Stag Beetle Elytra: Localized Shape Retention and Puncture/Wear Resistance



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Optical and Electron Microscopy
2.3. Mechanical Testing
2.4. Finite Element Simulation
2.5. Nanoindentation
3. Results
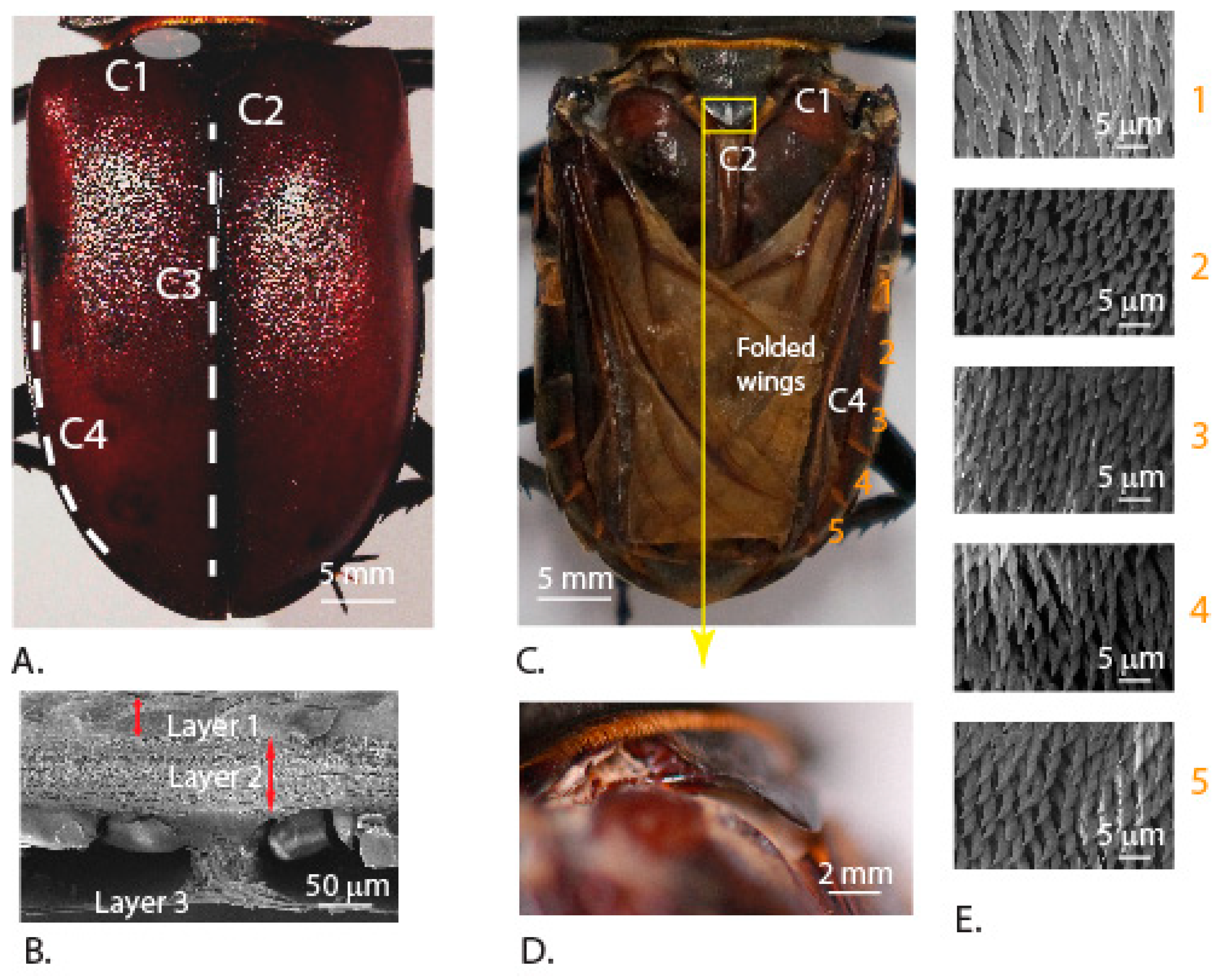
3.1. Microstructure of the Elytra and the Abdominal Surface
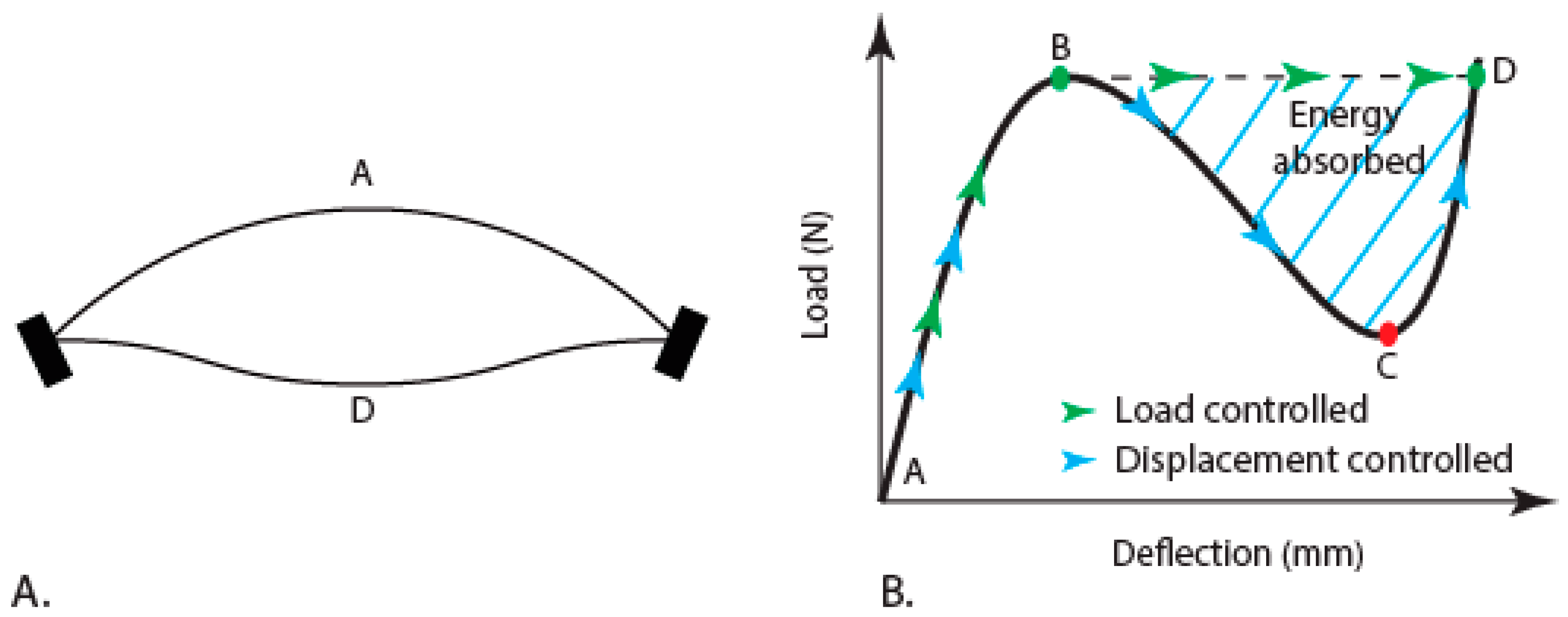
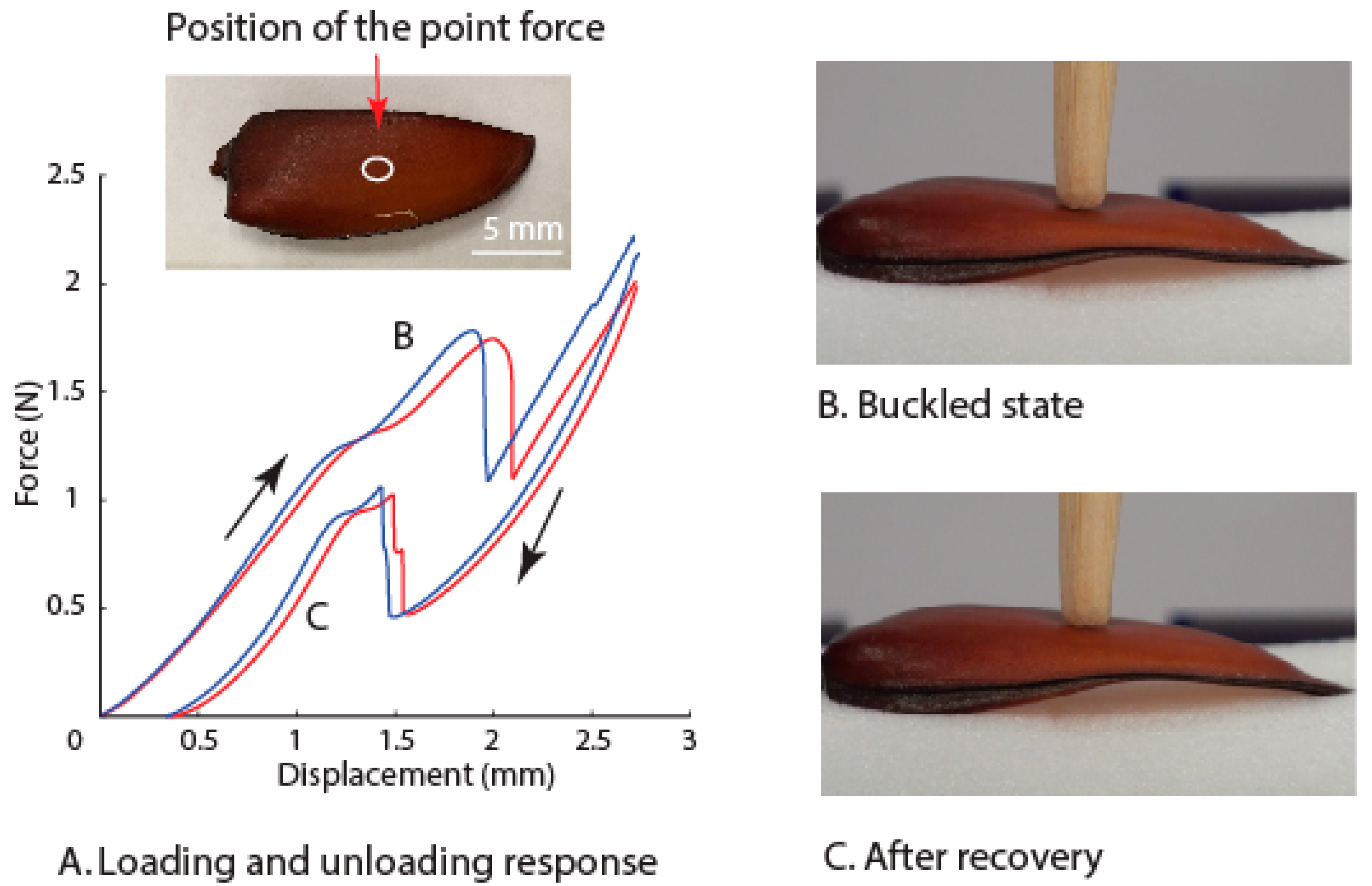
3.2. Deformation of the Elytra
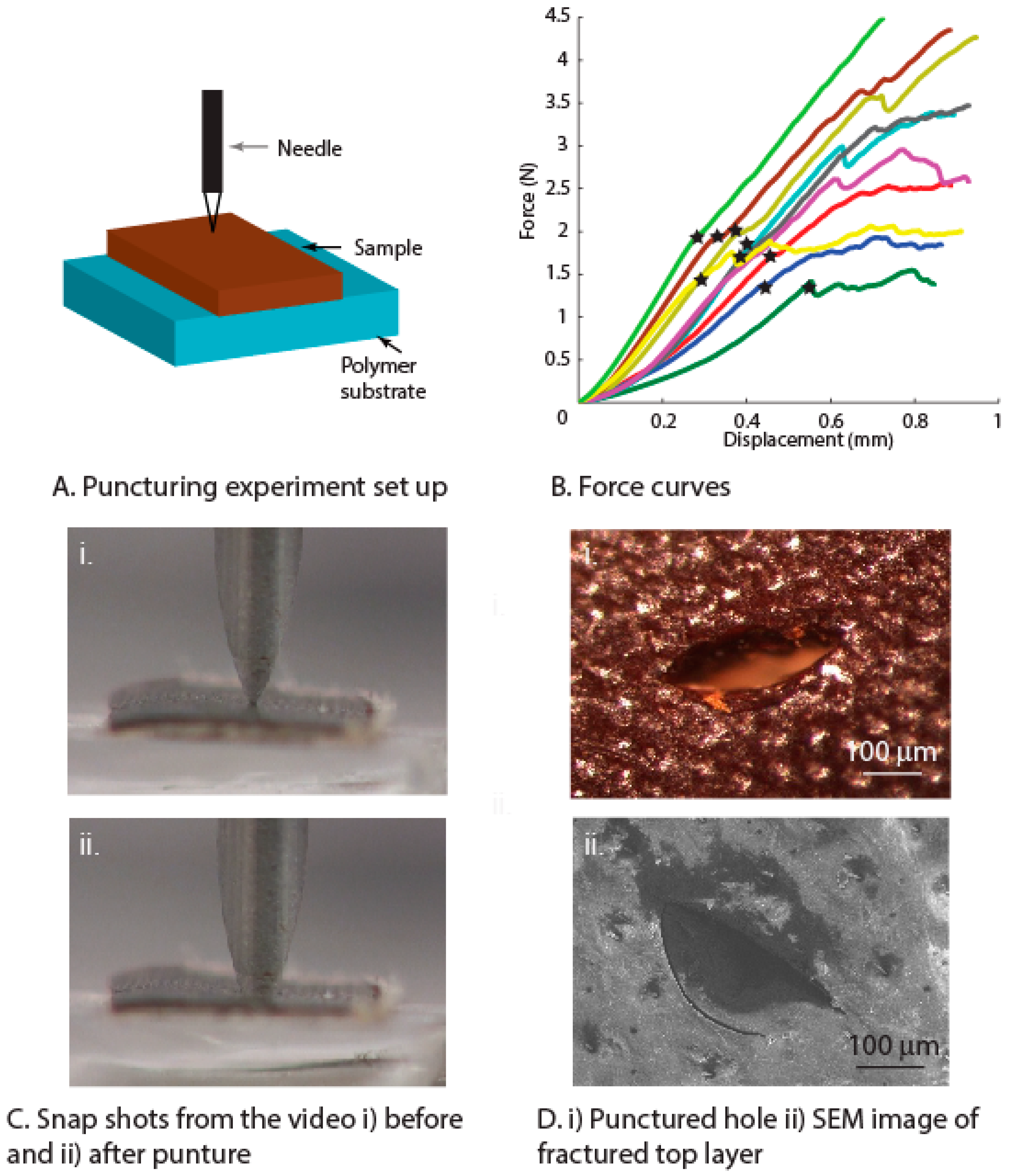
3.3. Puncture and Wear Resistance
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
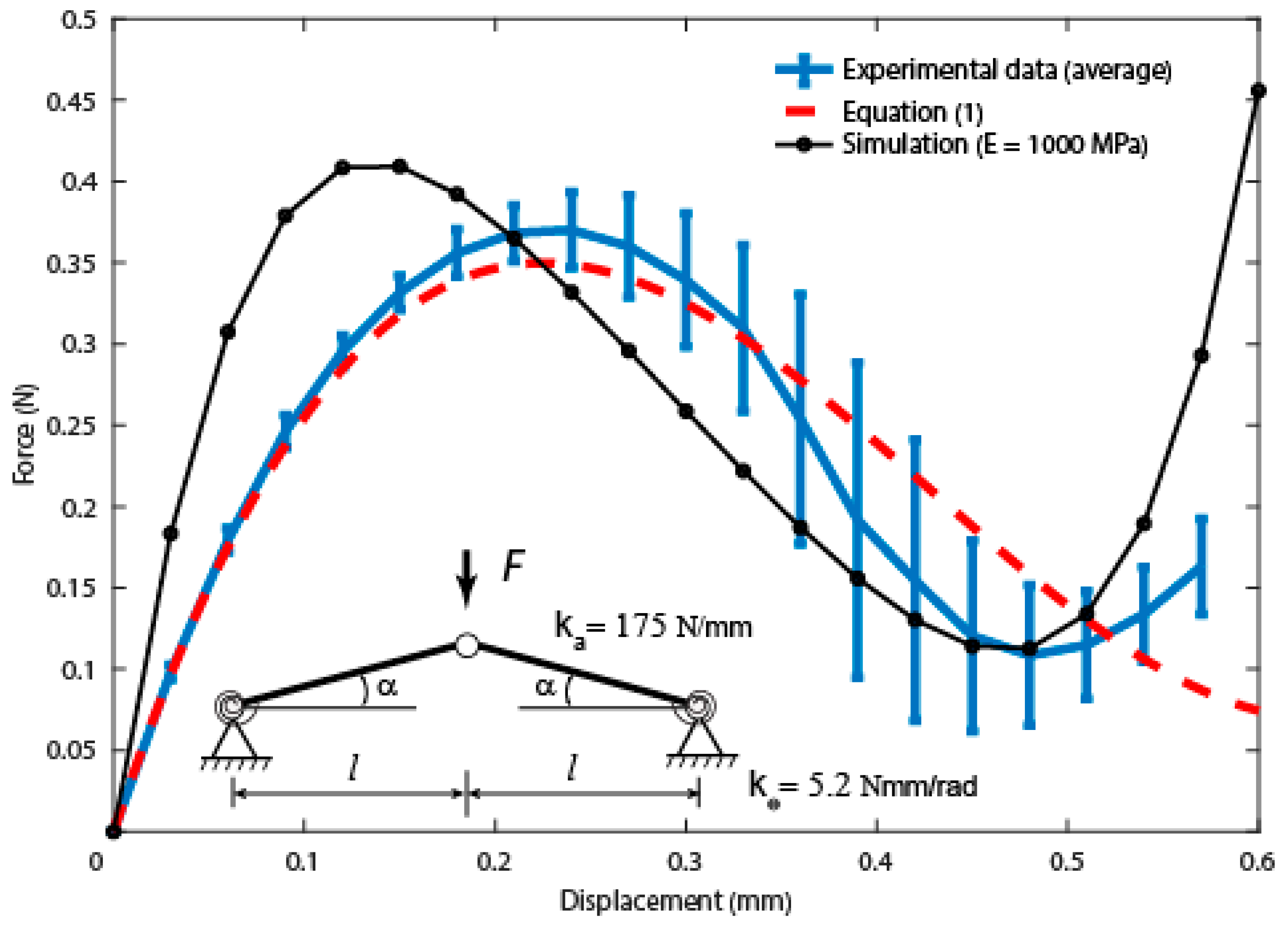
Analytical Estimation of the Snap-Through Curve

References
- Stork, N. Insect diversity: Facts, fiction and speculation. Biol. J. Linn. Soc. 1988, 35, 321–337. [Google Scholar] [CrossRef]
- Hunt, T. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.F.V.; Wegst, U.G.K. Design and mechanical properties of insect cuticle. Arthropod Struct. Dev. 2004, 33, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Van De Kamp, T.; Greven, H. On the architecture of beetle elytra. Entomol. Heute 2010, 22, 191–204. [Google Scholar]
- Lomakin, J.; Huber, P.A.; Eichler, C.; Arakane, Y.; Kramer, K.J.; Beeman, R.W.; Kanost, M.R.; Gehrke, S.H. Mechanical properties of the beetle elytron, a biological composite material. Biomacromolecules 2011, 12, 321–335. [Google Scholar] [CrossRef]
- Lomakin, J.; Arakane, Y.; Kramer, K.J.; Beeman, R.W.; Kanost, M.R.; Gehrke, S.H. Mechanical properties of elytra from Tribolium castaneum wild-type and body color mutant strains. J. Insect Physiol. 2010, 56, 1901–1906. [Google Scholar] [CrossRef]
- Conrad, M.B. Bioinspired Composites Design: Mechanical and Optical Characterization of the Hercules Beetle Elytra. Ph.D. Thesis, Naval Post Graduate School, Monterey, CA, USA, 2014. [Google Scholar]
- van de Kamp, T.; Doerstelmann, M.; dos Sanots Rolo, T.; Baumbach, T.; Menges, A.; Knippers, J. Beetle Elytra as Role Models for Lightweight Building Construction. Entomol. Heute 2015, 27, 149–158. [Google Scholar]
- Parle, E.; Herbaj, S.; Sheils, F.; Larmon, H.; Taylor, D. Buckling failures in insect exoskeletons. Bioinspir. Biomim. 2015, 11, 016003. [Google Scholar] [CrossRef] [Green Version]
- Kundanati, L.; Gundiah, N. Biomechanics of substrate boring by fig wasps. J. Exp. Biol. 2014, 217, 1946–1954. [Google Scholar] [CrossRef] [Green Version]
- Pini, V.; Ruz, J.J.; Kosaka, P.M.; Malvar, O.; Calleja, M.; Tamayo, J. How two-dimensional bending can extraordinarily stiffen thin sheets. Sci. Rep. 2016, 6, 29627. [Google Scholar] [CrossRef] [Green Version]
- Forterre, Y.; Skotheim, J.M.; Dumais, J.; Mahadevan, L. How the Venus flytrap snaps. Nature 2005, 433, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.; Burgueño, R. Buckling-induced smart applications: Recent advances and trends. Smart Mater. Struct. 2015, 24, 063001. [Google Scholar] [CrossRef]
- Overvelde, J.T.B.; Kloek, T.; D’haen, J.J.A.; Bertoldi, K. Amplifying the response of soft actuators by harnessing snap-through instabilities. Proc. Natl. Acad. Sci. USA 2015, 112, 10863–10868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmeyer, A.; Santer, M.; Pirrera, A.; Weaver, P.M. Pseudo-bistable self-actuated domes for morphing applications. Int. J. Solids Struct. 2012, 49, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- McKittrick, J.; Chen, P.Y.; Tombolato, L.; Novitskaya, E.E.; Trim, M.W.; Hirata, G.A.; Olevsky, E.A.; Horstemeyer, M.F.; Meyers, M.A. Energy absorbent natural materials and bioinspired design strategies: A review. Mater. Sci. Eng. C 2010, 30, 331–342. [Google Scholar] [CrossRef]
- Chen, I.H.; Kiang, J.H.; Correa, V.; Lopez, M.I.; Chen, P.Y.; McKittrick, J.; Meyers, M.A. Armadillo armor: Mechanical testing and micro-structural evaluation. J. Mech. Behav. Biomed. Mater. 2011, 4, 713–722. [Google Scholar] [CrossRef]
- Krauss, S.; Monsonego-Ornan, E.; Zelzer, E.; Fratzl, P.; Shahar, R. Mechanical function of a complex three-dimensional suture joining the bony elements in the shell of the red-eared slider turtle. Adv. Mater. 2009, 21, 407–412. [Google Scholar] [CrossRef]
- Amini, S.; Miserez, A. Wear and abrasion resistance selection maps of biological materials. Acta Biomater. 2013, 9, 7895–7907. [Google Scholar] [CrossRef]
- Müller, M.; Olek, M.; Giersig, M.; Schmitz, H. Micromechanical properties of consecutive layers in specialized insect cuticle: The gula of Pachnoda marginata (Coleoptera, Scarabaeidae) and the infrared sensilla of Melanophila acuminata (Coleoptera, Buprestidae). J. Exp. Biol. 2008, 211, 2576–2583. [Google Scholar] [CrossRef] [Green Version]
- Harvey, D.J.; Hawes, C.J.; Gange, A.C.; Finch, P.; Chesmore, D.; Farr, I. Development of non-invasive monitoring methods for larvae and adults of the stag beetle, Lucanus cervus. Insect Conserv. Divers. 2011, 4, 4–14. [Google Scholar] [CrossRef]
- Ha, N.S.; Le, V.T.; Goo, N.S. Investigation of Punch Resistance of the Allomyrira dichtoloma Beetle Forewing. J. Bionic Eng. 2018, 15, 57–68. [Google Scholar] [CrossRef]
- Zhu, D.; Szewciw, L.; Vernerey, F.; Barthelat, F. Puncture resistance of the scaled skin from striped bass: Collective mechanisms and inspiration for new flexible armor designs. J. Mech. Behav. Biomed. Mater. 2013, 24, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.J.; Triblehorn, J.D. Mechanical properties of the cuticles of three cockroach species that differ in their wind-evoked escape behavior. PeerJ 2014, 2, e501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundanati, L.; Signetti, S.; Gupta, H.S.; Menegon, M.; Pugno, N.M. Multilayer stag beetle elytra perform better under external loading via non-symmetric bending properties. J. R. Soc. Interface 2018, 15, 20180427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abràmoff, M.D.; Magalhães, P.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Fabritius, H.; Sachs, C.; Raabe, D.; Nikolov, S.; Friák, M.; Neugebauer, F. Chitin in the exoskeletons of arthropoda: From ancient design to novel materials science. In Chitin; Springer: Dordrecht, The Netherlands, 2011; p. 182. [Google Scholar] [CrossRef]
- Riks, E. An incremental approach to the solution of snapping and buckling problems. Int. J. Solids Struct. 1979, 15, 529–551. [Google Scholar] [CrossRef]
- Dai, Z.; Yang, Z. Macro-/Micro-Structures of Elytra, Mechanical Properties of the Biomaterial and the Coupling Strength Between Elytra in Beetles. J. Bionic Eng. 2010, 7, 6–12. [Google Scholar] [CrossRef]
- Gorb, S.N. Frictional Surfaces of the Elytra-to-Body Arresting Mechanism in Tenebrionid beetles (Coleoptera: Tenebrionidae): Design of co-opted fields of microtrichia and cutilcle ultrastructure. Int. J. Insect Morphol. Embryol. 1998, 27, 205–225. [Google Scholar] [CrossRef]
- Schultz, M.R.; Hyer, M.W. Snap-through of unsymmetric cross-ply laminates using piezoceramic actuators. J. Intell. Mater. Syst. Struct. 2003, 14, 795–814. [Google Scholar] [CrossRef]
- Brampton, C.J.; Betts, D.N.; Bowen, C.R.; Kim, H.A. Sensitivity of bistable laminates to uncertainties in material properties, geometry and environmental conditions. Compos. Struct. 2013, 102, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Dano, M.L.; Hyer, M.W. SMA-induced snap-through of unsymmetric fiber-reinforced composite laminates. Int. J. Solids Struct. 2003, 40, 5949–5972. [Google Scholar] [CrossRef]
- Arena, G.; Groh, R.M.J.; Brinkmeyer, A.; Theunissen, R.; Weaver, P.M.; Pirrera, A. Adaptive compliant structures for flow regulation. Proc. R. Soc. A Math. Phys. Eng. Sci. 2017, 473, 20170334. [Google Scholar] [CrossRef] [PubMed]
- Bellini, P.X. The concept of Snap-buckling illustrated by a simple model. Int. J. Non Linear. Mech. 1972, 7, 643–650. [Google Scholar] [CrossRef]
- Bazzucchi, F.; Manuello, A.; Carpinteri, A. Interaction between snap-through and Eulerian instability in shallow structures. Int. J. Non Linear Mech. 2017, 88, 11–20. [Google Scholar] [CrossRef]
- Sumirin, S.; Nuroji, N.; Besari, S. International Journal of Science and Engineering (IJSE) Snap-Through Buckling Problem of Spherical Shell Structure. Int. J. Sci. Eng. 2015, 8, 54–59. [Google Scholar] [CrossRef]
- Di Landro, L.; Sala, G.; Olivieri, D. Deformation mechanisms and energy absorption of polystyrene foams for protective helmets. Polym. Test. 2002, 21, 217–228. [Google Scholar] [CrossRef]
- Goyens, J.; Dirckx, J.; Dierick, M.; Van Hoorebeke, L.; Aerts, P. Biomechanical determinants of bite force dimorphism in Cyclommatus metallifer stag beetles. J. Exp. Biol. 2014, 217, 1065–1071. [Google Scholar] [CrossRef] [Green Version]
- Broomell, C.C.; Zok, F.W.; Waite, J.H. Role of transition metals in sclerotization of biological tissue. Acta Biomater. 2008, 4, 2045–2051. [Google Scholar] [CrossRef]
- Czapla, T.; Hopkins, T.; Kramer, K. Cuticular strains strength and pigmentation of five of adult blattella germanica (L.) during sclerotization: Correlations with catecholamines. J. Insect Physiol. 1990, 36, 647–654. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Lin, A.Y.-M.; McKittrick, J.; Meyers, M.A. Structure and mechanical properties of crab exoskeletons. Acta Biomater. 2008, 4, 587–596. [Google Scholar] [CrossRef]
- Klocke, D.; Schmitz, H. Water as a major modulator of the mechanical properties of insect cuticle. Acta Biomater. 2011, 7, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Politi, Y.; Priewasser, M.; Pippel, E.; Zaslansky, P.; Hartmann, J.; Siegel, S.; Li, C.; Barth, F.G.; Fratzl, P. A spider’s fang: How to design an injection needle using chitin-based composite material. Adv. Funct. Mater. 2012, 22, 2519–2528. [Google Scholar] [CrossRef]
- Lichtenegger, H.C.; Schöberl, T.; Bartl, M.H.; Waite, H.; Stucky, G.D. High abrasion resistance with sparse mineralization: Copper biomineral in worm jaws. Science 2002, 298, 389–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Zhang, P.; Wang, S. Recent progress of abrasion-resistant materials: Learning from nature. Chem. Soc. Rev. 2015, 45, 237–251. [Google Scholar] [CrossRef]
- Goyens, J.; Wassenbergh, S.; Van Dirckx, J.; Aerts, P. Cost of flight and the evolution of stag beetle weaponry. J. R. Soc. Interface 2015, 12, 20150222. [Google Scholar] [CrossRef] [Green Version]
- Munk, Y.; Yanoviak, S.P.; Koehl, M.A.R.; Dudley, R. The descent of ant: Field-measured performance of gliding ants. J. Exp. Biol. 2015, 218, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Yanoviak, S.P.; Munk, Y.; Dudley, R. Arachnid aloft: Directed aerial descent in neotropical canopy spiders. J. R. Soc. Interface 2015, 12, 20150534. [Google Scholar] [CrossRef]
- Yanoviak, S.P.; Dudley, R.; Kaspari, M. Directed aerial descent in canopy ants. Nature 2005, 433, 624–626. [Google Scholar] [CrossRef]
- Reichel, S.V.; Labisch, S.; Dirks, J.-H. What goes up must come down Biomechanical impact analysis of jumping locusts. J. Exp. Biol. 2019, 222, 1–15. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tip Diameter (µm) Measured at 100 µm from the Tip | Average Puncture Force (N) | Puncture Pressure (kPa) | |
|---|---|---|---|
| Mandible | 338 | 6.9 | 64 |
| Needle | 180 | 1.8 | 597 |
| Terminal Velocity (vmax) (m/s) | Height to Reach Terminal Velocity, Hmax (m) | Optimal Velocity (v) | Height to Reach Optimal Velocity, H (m) | Maximum Kinetic Energy (Ek_max) (mJ) | Energy Absorbed by Elytra, (2Ea) (mJ) |
|---|---|---|---|---|---|
| 9.3 | 4.4 | 1.8 | 0.35 | 72.6 | 5.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kundanati, L.; Guarino, R.; Pugno, N.M. Stag Beetle Elytra: Localized Shape Retention and Puncture/Wear Resistance. Insects 2019, 10, 438. https://doi.org/10.3390/insects10120438
Kundanati L, Guarino R, Pugno NM. Stag Beetle Elytra: Localized Shape Retention and Puncture/Wear Resistance. Insects. 2019; 10(12):438. https://doi.org/10.3390/insects10120438
Chicago/Turabian StyleKundanati, Lakshminath, Roberto Guarino, and Nicola M. Pugno. 2019. "Stag Beetle Elytra: Localized Shape Retention and Puncture/Wear Resistance" Insects 10, no. 12: 438. https://doi.org/10.3390/insects10120438
APA StyleKundanati, L., Guarino, R., & Pugno, N. M. (2019). Stag Beetle Elytra: Localized Shape Retention and Puncture/Wear Resistance. Insects, 10(12), 438. https://doi.org/10.3390/insects10120438