, N.J. Aphid-tending ants affect secondary users in leaf shelters and rates of herbivory on Salix hookeriana in a coastal dune habitat. Am. Midl. Nat. 2005, 154, 296–304. [Google Scholar] [CrossRef]
- Del-Claro, K.; Byke, J.; Yugue, G.M.; Morato, M.G. Conservative benefits in an ant-hemipteran association in the Brazilian tropical savanna. Sociobiology 2006, 47, 415–421. [Google Scholar]
- Fagundes, R.; Dáttilo, W.; Riberio, S.P.; Rico-Gray, V.; Jordano, P.; Del-Claro, K. Differences among ant species in plant protection are related to production of extrafloral nectar and degree of leaf herbivory. Biol. J. Linn. Soc. 2017, 122, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Sendoya, S.F.; Blüthgen, N.; Tamashiro, J.Y.; Fernandez, F.; Oliveira, P.S. Foliage-dwelling ants in a neotropical savanna: Effects of plant and insect exudates on ant communities. Arthropod-Plant Interact. 2016, 10, 183–195. [Google Scholar] [CrossRef]
- Letourneau, D.K. Ants, stem-borers, and fungal pathogens: Experimental tests of a fitness advantage in Piper ant–plants. Ecology 1998, 79, 593–603. [Google Scholar] [CrossRef]
- Freitas, L.; Galetto, L.; Bernardello, G.; Paoli, A.A.S. Ant exclusion and reproduction of Croton sarcopetalus (Euphorbiaceae). Flora 2000, 195, 398–402. [Google Scholar] [CrossRef]
- Chamberlain, S.C.; Holland, J.N. Quantitative synthesis of contex dependency in ant-plant protection mutualisms. Ecology 2009, 90, 2385–2392. [Google Scholar] [CrossRef]
- De Stefani, T. Cinipidi e loro galle. In Atti della Reale Accademia di Scienze, Lettere e Belle Arti di Palermo; Tipografia E Ferrigno e F Ando: Palermo, Italia, 1888; Volume 10, p. 12. [Google Scholar]
- De Stefani, T. Miscellanea imenotterologica sicula. Nat. Sicil. 1889, 8, 203–208. [Google Scholar]
- Graziosi, I.; Santi, F. Chestnut gall wasp (Dryocosmus kuriphilus): Spreading in Italy and new records in Bologna province. Bull. Insectol. 2008, 61, 343–348. [Google Scholar]
- Cerasa, G.; Verde, G.L.; Caleca, V.; Massa, B.; Nicholls, J.A.; Melika, G. Description of Dryocosmus destefanii new species (Hymenoptera: Cynipidae: Cynipini) from Quercus suber L. in Italy. Zootaxa 2018, 4370, 535–548. [Google Scholar] [CrossRef]
- Panzavolta, T.; Croci, F.; Bracalini, M.; Melika, G.; Benedettelli, S.; Tellini Florenzano, G.; Tiberi, R. Population dynamics of native parasitoids associated with the Asian Chestnut Gall Wasp (Dryocosmus kuriphilus) in Italy. Psyche 2018, 2018, 8078049. [Google Scholar] [CrossRef]
- Rokas, A.; Atkinson, R.J.; Webster, L.M.I.; Csόka, G.; Stone, G.N. Out of Anatolia: Longitudinal gradients in genetic diversity support an eastern origin for a circum-Mediterranean oak gallwasp Andricus quercustozae. Mol. Ecol. 2003, 12, 2153–2174. [Google Scholar] [CrossRef]
- Cook, J.M.; Rokas, A.; Pagel, M.; Stone, G.N. Evolutionary shifts between host oak sections and host-plant organs in Andricus gallwasps. Evolution 2002, 56, 1821–1830. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants, 1st ed.; Harvard University Press: Cambridge, MA, USA, 1990; p. 746. [Google Scholar]
- Seifert, B. The Ants of Central and North Europe, 1st ed.; Lutra Verlags Vertriebsgesellschaft: Tauer, Germany, 2018; p. 408. [Google Scholar]
- Consani, M.; Zangheri, P. Fauna di Romagna. Imenotteri—Formicidi. Memorie della Società Entomologica Italiana 1952, 31, 38–48. [Google Scholar]
- Baroni Urbani, C. Catalogo delle specie di Formicidae d’Italia. Memorie della Società Entomologica Italiana 1971, 50, 5–287. [Google Scholar]
- Krausse, A.H. Ueber sardische Ameisen. Archiv für Naturgeschichte 1912, 78, 162–166. [Google Scholar]
- Monastero, S. Notizie sulla formica argentina e altre dannose formiche di Sicilia. Metodi di difesa e di lotta. Atti. R. Acc. Lett. Arti Palermo 1942, 3, 3–40. [Google Scholar]
- Almeida, M.F.B.; Santos, L.R.; Carneiro, M.A.A. Senescent stem-galls in trees of Eremanthus erythropappus as a resource for arboreal ants. Rev. Bras. Entomol. 2014, 58, 265–272. [Google Scholar] [CrossRef]
- Bernard, F. Les Fourmis (Hymenoptera Formicidae) d’Europe occidentale et septentrionale, 1st ed.; Masson et Cie: Paris, France, 1968; p. 411. [Google Scholar]
- Lebas, C.; Galkowski, C.; Blatrix, R.; Wegnez, P. Fourmis d’Europe occidentale, 1st ed.; Delachaux et Niestlè: Paris, France, 2016; p. 415. [Google Scholar]
- Casevitz Weulersse, J. Sur la Biologie de Crematogaster scutellaris Olivier (Hymenoptera, Formicidae, Myrmicinae). L’Entomologiste 1970, 36, 68–75. [Google Scholar]
- Cammell, M.E.; Way, M.J.; Paiva, M.R. Diversity and structure of ant communities associated with oak, pine, eucalyptus and arable habitats in Portugal. Insectes Sociaux 1996, 43, 37–46. [Google Scholar] [CrossRef]
- Way, M.J.; Cammell, M.E.; Paiva, M.R.; Collingwood, C.A. Distribution and dynamics of the Argentine ant Linepithema (Iridomyrmex) humile (Mayr) in relation to vegetation, soil conditions, topography and native competitor ants in Portugal. Insectes Sociaux 1997, 44, 415–433. [Google Scholar] [CrossRef]
- Santini, G.; Tucci, L.; Ottonetti, L.; Frizzi, F. Competition trade-offs in the organisation of a Mediterranean ant assemblage. Ecol. Entomol. 2007, 32, 319–326. [Google Scholar] [CrossRef]
- Santini, G.; Ramsay, M.P.; Tucci, L.; Ottonetti, L.; Frizzi, F. Spatial patterns of the ant Crematogaster scutellaris in a model ecosystem. Ecol. Entomol. 2011, 36, 625–634. [Google Scholar] [CrossRef]
- Solida, L.; Scalisi, M.; Fanfani, A.; Mori, A.; Grasso, D.A. Interspecific space partitioning during the foraging activity of two syntopic species of Messor harvester ants. J. Biol. Res. Thessalon. 2010, 13, 3–12. [Google Scholar]
- Casevitz Weulersse, J. Habitats et comportement nidificateur de Crematogaster scutellaris Olivier. Bull. Soc. Entomol. 1972, 77, 12–19. [Google Scholar]
- Debout, G.; Schatz, B.; Elias, M.; Mckey, D. Polydomy in ants: What we know, what we think we know, and what remains to be done. Biol. J. Linn. Soc. 2007, 9, 319–348. [Google Scholar] [CrossRef]
- Frizzi, F.; Ciofi, C.; Dapporto, L.; Natali, C.; Chelazzi, G.; Turillazzi, S.; Santini, G. The rules of aggression: How genetic, chemical and spatial factors affect intercolony fights in a dominant species, the Mediterranean acrobat ant Crematogaster scutellaris. PLoS ONE 2015, 10, e0137919. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; McShea, D.W. Intermediate-level parts in insect societies: Adaptive structures that ants build away from the nest. Insectes Sociaux 2001, 48, 291–301. [Google Scholar] [CrossRef]
- Lanan, M.C.; Dornhaus, A.; Bronstein, J.L. The function of polydomy: The ant Crematogaster torosa preferentially forms new nests near food sources and fortifies outstations. Behav. Ecol. Sociobiol. 2011, 65, 959–968. [Google Scholar] [CrossRef]
- Tschinkel, W.R. The nest architecture of the Florida harvester ant, Pogonomyrmex badius. J. Insect Sci. 2004, 4, 21. [Google Scholar] [CrossRef]
- Tschinkel, W.R.; Hanley, N. Vertical organization of the division of labor within nests of the Florida harvester ant, Pogonomyrmex badius. PLoS ONE 2017, 12, e0188630. [Google Scholar] [CrossRef]
- Varoudis, T.; Swenson, A.G.; Kirkton, S.D.; Waters, J.S. Exploring nest structures of acorn dwelling ants with X-ray microtomography and surface-based three-dimensional visibility graph analysis. Philos. Trans. R. Soc. B 2018, 373, 20170237. [Google Scholar] [CrossRef]
- Karlik, J.; Jane Epps, M.; Dunn, R.R.; Penic, C.A. Life Inside an Acorn: How Microclimate and Microbes Influence Nest Organization in Temnothorax Ants. Ethology 2016, 122, 790–797. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Rutter, M.T. Dynamic anti-herbivore defense in ant-plants: The role of induced responses. Oikos 1998, 83, 227–236. [Google Scholar] [CrossRef]
- Bronstein, J.L. The contribution of ant–plant protection studies to our understanding of mutualism. Biotropica 1998, 30, 150–161. [Google Scholar] [CrossRef]
- Heil, M. Indirect defence via tritrophic interactions. New Phytol. 2008, 178, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Campolo, O.; Palmeri, V.; Malacrinò, A.; Laudani, F.; Castracani, C.; Mori, A.; Grasso, D.A. Interaction between ants and the Mediterranean fruit fly: New insights for biological control. Biol. Control. 2015, 90, 120–127. [Google Scholar] [CrossRef]
- Castracani, C.; Bulgarini, G.; Giannetti, D.; Spotti, F.A.; Maistrello, L.; Mori, A.; Grasso, D.A. Predatory ability of the ant Crematogaster scutellaris on the brown marmorated stink bug Halyomorpha halys. J. Pest Sci. 2017, 90, 1181–1190. [Google Scholar] [CrossRef]
- Letourneau, D.K. Passive aggression: An alternative hypothesis for the Piper-Pheidole association. Oecologia 1983, 60, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Gonzàlez-Teuber, M.; Kaltenpoth, M.; Boland, W. Mutualistic ants as an indirect defence against leaf pathogens. New Phytol. 2014, 202, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Traniello, J.F.A.; Rosengaus, R.B.; Savoie, K. The development of immunity in a social insect: Evidence for the group facilitation of disease resistance. Proc. Natl. Acad. Sci. USA 2002, 99, 6838–6842. [Google Scholar] [CrossRef] [Green Version]
- Solida, L.; Grasso, D.A.; Testi, A.; Fanelli, G.; Scalisi, M.; Bartolino, V.; Mori, A.; Fanfani, A. Differences in the nesting sites microhabitat characteristics of two syntopic species of Messor harvester ants in a phytosociological homogeneous grassland area. Ethol. Ecol. Evol. 2011, 23, 229–239. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. Trends Ecol. Evol. 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Heil, M.; Koch, T.; Hilpert, A.; Fiala, B.; Boland, W.; Linsenmair, K.E. Extrafloral nectar production of the ant-associated plant, Macaranga tanarius, is an induced, indirect, defensive response elicited by jasmonic acid. Proc. Natl. Acad. Sci. USA 2001, 98, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Pierce, M.P. The ecological and evolutionary importance of nectar-secreting galls. Ecosphere 2019, 10, e02670. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Field Survey | Field Experiment 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tot | Colony Composition | Tot | Colony Composition | |||||||
| Q | QWB | WB | W | Q | QWB | WB | W | |||
| Crematogaster scutellaris (Olivier, 1792) | 39 | 3 | 14 | 19 | 3 | 69 | 23 | 9 | 28 | 9 |
| Colobopsis truncata (Spinola, 1808) | 9 | - | 1 | - | 8 | 12 | 3 | - | - | 9 |
| Temnothorax albipennis (Curtis, 1854) | 6 | - | 6 | - | - | 7 | - | 7 | - | - |
| Temnothorax unifasciatus (Latreille, 1798) | 2 | - | 2 | - | - | 1 | - | 1 | - | - |
| Temnothorax italicus (Consani,1952) | 10 | - | 10 | - | - | 2 | - | 2 | - | - |
| Dolichoderus quadripunctatus (Linneus, 1771) | 4 | - | - | - | 4 | 3 | - | - | - | 3 |
| Camponotus fallax (Nylander, 1856) | 1 | - | 1 | - | - | - | - | - | - | - |
| Camponotus lateralis (Olivier, 1792) | 1 | - | 1 | - | - | - | - | - | - | - |
| Crematogaster scutellaris nigra (Krausse, 1912) | 1 | - | - | - | 1 | - | - | - | - | - |
| Temnothorax nylanderi (Foerster, 1850) | 1 | - | 1 | - | - | - | - | - | - | - |
| Total | 74 | 94 | ||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannetti, D.; Castracani, C.; Spotti, F.A.; Mori, A.; Grasso, D.A. Gall-Colonizing Ants and Their Role as Plant Defenders: From ’Bad Job’ to ’Useful Service’. Insects 2019, 10, 392. https://doi.org/10.3390/insects10110392
Giannetti D, Castracani C, Spotti FA, Mori A, Grasso DA. Gall-Colonizing Ants and Their Role as Plant Defenders: From ’Bad Job’ to ’Useful Service’. Insects. 2019; 10(11):392. https://doi.org/10.3390/insects10110392
Chicago/Turabian StyleGiannetti, Daniele, Cristina Castracani, Fiorenza A. Spotti, Alessandra Mori, and Donato A. Grasso. 2019. "Gall-Colonizing Ants and Their Role as Plant Defenders: From ’Bad Job’ to ’Useful Service’" Insects 10, no. 11: 392. https://doi.org/10.3390/insects10110392
APA StyleGiannetti, D., Castracani, C., Spotti, F. A., Mori, A., & Grasso, D. A. (2019). Gall-Colonizing Ants and Their Role as Plant Defenders: From ’Bad Job’ to ’Useful Service’. Insects, 10(11), 392. https://doi.org/10.3390/insects10110392