Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Interactions and Chemical Crosstalk between Microorganisms, Plants, and Insects: Ecological Models and Study Cases
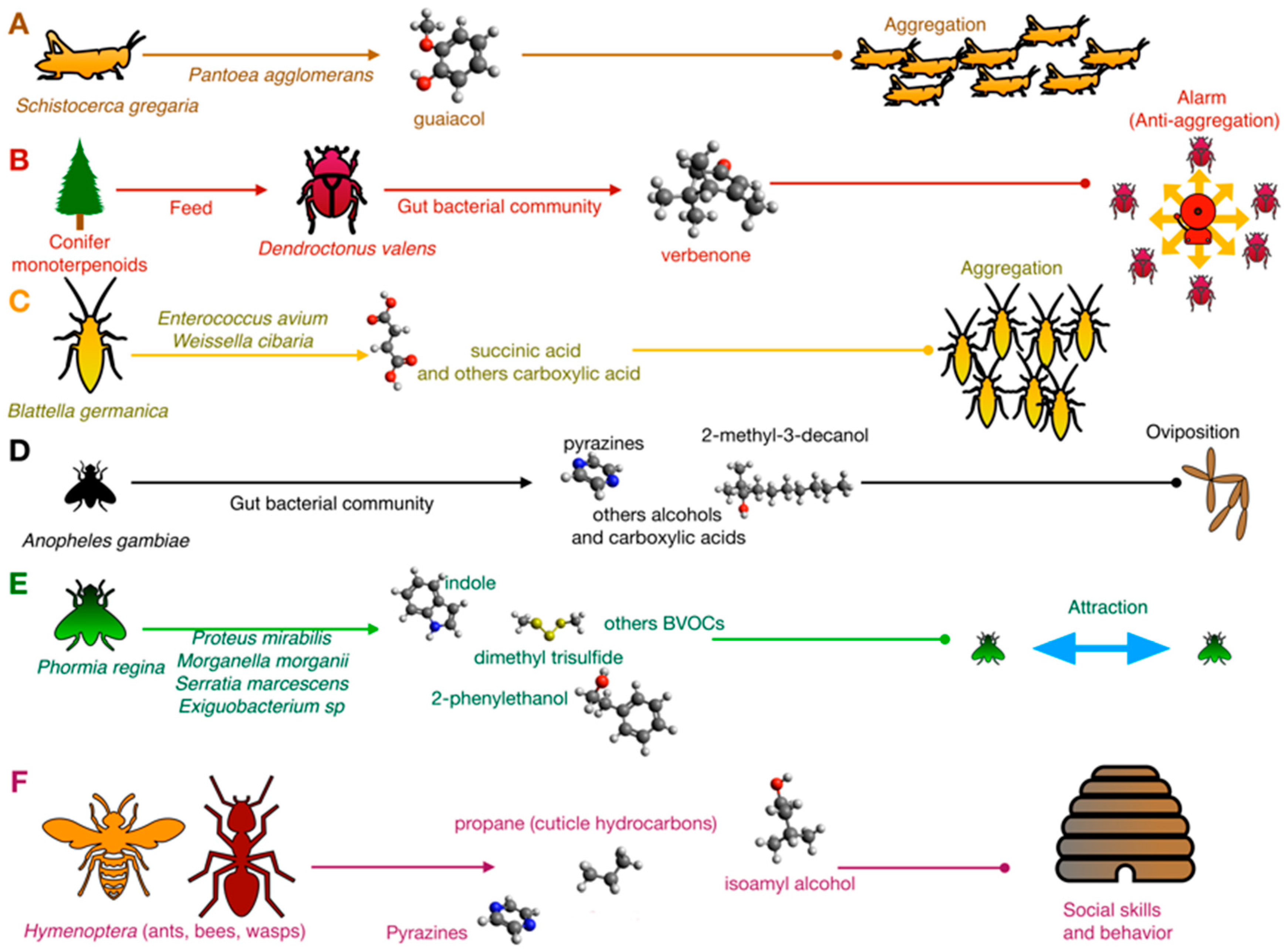
2.1. Bacteria and Presocial Insect Communication
2.2. Bacteria and Eusocial Insect Communication
2.3. Bacteria and Sexual Communication in Insects
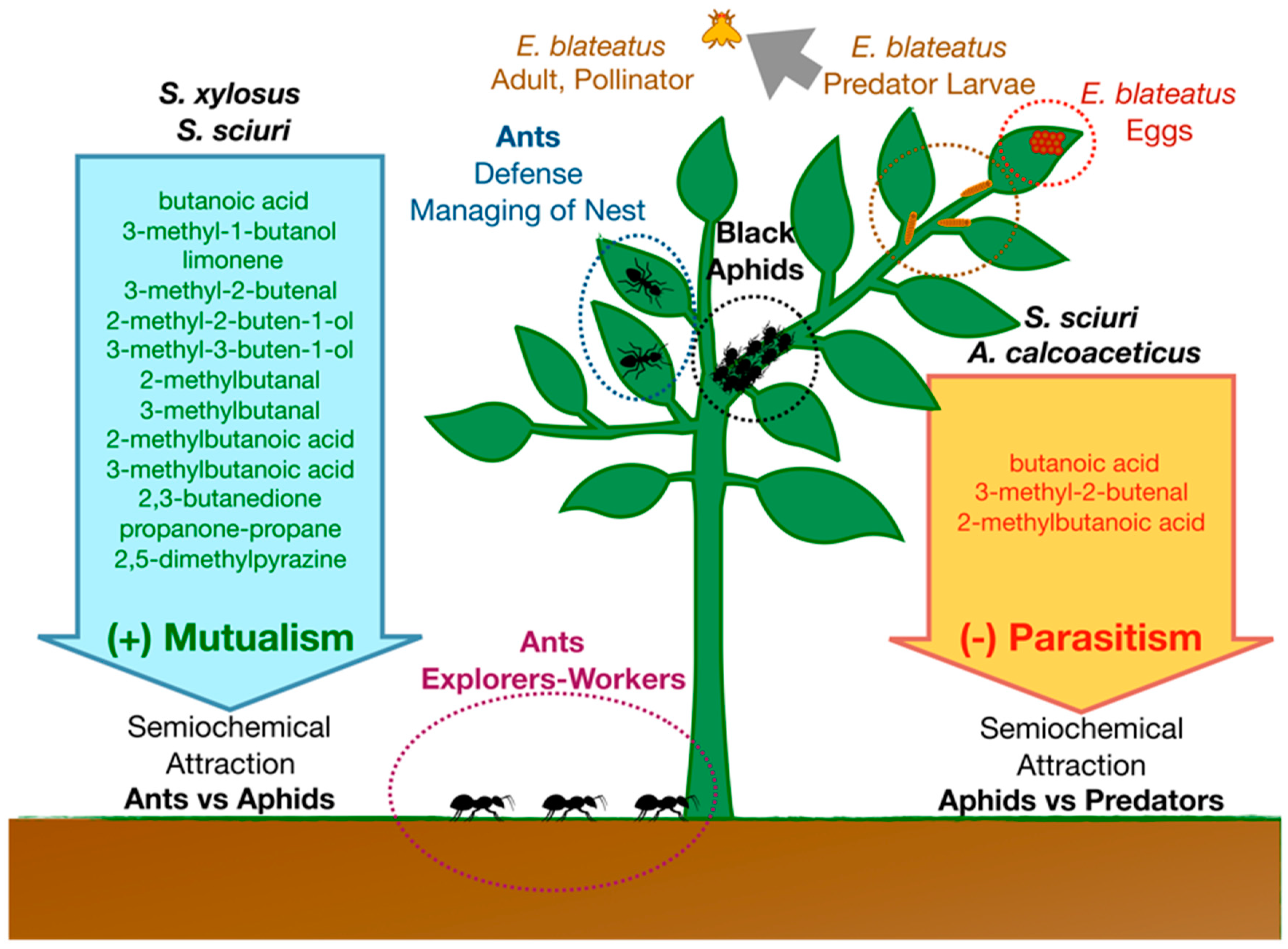
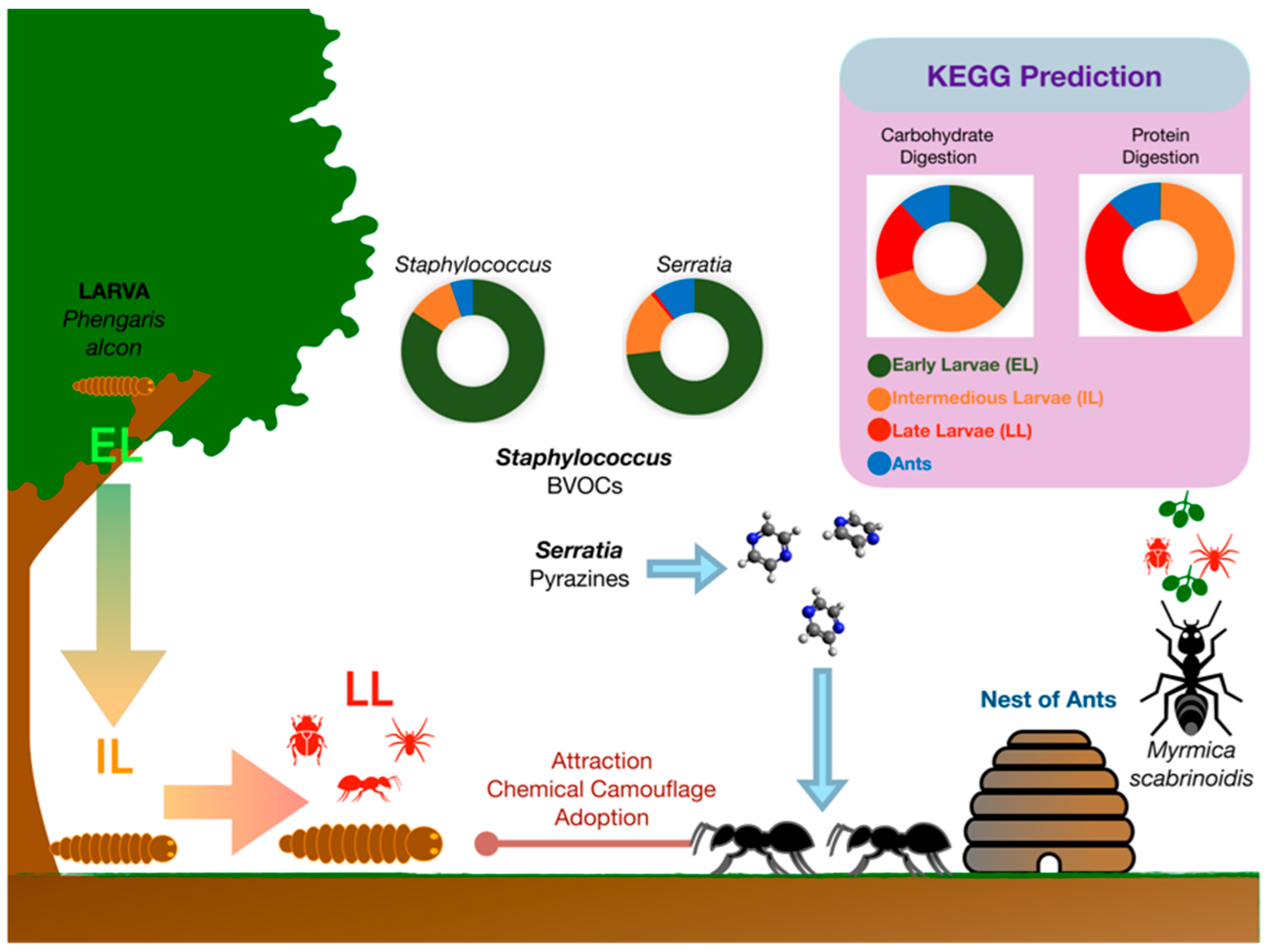
2.4. Bacteria and Interspecies Insect Communication
2.5. Bacteria and Insect–Plant Communication
3. BVOC Biosynthetic Pathways and Their Intersections with Primary and Secondary Metabolism
3.1. BVOCs and Primary Metabolism
3.2. BVOCs and Secondary Metabolism
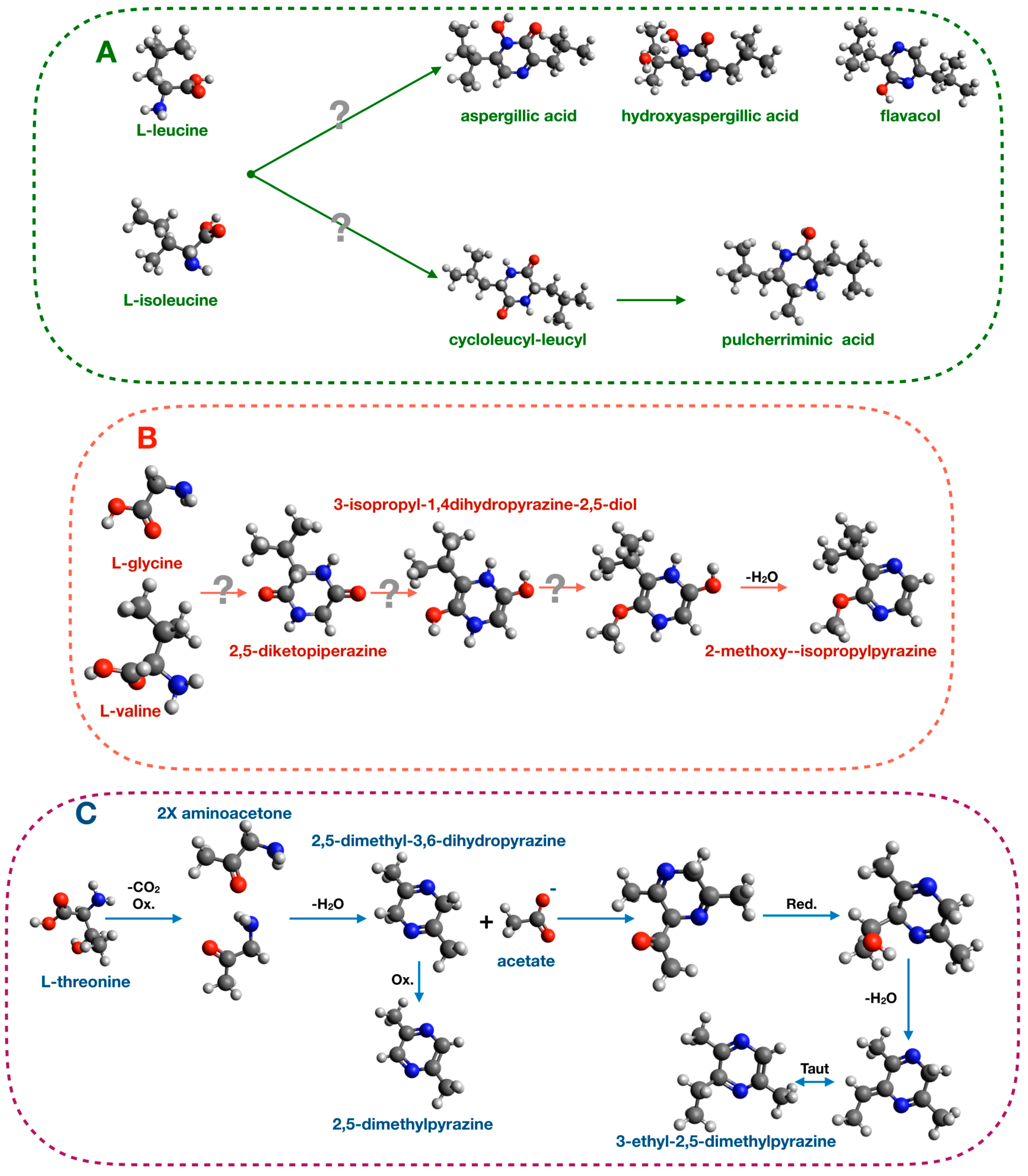
3.2.1. Pyrazine Metabolism in Bacteria
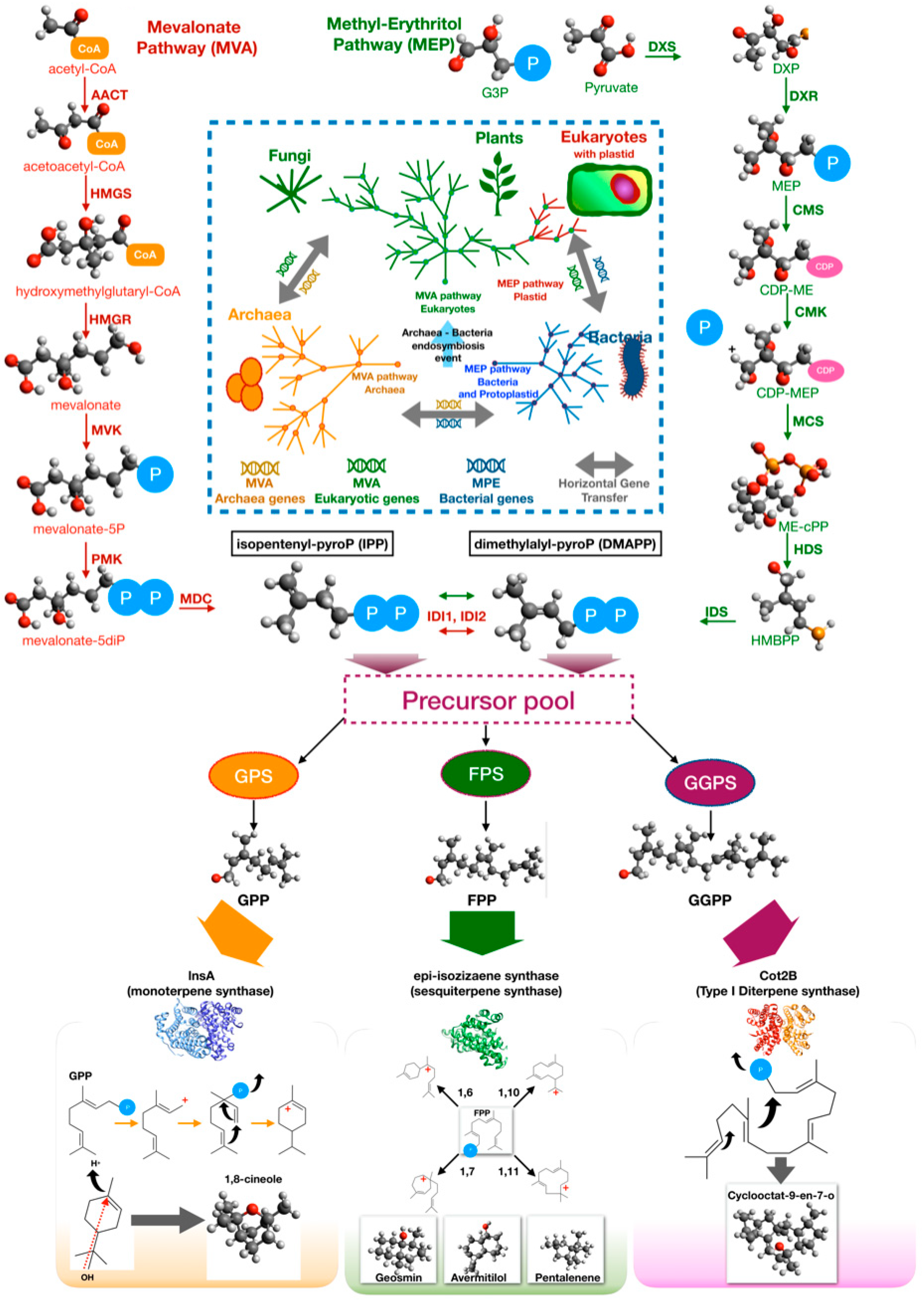
3.2.2. Terpene and Terpenoid Metabolism in Bacteria
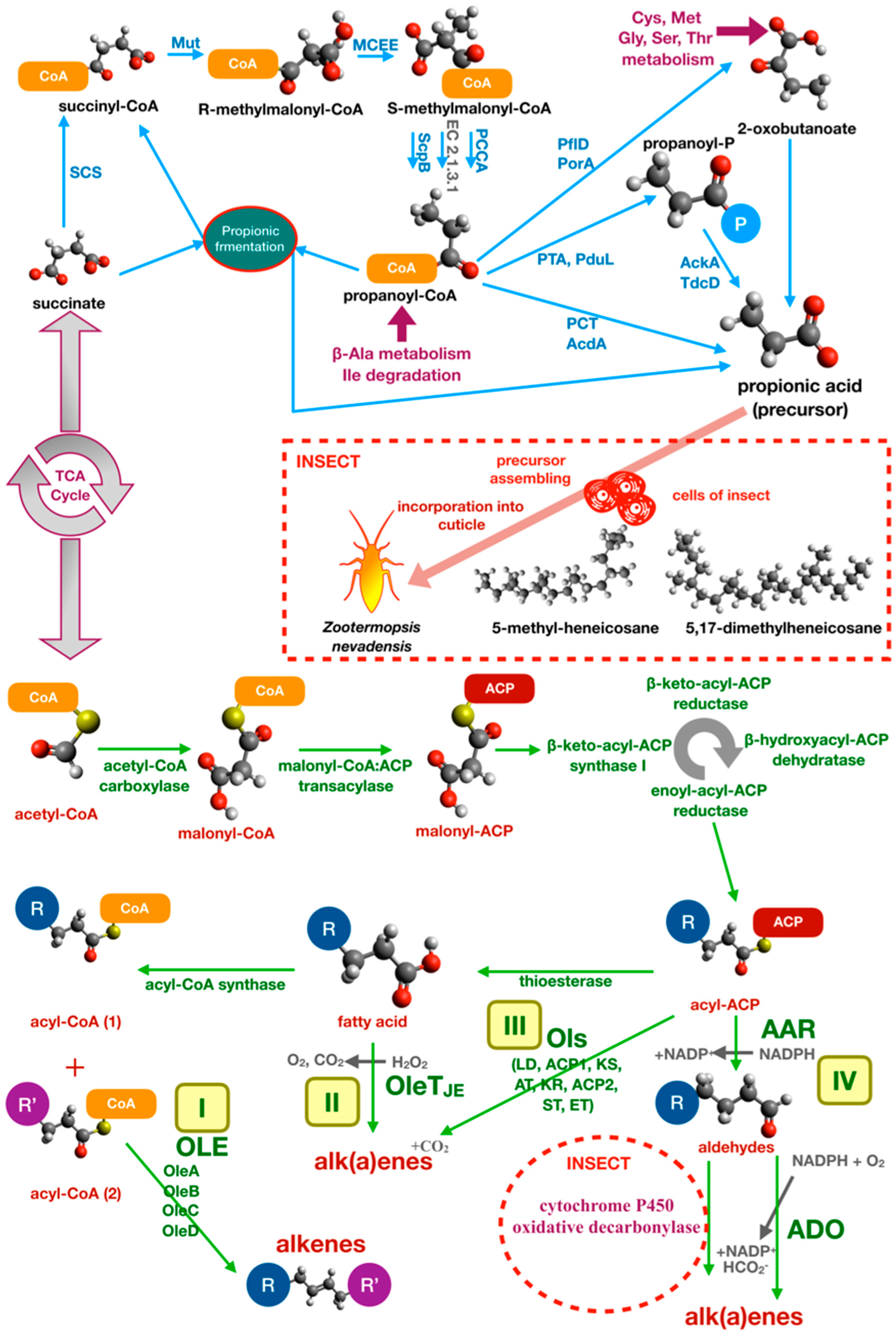
3.2.3. Alkane and Alkene Metabolism in Bacteria
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol. Agron Soc. Environ. 2011, 15, 327–337. [Google Scholar]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 3, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.K.; Kasotia, A.; Jain, S.; Vaishnav, A.; Kumari, S.; Sharma, K.P.; Varma, A. Bacterial-Mediated Tolerance and Resistance to Plants Under Abiotic and Biotic Stresses. J. Plant Growth Regul. 2016, 35, 276–300. [Google Scholar] [CrossRef]
- Heil, M.; Bostock, R.M. Induced Systemic Resistance (ISR) Against Pathogens in the Context of Induced Plant Defences. Ann. Bot. 2002, 89, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The Interactions of Rhizodeposits with Plant Growth-Promoting Rhizobacteria in the Rhizosphere: A Review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Vavre, F.; Kremer, N. Microbial impacts on insect evolutionary diversification: From patterns to mechanisms. Curr. Opin Insect Sci. 2014, 4, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Zheng, S.; van Loon, J.J.A.; Pieterse, C.M.J.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef]
- Ueda, K.; Tawaraya, K.; Murayama, H.; Sato, S.; Nishizawa, T.; Toyomasu, T.; Murayama, T.; Shiozawa, S.; Yasuda, H. Effects of arbuscular mycorrhizal fungi on the abundanceof foliar-feeding insects and their natural enemy. Appl. Entomol. Zool. 2013, 48, 79–85. [Google Scholar] [CrossRef]
- Hempel, S.; Stein, C.; Unsicker, S.B.; Renker, C.; Auge, H.; Weisser, W.W.; Buscot, F. Specific bottom–up effects of arbuscular mycorrhizal fungi across a plant–herbivore–parasitoid system. Oecologia 2009, 160, 267. [Google Scholar] [CrossRef] [Green Version]
- Zebelo, S.A.; Bertea, C.M.; Bossi, S.; Occhipinti, A.; Gnavi, G.; Maffei, M.E. Chrysolina herbacea Modulates Terpenoid Biosynthesis of Mentha aquatica L. PLoS ONE 2011, 6, e17195. [Google Scholar] [CrossRef] [Green Version]
- Engl, T.; Kaltenpot, M. Influence of microbial symbionts on insect pheromones. Nat. Prod. Rep. 2018, 35, 386. [Google Scholar] [CrossRef] [PubMed]
- Leroy, P.D.; Sabri, A.; Verheggen, F.J.; Francis, F.; Thonart, P.; Haubruge, E. The semiochemically mediated interactions between bacteria and insects. Chemoecology 2011, 21, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Van der Meer, R. Ant Interactions with Soil Organisms and Associated Semiochemicals. J. Chem. Ecol. 2012, 38, 728–745. [Google Scholar] [CrossRef] [PubMed]
- Wielkopolan, B.; Obrępalska-Stęplowska, A. Three-way interaction among plants, bacteria, and coleopteran insects. Planta 2016, 244, 313–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kather, R.; Martin, S.J.J. Evolution of Cuticular Hydrocarbons in the Hymenoptera: A Meta-Analysis. Chem. Ecol. 2015, 41, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schettino, M.; Grasso, D.A.; Weldegergis, B.T.; Castracani, C.; Mori, A.; Dicke, M.; Van Lenteren, J.C.; Van Loon, J.J.A. Response of a Predatory ant to Volatiles Emitted by Aphid-and Caterpillar-Infested Cucumber and Potato Plants. J. Chem. Ecol. 2017, 43, 1007. [Google Scholar] [CrossRef]
- Beck, J.J.; Alborn, H.T.; Block, A.K.; Christensen, S.A.; Hunter, C.T.; Rering, C.C.; Seidl-Adams, I.; Stuhl, C.J.; Torto, B.; Tumlinson, J.H. Interactions Among Plants, Insects, and Microbes: Elucidation of Inter-Organismal Chemical Communications in Agricultural Ecology. J. Agric. Food Chem. 2018, 66, 6663–6674. [Google Scholar] [CrossRef]
- Jordan, H.R.; Tomberlin, J.K. Abiotic and Biotic Factors Regulating Inter-Kingdom Engagement between Insects and Microbe Activity on Vertebrate Remains. Insects 2017, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial Volatile Emissions as Insect Semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Dillon, R.; Charnley, K. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef]
- Dillon, R.; Vennard, C.T.; Charnley, A.K. Exploitation of gut bacteria in the locust. Nature 2000, 403, 851. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lou, Q.; Cheng, C.; Lu, M.; Sun, J. Gut-Associated Bacteria of Dendroctonus valens and their Involvement in Verbenone Production. Microb. Ecol. 2015, 70, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Boone, C.K.; Bohlmann, J.; Raffa, K.F. Responses of Bark Beetle-Associated Bacteria to Host Monoterpenes and Their Relationship to Insect Life Histories. J. Chem. Ecol. 2011, 37, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Boone, C.K.; Keefover-Ring, K.; Mapes, A.C.; Adams, A.S.; Bohlmann, J.; Raffa, K.F. Bacteria Associated with a Tree-Killing Insect Reduce Concentrations of Plant Defense Compounds. J. Chem. Ecol. 2013, 39, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurekc, L.; Nalyanya, G.; Roelofs, W.L.; Zhangf, A.; Schala, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 15678–15683. [Google Scholar] [CrossRef] [Green Version]
- Uriel, Y.; Gries, R.; Tu, L.; Carroll, C.; Zhai, H.; Moore, M.; Gries, G. The fly factor phenomenon is mediated by interkingdom signaling between bacterial symbionts and their blow fly hosts. Insect Sci. 2018, 1–10. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Diez, L.; Sablon, L.; Fischer, C.; Bartram, S.; Haubruge, E.; Detrain, C. Aphid alarm pheromone as a cue for ants to locate aphid partners. PLoS ONE 2012, 7, e41841. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.Y.; Lognay, G.C.; Detrain, C.; Heil, M.; Grigorescu, A.; Sabri, A.; Thonart, P.; Haubruge, E.; Verheggen, F.J. Bacteria may enhance species association in an ant–aphid mutualistic relationship. Chemoecology 2015, 25, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.Y.; Detrain, C.; Thonart, P.; Haubruge, E.; Francis, F.; Verheggen, J.F.; Lognay, G.C. Bacteria may contribute to distant species recognition in ant–aphid mutualistic relationships. Insect Sci. 2017, 24, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Francis, F.; Vandermoten, S.; Verheggen, F.; Lognay, G.C.; Haubruge, E. Is the (E)-β-farnesene only volatile terpenoid in aphids? J. Appl. Entomol. 2005, 129, 6–11. [Google Scholar] [CrossRef]
- Sumba, L.A.; Guda, T.O.; Deng, A.L.; Hassanali, A.; Beier, J.C.; Knols, B.G.J. Mediation of oviposition site selection in the African malaria mosquito Anopheles gambiae (Diptera: Culicidae) by semiochemicals of microbial origin. Int. J. Trop. Insect Sci. 2004, 24, 260–265. [Google Scholar] [CrossRef]
- Lindh, J.M.; Kännaste, A.; Knols, B.G.J.; Faye, I.; Borg-Karlson, A.K. Oviposition Responses of Anopheles gambiae s.s. (Diptera: Culicidae) and Identification of Volatiles from Bacteria-Containing Solutions. J. Med. Entomol. 2008, 45, 45–1039. [Google Scholar] [CrossRef] [Green Version]
- Verhulst, N.O.; Qiu, Y.T.; Beijleveld, H.; Maliepaard, C.; Knights, D.; Schulz, S.; Berg-Lyons, D.; Lauber, C.L.; Verduijn, W.; Haasnoot, G.W.; et al. Composition of Human Skin Microbiota Affects Attractiveness to Malaria Mosquitoes. PLoS ONE 2011, 6, e28991. [Google Scholar] [CrossRef]
- Verhulst, N.O.; Beijleveld, H.; Knols, B.G.J.; Takken, W.; Schraa, G.; Bouwmeester, H.J.; Smallegange, R.C. Cultured skin microbiota attracts malaria mosquitoes. Malar. J. 2009, 8, 302d. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Crippen, T.L.; Coates, C.J.; Wood, T.K.; Tomberlin, J.K. Effect of Quorum Sensingby Staphylococcus epidermidis on the Attraction Response of Female Adult Yellow Fever Mosquitoes, Aedes aegypti aegypti (Linnaeus) (Diptera:Culicidae), to a Blood-Feeding Source. PLoS ONE 2015, 10, e0143950. [Google Scholar] [CrossRef] [Green Version]
- Silva-Junior, E.A.; Ruzzini, A.C.; Paludo, C.R.; Nascimento, F.S.; Currie, C.R.; Clardy, J.; Pupo, M.T. Pyrazines from bacteria and ants: Convergent chemistry within an ecological niche. Sci. Rep. 2018, 8, 2595. [Google Scholar] [CrossRef] [Green Version]
- Babcock, T.; Borden, J.H.; Gries, R.; Carroll, C.; Lafontaine, J.P.; Moore, M.; Gries, G. Inter-kingdom signaling—Symbiotic yeasts produce semiochemicals that attract their yellowjacket hosts. Entomol. Exp. Appl. 2019, 167, 220–230. [Google Scholar] [CrossRef]
- Matsuura, K. Nestmate recognition mediated by intestinal bacteria in a termite, Reticulitermes speratus. Oikos 2001, 92, 20–26. [Google Scholar] [CrossRef]
- Nalepa, C.A. Origin of termite eusociality: Trophallaxis integrates the social, nutritional, and microbial environments. Ecol. Entomol. 2015, 40, 323–335. [Google Scholar] [CrossRef]
- Guo, L.; Quilici, D.R.; Chase, J.; Blomquis, G.J. Gut tract microorganisms supply the precursors for methyl-branched hydrocarbon biosynthesis in the termite, Zootermopsis nevadensis. Insect Biochem. 1991, 21, 327–333. [Google Scholar] [CrossRef]
- Teseo, S.; van Zweden, J.S.; Pontieri, L.; Kooij, P.W.; Sørensen, S.J.; Wenseleers, T.; Poulsen, M.; Boomsma, J.J.; Sapountzis, P. The scent of symbiosis: Gut bacteria may affect social interactions in leaf-cutting ants. Anim. Behav. 2019, 150, 239–254. [Google Scholar] [CrossRef]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of Acquisition of the Gut Microbiota of the Honey Bee Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.E.; Eiri, D.; Moran, N.A.; Rangel, J. Modulation of the honey bee queen microbiota: Effects of early social contact. PLoS ONE 2018, 13, e0200527. [Google Scholar] [CrossRef]
- Yun, J.H.; Jung, M.J.; Kim, P.S.; Bae, J.W. Social status shapes the bacterial and fungal gut communities of the honey bee. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kapheim, K.M.; Rao, V.D.; Yeoman, C.J.; Wilson, B.A.; White, B.A.; Goldenfeld, N.; Robinson, G.E. Caste-specific dif- ferences in hindgut microbial communities of honey bees (Apis mellifera). PLoS ONE 2015, 10, e0123911. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Mattila, H.R.; Newton, I.L.G. Development of the honey bee gut microbiome throughout the queen-rearing process. Appl. Environ. Microbiol. 2015, 81, 3182–3191. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.C.; Fruciano, C.; Hildebrand, F.; Al Toufalilia, H.; Balfour, N.J.; Bork, P.; Engel, P.; Ratnieks, F.L.W.; Hughes, W.O.H. Gut microbiota composition is associated with environmental landscape in honey bees. Ecol. Evol. 2018, 8, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, D.; Narita, S.; Watanabe, M. Insect Sex Determination Manipulated by Their Endosymbionts: Incidences, Mechanisms and Implications. Insects 2012, 3, 161–199. [Google Scholar] [CrossRef] [Green Version]
- Burand, J.P.; Tan, W.J. Mate preference and mating behavior of male Helicoverpa zea (Lepidoptera: Noctuidae) infected with the sexually transmitted insect virus Hz-2V. Ann. Entomol. 2006, 99, 969–973. [Google Scholar] [CrossRef]
- Wigul, G.; Sikorowski, P.P. The Effect of Staphylococcal Enterotoxin B on Pheromone Production in Fat Bodies Isolated from Male Boll Weevils. J. Invertebr. Pathol. 1986, 47, 116–119. [Google Scholar] [CrossRef]
- Hoyt, C.P.; Osborne, G.O.; Mulcock, A.P. Production of an insect sex attractant by symbiotic bacteria. Nature 1971, 230, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.G.; Jackson, T.A.; Unelius, C.R.; Wee, S.L.; Young, S.D.; Townsend, R.J.; Suckling, D.M. Morganella morganii bacteria produces phenol as the sex pheromone of the New Zealand grass grub from tyrosine in the colleterial gland. Sci. Nat. 2016, 103, 59. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Lautenschläger, B.; Heinze, J. A transitional stage between the ergatoid and winged male morph in the ant Cardiocondyla obscurior. Insectes Soc. 2002, 49, 221–228. [Google Scholar] [CrossRef]
- Heinze, J. Life-history evolution in ants: The case of Cardiocondyla. Proc. R Soc. 2017, B284, 20161406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.; Schrader, L.; Gil, R.; Manzano-Marin, A.; Florez, L.; Wheeler, D.; Werren, J.H.; Latorre, A.; Heinze, J.; Kaltenpoth, M.; et al. A novel intracellular mutualistic bacterium in the invasive ant Cardiocondyla obscurior. ISME J. 2016, 10, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzolante, G.; Cordero, C.; Tredici, S.M.; Vergara, D.; Pontieri, P.; Del Giudice, L.; Capuzzo, A.; Rubiolo, P.; Kanchiswamy, C.N.; Zebelo, S.A.; et al. Cultivable gut bacteria provide a pathway for adaptation of Chrysolina herbacea to Mentha aquatica volatiles. BMC Plant Biol. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Maffei, M.E.; Chialva, F.; Sacco, T. Glandular trichomes and essential oil in developing peppermint leaves. I. Variation in peltate trichome number and terpene distribution within leaves. New Phytol. 1989, 111, 707–716. [Google Scholar]
- Wood, W.F.; Palmer, T.M.; Stanton, M.L. A comparison of volatiles in mandibular glands from three Crematogaster ant symbionts of the whistling thorn acacia. Biochem. Syst. Ecol. 2002, 30, 217–222. [Google Scholar] [CrossRef]
- El-Sayed, A.M. The Pherobase: Database of Pheromones and Semiochemicals. Available online: http://www.pherobase.com (accessed on 10 November 2019).
- Whitehead, S.R.; Reid, E.; Sapp, J.; Poveda, K.; Royer, AM.; Posto, A.L.; Kessler, A. A Specialist Herbivore Uses Chemical Camouflage to Overcome the Defenses of an Ant-Plant Mutualism. PLoS ONE 2014, 9, e102604. [Google Scholar] [CrossRef]
- Oliver, T.H.; Mashanova, A.; Leather, S.R.; Cook, J.M.; Jansen, V.A.A. Ant semiochemicals limit apterous aphid dispersal. Proc. R. Soc. B 2007, 274, 3127–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, P.D.; Sabri, A.; Heuskin, S.; Thonart, P.; Lognay, G.; Verheggen, F.J.; Francis, F.; Brostaux, Y.; Felton, G.W.; Haubruge, E. Microorganisms from aphid honeydew attract and enhance the efficacy of natural enemies. Nat. Commun. 2011, 2, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, N.E.; Braby, M.F.; Heath, A.; Lohman, D.J.; Mathew, J.; Rand, D.B.; Travassos, M.A. The ecology and evolution of ant association in the Lycaenidae (Lepidoptera). Annu. Rev. Entomol. 2002, 47, 733–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Als, T.D.; Nash, D.R.; Boomsma, J.J. Adoption of parasitic Maculinea alcon caterpillars (Lepidoptera:Lycaenidae) by three Myrmica ant species. Anim. Behav. 2001, 62, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Barbero, F. Cuticular Lipids as a Cross-Talk among Ants, Plants and Butterflies. Int. J. Mol. Sci. 2016, 17, 1966. [Google Scholar] [CrossRef] [Green Version]
- Di salvo, M.; Calcagnile, M.; Talà, A.; Tredici, S.M.; Maffei, M.E.; Schönrogge, K.; Barbero, F.; Alifano, P. The Microbiome of the Maculinea-Myrmica Host-parasite Interaction. Sci. Rep. 2019, 9, 8048. [Google Scholar] [CrossRef] [Green Version]
- Szenteczki, M.A.; Pitteloud, C.; Casacci, L.P.; Kešnerová, L.; Whitaker, M.R.L.; Enge, P.; Vila, R.; Alvarez, N. Bacterial communities within Phengaris (Maculinea) alcon caterpillars are shifted following transition from solitary living to social parasitism of Myrmica ant colonies. Ecol. Evol. 2019, 9, 4452–4464. [Google Scholar] [CrossRef] [Green Version]
- Kronauer, D.J.C.; Pierce, N.E. Myrmecophiles. Curr. Biol. 2011, 21, 6. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T. Symbiotic bacterium modifies aphid body color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef]
- Menzel, R.; Backhaus, W. Colour vision in insects. In Vision and Visual Dysfunction; Gouras, P., Ed.; Mac-Millan: London, UK, 1991; pp. 262–293. [Google Scholar]
- Greenfield, J.C. Vetiver Grass: The Hedge Against Erosion; World Bank: Washington, DC, USA, 1990; ISBN 10: 082131405X. [Google Scholar]
- Babalola, O.; Oshunsanya, S.O.; Are, K. Effects of vetiver grass (Vetiveria nigritana) strips, vetiver grassmulch and an organomineral fertilizer on soil, water andnutrient losses and maize (Zea mays, L.) yields. Soil Tillage Res. 2007, 96, 6–18. [Google Scholar] [CrossRef]
- Danh, L.T.; Truong, P.; Mammucari, R.; Foster, N. Economic Incentive for Applying Vetiver Grass to Remediate Lead, Copper and Zinc Contaminated Soils. Int. J. Phytoremed. 2010, 13, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Brandt, R.; Merkl, N.; Schultze-Kraft, R.; Infante, C.; Broll, G. Potential of vetiver (Vetiveria zizanioides (L.) Nash) for phytoremediation of petroleum hydrocarbon-contaminated soils in Venezuela. Int. J. Phytoremed. 2006, 8, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E. Vetiveria: The genus Vetiveria; Taylor & Francis Inc.: New York, NY, USA, 2002; ISBN 0-203-27411-3. [Google Scholar]
- Yoon, P.K. Extracts from A Look-See at Vetiver Grass in Malaysia-First Progress Report. Vetiver Newsl. 1991, 6, 86–96. [Google Scholar]
- Zisong, W. Excerpts from the experiments and popularization of vetiver grass, Nanpang Prefecture. Vetiver Newsl. 1991, 6, 105–108. [Google Scholar]
- Van den Berg, J. Vetiver grass (Vetiveria zizanioides (L.) Nash) as trap plant for Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae) and Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Ann. Soc. Entomol. Fr. 2006, 42, 449–454. [Google Scholar] [CrossRef]
- Van den Berg, J. Can Vetiver Grass be Used to Manage Insect Pests on Crops? In Proceedings of the Third Internatioal Vetiver Conference, Guangzhou, China, 6–9 October 2003. [Google Scholar]
- Chomchalow, N. Vetiver: A Living Trap; PRVN Tech. Bull.No. 2015/2; ORDPB: Bangkok, Thailand, 2015. [Google Scholar]
- Jain, S.C.; Nowicki, S.; Eisner, T.; Meinnald, J. Insect repellents from vetiver oil: Zizanal and epizizanal. Tetrahedron Lett. 1982, 23, 4639–4642. [Google Scholar] [CrossRef]
- Zhu, B.C.; Henderson, G.; Chen, F.; Maistrello, L.; Laine, R.A. Nootkatone is a repellent for Formosan subterranean termite (Coptotermes formosanus). J. Chem. Ecol. 2001, 27, 523–531. [Google Scholar] [CrossRef]
- Maistrello, L.; Henderson, G.; Laine, R.A. Efficacy of vetiver oil and nootkatone as soil barriers against Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2001, 94, 1532–1537. [Google Scholar] [CrossRef]
- Alifano, P.; Del Giudice, L.; Talà, A.; De Stefano, M.; Maffei, M.E. Microbes at work in perfumery: The microbial community of vetiver root and its involvement in essential oil biogenesis. Flavour Fragr. J. 2010, 25, 121–122. [Google Scholar] [CrossRef]
- Del Giudice, L.; Massardo, D.R.; Pontieri, P.; Bertea, C.M.; Mombello, D.; Carata, E.; Tredici, S.M.; Talà, A.; Mucciarelli, M.; Ivanova Groudeva, V. The microbial community of Vetiver root and its involvement into essential oil biogenesis. Environ. Microbiol. 2008, 10, 2824–2841. [Google Scholar] [CrossRef]
- Rering, C.; Beck, J.J.; Hall, G.W.; McCartney, M.M.; Vannette, R.L. Nectar-inhabiting microorganisms influence nectar volatile composition and attractiveness to a generalist pollinator. New Phytol. 2017, 3, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schildknecht, H.; Koob, K. Myrmicacin, the first insect herbicide. Angew. Chem. Int. Ed. Engl. 1971, 10, 124–125. [Google Scholar] [CrossRef] [PubMed]
- Iwanami, Y.; Iwadare, T. Myrmic Acids: A Group of New Inhibitors Analogous to Myrmicacin (β-Hydroxydecanoic Acid). Bot. Gaz. 1979, 140, 1–4. [Google Scholar] [CrossRef]
- Sjögren, J.; Magnusson, J.; Broberg, A.; Schnürer, J.; Kenne, L. Antifungal 3-Hydroxy Fatty Acids from Lactobacillus plantarum MiLAB 14. Appl. Environ. Microbiol. 2003, 69, 7554–7557. [Google Scholar] [CrossRef] [Green Version]
- Barke, J.; Seipke, R.F.; Yu, D.W.; Hutchings, M.I. A mutualistic microbiome: How do fungus-growing ants select their antibiotic-producing bacteria? Commun. Integr. Biol. 2011, 4, 41–43. [Google Scholar] [CrossRef]
- Dicke, M. Plant phenotypic plasticity in the phytobiome: A volatile issue. Curr. Opin. Plant Biol. 2016, 32, 17–23. [Google Scholar] [CrossRef]
- Van de Mortel, J.E.; de Vos, R.C.H.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; van Loon, J.J.A.; Dicke, M.; Raaijmakers, J.M. Metabolic and Transcriptomic Changes Induced in Arabidopsis by the Rhizobacterium Pseudomonas fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; Van Loon, J.J.A.; Dicke, M. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef]
- Soler, R.; Badenes-Pérez, F.R.; Broekgaarden, C.; Zheng, S.J.; David, A.; Boland, W.; Dicke, M. Plant-mediated facilitation between a leaf-feeding and a phloem-feeding insect in a brassicaceous plant: From insect performance to gene transcription. Funct. Ecol. 2012, 26, 156–166. [Google Scholar] [CrossRef]
- Cusumano, A.; Zhu, F.; Volkoff, A.N.; Verbaarschot, P.; Bloem, J.; Vogel, H.; Dicke, M.; Poelman, E.H. Parasitic wasp-associated symbiont affects plant-mediated species interactions between herbivores. Ecol. Lett. 2018, 21, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef] [Green Version]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef] [Green Version]
- Kolesinska, B.; Fraczyk, J.; Binczarski, M.; Modelska, M.; Berlowska, J.; Dziugan, P.; Antolak, H.; Kaminski, Z.J.; Witonska, I.A.; Kregiel, D. Butanol Synthesis Routes for Biofuel Production: Trends and Perspectives. Materials 2019, 12, 350. [Google Scholar] [CrossRef] [Green Version]
- Bowman, K.S.; Rainey, F.A.; Moe, W.M. Production of hydrogen by Clostridium species in the presence of chlorinated solvents. FEMS Microbiol. Lett. 2009, 290, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, F. Über die bedingungen der fuselölbildung und über ihren zusammenhang mit dem eiweissaufbau der hefe. Ber. Dtsch. Chem. Ges. 1907, 40, 1027–1047. [Google Scholar] [CrossRef] [Green Version]
- Vogt, M.; Brüsseler, C.; van Ooyen, J.; Bott, M.; Marienhagen, J. Production of 2-methyl-1-butanol and 3-methyl-1-butanol in engineered Corynebacterium glutamicum. Metab. Eng. 2016, 38, 436–445. [Google Scholar] [CrossRef]
- Hasegawa, M.; Nishiza, A.W.A.; Tsuji, K.; Kimura, S.; Harada, K.I. Volatile Organic Compounds Derived from 2-Keto-Acid Decarboxylase in Microcystis aeruginosa. Microbes Environ. 2012, 27, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Connor, M.R.; Cann, A.F.; Liao, J.C. 3-Methyl-1-butanol production in Escherichia coli: Random mutagenesis and two-phase fermentation. Appl. Microbiol. Biotechnol. 2010, 86, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Sato, D.; Nozaki, T. Methionine gamma-lyase: The unique reaction mechanism, physiological roles, and therapeutic applications against infectious diseases and cancers. IUBMB Life 2009, 61, 1019–1028. [Google Scholar] [CrossRef]
- Kadota, H.; Ishida, Y. Production of volatile sulfur compounds by microorganisms. Annu. Rev. Microbiol. 1972, 26, 127–138. [Google Scholar] [CrossRef]
- Nakano, Y.; Yoshimura, M.; Koga, T. Correlation between oral malodor and periodontal bacteria. Microbes Infect. 2002, 4, 679–683. [Google Scholar] [CrossRef]
- Landaud, S.; Helinck, S.; Bonnarme, P. Formation of volatile sulfur compounds and metabolism of methionine and other sulfur compounds in fermented food. Appl. Microbiol. Biotechnol. 2008, 77, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Mafra-Neto, A.; Fettig, C.J.; Munson, S.; Stelinski, L.L. Use of repellents formulated in specialized pheromone and lure application technology for effective insect pest management. In Insect Repellents Handbook, 2nd ed.; Debboun, M., Francis, S.P., Strickman, D.A., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2014; Chapter 16; pp. 291–314. ISBN 9781466553583 1466553588; 9781466553552 1466553553. [Google Scholar]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, G.W.H. Recent advances in pyrazine chemistry. In Advances in Heterocyclic Chemistry; Katritzky, A.R., Boulton, A.J., Eds.; Academic Press Inc.: London, UK, 1972; pp. 99–209. ISBN 978-0120207787. [Google Scholar]
- Kurniadi, T.; Belrhild, R.; Berger, R.F.; Juillerat, M.A.; Fay, L.B. Preparation of Pyrazines. European Patent 1,357,193, 29 October 2003. [Google Scholar]
- Moser, B.R. Review of cytotoxic cephalostatins and ritterazines: Isolation and synthesis. J. Nat. Prod. Rep. 2008, 71, 487–491. [Google Scholar] [CrossRef]
- Prochaska, H.J.; Yeh, Y.; Baron, P.; Polsky, B. Oltipraz (OLT), a novel inhibitor of HIV-1 replication. J. Cell. Biochem. 1995, 59, 117–125. [Google Scholar] [CrossRef]
- Robacker, D.C.; Lauzon, C.R. Purine metabolizing capability of Enterobacter agglomerans affects volatiles production and attractiveness to Mexican fruit fly. J. Chem. Ecol. 2002, 28, 1549–1562. [Google Scholar] [CrossRef]
- Deetae, P.; Bonnarme, P.; Spinnler, H.E.; Henlick, H. Production of volatile aroma compounds by bacterial strains isolated from different surface-ripened french cheeses. Appl. Microbiol. Biotechnol. 2007, 76, 1161–1171. [Google Scholar] [CrossRef]
- Leejeerajumnean, A.; Duckham, S.C.; Owens, J.D.; Ames, J.M. Volatile compounds in Bacillus-fermented soyabeans. J. Sci. Food Agric. 2001, 81, 525–529. [Google Scholar] [CrossRef]
- Rajini, K.S.; Aparna, P.; Sasikala, C.H.; Ramana, C.V. Microbial metabolism of pyrazines. Crit. Rev. Microbiol. 2011, 37, 99–112. [Google Scholar] [CrossRef]
- Beck, H.C.; Hansen, A.M.; Lauritsen, F.R. Novel pyrazine metabolites found in polymyxin biosynthesis by Paenibacillus polymyxa. FEMS Microbiol. Lett. 2003, 220, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Micetich, R.G.; Mac Donald, J.C. Biosynthesis of neoaspergillic acid and neohydroxyaspergillic acids. J. Biol. Chem. 1965, 240, 1692–1695. [Google Scholar] [PubMed]
- Mac Donald, J.C. Toxicity, analyses and production of aspergillic acid and its analogues. Can. J. Biochem. 1973, 51, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.D.; Kwak, M.K.; Sim, SG. Inhibition of cytochrome P450 2E1 expression by 2-(allylthio) pyrazine, a potential chemoprotective agent: Hepatoprotective effects. Biochem. Pharm. 1997, 53, 261–269. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Houston, W.M. Production of blue-fluorescent pyrazines by Aspergillus parasiticus. J. Food Sci. 1982, 47, 779–782. [Google Scholar] [CrossRef]
- Uffen, R.L.; Canale-Parola, E. Synthesis of pulcherriminic acid by Bacillus subtilis. J. Bacteriol. 1972, 111, 86–93. [Google Scholar] [PubMed]
- Cheng, T.B.; Reineccius, G.A.; Bjorklund, J.A.; Leete, E. Biosynthesis of 2-methoxy-3-isopropylpyrazine in Pseudomonas Perolens. J. Agric. Food Chem. 1991, 39, 1009–1101. [Google Scholar] [CrossRef]
- Gallois, G.; Grimont, P.A.D. Pyrazines responsible for the potatolike odour produced by some Serratia and Cedecea strains. Appl. Environ. Microbiol. 1985, 50, 1048–1051. [Google Scholar]
- Mattey, M.; Harle, E.M. Aerobic metabolism of pyrazine compounds by a Pseudomonas species. Biochem. Soc. Trans. 1976, 4, 492–494. [Google Scholar] [CrossRef] [Green Version]
- Rajini, K.S.; Sasikala, C.; Ramana, C.V. Reductive degradation of pyrazine 2-carboxylate by a newly isolated Stenotrophomonas sp. HCU1. Biodegradation 2010, 21, 801–813. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, J.Y.; Bi, L.J.; Zhou, Y.F.; Zhang, Z.P.; Zhang, C.G.; Zhang, Y.; Zhang, E.X. Characterization of Mycobacterium tuberculosis nicotinamidase/pyrazinamidase. FEBS J. 2008, 275, 753–762. [Google Scholar] [CrossRef]
- Zhang, Y.; Scorpio, A.; Nikaido, H.; Sun, Z. Role of acid pH and deficient efflux of pyrazinoic acid in unique susceptibility of Mycobacterium tuberculosis to pyrazinamide. J. Bacteriol. 1999, 181, 2044–2049. [Google Scholar] [PubMed]
- Zhang, Y.; Wade, M.M.; Scorpio, A.; Zhang, H.; Sun, J. Mode of action of pyrazinamide: Disruption of Mycobacterium tuberculosis membrane transport and energetics by pyrazinoic acid. J. Antimicrob. Chemother. 2003, 52, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Hillier, S.G.; Lathe, R. Terpenes, hormones and life: Isoprene rule revisited. J. Endocrinol. 2019, 242, 2. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Biosynthesis, accumulation and emission of carotenoids, α-tocopherol, plastoquinone, and isoprene in leaves under high photosynthetic irradiance. Photosynth. Res. 2007, 92, 163. [Google Scholar] [CrossRef]
- Villanueva, L.; Sinninghe Damsté, J.S.; Schouten, S. A re-evaluation of the archaeal membrane lipid biosynthetic pathway. Nat. Rev. Microbiol. 2014, 12, 438–448. [Google Scholar] [CrossRef] [Green Version]
- Lehninger, A.L.; Cox, M.M.; Nelson, D.L. Lehninger Principles of Biochemistry, 5th ed.; Freeman, W.H., Ed.; W.H. Freeman and Company: New York, NY, USA, 2008; ISBN 978-1429222631. [Google Scholar]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural products (secondary metabolites). In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Biologists: Rock-ville, MD, USA, 2000; pp. 1250–1268. [Google Scholar]
- Chappell, J. The genetics and molecular genetics of terpene and sterol origami. Curr. Opin. Plant Biol. 2002, 5, 151–157. [Google Scholar] [CrossRef]
- Mumm, R.; Posthumusm, A.; Dicke, M. Significance of terpenoids in induced indirect plant defence against herbivorous arthropods. Plant Cell Environ. 2008, 31, 575–585. [Google Scholar] [CrossRef]
- Eaton, R.W.; Sandusky, P. Biotransformations of (+/−)-geosmin by terpene-degrading bacteria. Biodegradation 2010, 21, 71. [Google Scholar] [CrossRef]
- Morrone, D.; Chambers, J.; Lowry, L.; Kim, G.; Anterola, A.; Bender, K.; Peters, R.J. Gibberellin biosynthesis in bacteria: Separate ent-copalyl diphosphate and ent-kaurene synthases in Bradyrhizobium japonicum. FEBS Lett. 2009, 583, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [Green Version]
- Lombard, J.; Moreira, D. Origins and Early Evolution of the Mevalonate Pathway of Isoprenoid Biosynthesis in the Three Domains of Life. Mol. Biol. Evol. 2011, 28, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, C.J. Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 2003, 19, 415–417. [Google Scholar] [CrossRef]
- Zhao, B.; Lei, L.; Vassylyev, D.G.; Lin, X.; Cane, D.E.; Kelly, S.L.; Yuan, H.; Lamb, D.C.; Waterman, M.R. Crystal structure of albaflavenone monooxygenase containing a moonlighting terpene synthase active site. J. Biol. Chem. 2009, 284, 36711–36719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Voynova, N.E.; Rios, S.E.; Miziorko, H.M. Staphylococcus aureus mevalonate kinase: Isolation and characterization of an enzyme of the isoprenoid biosynthetic pathway. J. Bacteriol. 2004, 186, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Wilding, E.I.; Brown, J.R.; Brayant, A.P.; Chalker, A.F.; Holmes, D.J.; Ingrahm, K.A.; Iordanescu, S.; So, C.Y.; Rosenberg, M.; Gwynn, M.N. Identification, Evolution, and Essentiality of the Mevalonate Pathway for Isopentenyl Diphosphate Biosynthesis in Gram-Positive Cocci. J. Bacteriol. 2000, 182, 4319–4327. [Google Scholar] [CrossRef] [Green Version]
- Rodrìguez-Concepcion, M.; Boronat, A. Elucidation of the Methylerythritol Phosphate Pathway for Isoprenoid Biosynthesis in Bacteria and Plastids. A Metabolic Milestone Achieved through Genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, F.S.; Blanchard, J.L.; Cherkasov, A.; Av-Gay, Y.; Brunham, R.C.; Fernandez, R.C.; Finlay, B.B.; Otto, S.P.; Ouellette, B.F.; Keeling, P.J.; et al. Evidence that plant-like genes in Chlamydia species reflect an ancestral relationship between Chlamydiaceae, cyanobacteria, and the chloroplast. Genome Res. 2002, 12, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Rohdich, F.; Kis, K.; Bacher, A.; Eisenreich, W. The non-mevalonate pathway of isoprenoids: Genes, enzymes and intermediates. Curr. Opin. Chem. Biol. 2001, 5, 535–540. [Google Scholar] [CrossRef]
- Sprenger, G.A.; Schorken, U.; Wiegert, T.; Grolle, S.; de Graaf, A.A.; Taylor, S.V.; Begley, T.P.; Bringer-Meyer, S.; Sahm, H. Identification of a thiamin-dependent synthase in Escherichia coli required for the formation of the 1-deoxy-D-xylulose 5-phosphate precursor to isoprenoids, thiamin, and pyridoxol. Proc. Natl. Acad. Sci. USA 1997, 94, 12857–12862. [Google Scholar] [CrossRef] [Green Version]
- Lois, L.M.; Campos, N.; Rosa Putra, S.; Danielsen, K.; Rohmer, M.; Boronat, A. Cloning and characterization of a gene from Escherichia coli encoding a transketolase-like enzyme that catalyzes the synthesis of D-1-deoxyxylulose 5-phosphate, a common precursor for isoprenoid, thia- min, and pyridoxol biosynthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 2105–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzuyama, T.; Takahashi, S.; Watanabe, H.; Seto, H. Direct formation of 2-C-methyl-D-erythritol 4-phosphate from 1-deoxy-D-xylulose 5-phosphate by 1-deoxy-D-xylulose 5-phosphate reductoisomerase, a new enzyme in the non-mevalonate pathway to isopentenyl diphosphate. Tetrahedron Lett. 1998, 39, 4509–4512. [Google Scholar] [CrossRef]
- Takahashi, S.; Kuzuyama, T.; Watanabe, H.; Seto, H. A 1-deoxy-D- xylulose 5-phosphate reductoisomerase catalyzing the formation of 2-C- methyl-D-erythritol 4-phosphate in an alternative nonmevalonate pathway for terpenoid biosynthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 9879–9884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohdich, F.; Wungsintaweekul, J.; Fellermeier, M.; Sagner, S.; Herz, S.; Kis, K.; Eisenreich, W.; Bacher, A.; Zenk, M.H. Cytidine 5′-triphosphate- dependent biosynthesis of isoprenoids: YgbP protein of Escherichia coli catalyzes the formation of 4-diphosphocytidyl-2-C-methylerythritol. Proc. Natl. Acad. Sci. USA 1999, 96, 11758–11763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luttgen, H.; Rohdich, F.; Herz, S.; Wungsintaweekul, J.; Hecht, S.; Schuhr, C.A.; Fellermeier, M.; Sagner, S.; Zenk, M.H.; Bacher, A.; et al. Biosynthesis of terpenoids: YchB protein of Escherichia coli phosphorylates the 2-hydroxy group of 4-diphosphocytidyl-2-C-methyl-D-erythritol. Proc. Natl. Acad. Sci. USA 2000, 97, 1062–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herz, S.; Wungsintaweekul, J.; Schuhr, C.A.; Hecht, S.; Luttgen, H.; Sagner, S.; Fellermeier, M.; Eisenreich, W.; Zenk, M.H.; Bacher, A.; et al. Biosynthesis of terpenoids: YgbB protein converts 4-diphosphocytidyl- 2C-methyl-D-erythritol 2-phosphate to 2C-methyl-D-erythritol 2,4- cyclodiphosphate. Proc. Natl. Acad. Sci. USA 2000, 97, 2486–2490. [Google Scholar] [CrossRef] [Green Version]
- Rohdich, F.; Hecht, S.; Gartner, K.; Adam, P.; Krieger, C.; Amslinger, S.; Arigoni, D.; Bacher, A.; Eisenreich, W. Studies on the nonmevalonate terpene biosynthetic pathway: Metabolic role of IspH (LytB) protein. Proc. Natl. Acad. Sci. USA 2002, 99, 1158–1163. [Google Scholar] [CrossRef] [Green Version]
- Hahn, F.M.; Hurlburt, A.P.; Poulter, C.D. Escherichia coli open reading frame 696 is idi, a nonessential gene encoding isopentenyl diphosphate isomerase. J. Bacteriol. 1999, 181, 4499–4504. [Google Scholar]
- Nakano, C.; Kim, H.K.; Ohnishi, Y. Identification of the first bacterial monoterpene cyclase, a 1,8-cineole synthase, that catalyzes the direct conversion of geranyl diphosphate. ChemBioChem 2011, 5, 1988–1991. [Google Scholar] [CrossRef]
- Köksal, M.; Chou, W.K.; Cane, D.E.; Christianson, D.W. Unexpected reactivity of 2-fluorolinalyl diphosphate in the active site of crystalline 2-methylisoborneol synthase. Biochemistry 2013, 6, 5247–5255. [Google Scholar] [CrossRef] [Green Version]
- Nakano, C.; Kim, H.K.; Ohnishi, Y. Identification and characterization of the linalool/nerolidol synthase from Streptomyces clavuligerus. ChemBioChem 2011, 4, 2403–2407. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S.; Pahirulzaman, K.A.; Rabe, P.; Klapschinski, T.A. An improved technique for the rapid chemical characterisation of bacterial terpene cyclases. ChemBioChem 2014, 14, 810–814. [Google Scholar] [CrossRef]
- Brown, W.H.; Foote, C.S.; Iverson, B.L.; Anslyn, E.V. Organic Chemistry, 5th ed.; Harcourt College Publishers: Austin, TX, USA, 2007; ISBN 978-0981519432. [Google Scholar]
- Koe, B.K.; Sobin, B.A.; Celmer, W.D. PA 132, a new antibiotic. I. Isolation and chemical properties. Antibiot. Annu. 1956, 672, 5. [Google Scholar]
- Cane, D.E.; Watt, R.M. Expression and mechanistic analysis of a germacradienol synthase from Streptomyces coelicolor implicated in geosmin biosynthesis. Proc. Natl. Acad. Sci. USA 2003, 18, 1547–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagrain, B.; Martens, J.; Roelants, I.; Wenseleers, T. Method for Controlling Ant Colonies. Worldwide Application WO2015081390A3, 11 June 2015. [Google Scholar]
- Kim, S.Y.; Zhao, P.; Igarashi, M.; Sawa, R.; Tomita, T.; Nishiyama, M.; Kuzuyama, T. Cloning and heterologous expression of the cyclooctatin biosynthetic gene cluster afford a diterpene cyclase and two p450 hydroxylases. Chem. Biol. 2009, 16, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorner, C.; Hauslein, I.; Schrepfer, P.; Eisenreich, W.; Bruck, T. Targeted Engineering of Cyclooctat-9-en-7-ol Synthase: A Stereospecific Access to Two New Non-natural Fusicoccane-Type Diterpenes. ChemCatChem 2013, 5, 3289. [Google Scholar] [CrossRef]
- Dairi, T.; Hamano, Y.; Kuzuyama, T.; Itoh, N.; Furihata, K.; Seto, H. Eubacterial diterpene cyclase genes essential for production of the isoprenoid antibiotic terpentecin. J. Bacteriol. 2001, 183, 6085. [Google Scholar] [CrossRef] [Green Version]
- Hamano, Y.; Kuzuyama, T.; Itoh, N.; Furihata, K.; Seto, H.; Dairi, T. Functional analysis of eubacterial diterpene cyclases responsible for biosynthesis of a diterpene antibiotic, terpentecin. J. Biol. Chem. 2002, 277, 37098. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Lu, X. Microbial Synthesis of Alka(e)nes. Front. Bioeng. Biotechnol. 2013, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, A.; Rude, M.A.; Li, X.Z.; Popova, E.; del Cardayre, S.B. Microbial biosynthesis of alkanes. Science 2010, 329, 559–562. [Google Scholar] [CrossRef]
- Li, N.; Chang, W.C.; Warui, D.M.; Booker, S.J.; Krebs, C.; Bollinger, J.M. Evidence for only oxygenative cleavage of aldehydes to alk(a/e)nes and formate by cyanobacterial aldehyde decarbonylases. Biochemistry 2012, 51, 7908–7916. [Google Scholar] [CrossRef] [PubMed]
- Beller, H.R.; Goh, E.B.; Keasling, J.D. Genes involved in long- chain alkene biosynthesis in Micrococcus luteus. Appl. Environ. Microbiol. 2010, 76, 1212–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rude, M.A.; Baron, T.S.; Brubaker, S.; Alibhai, M.; Del Cardayre, S.B.; Schirmer, A. Terminal olefin (1-alkene) biosynthesis by a novel P450 fatty acid decarboxylase from Jeotgalicoccus species. Appl. Environ. Microbiol. 2011, 77, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez-Perez, D.; Begemann, M.B.; Pfleger, B.F. Modular synthase-encoding gene involved in alpha-olefin biosynthesis in Synechococcus sp strain PCC 7002. Appl. Environ. Microbiol. 2011, 77, 4264–4267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, M.K.; Turner, N.J.; Jones, P.R. Carboxylic acid reductase is a versatile enzyme for the conversion of fatty acids into fuels and chemical commodities. Proc. Natl. Acad. Sci. USA 2013, 110, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, T.P.; Middelhaufe, S.; Moore, K.; Edner, C.; Kolak, D.M.; Taylor, G.N. Synthe- sis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 7636–7641. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; Le Goff, G.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, G.J.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatile-mediated interactions between phylogenetically different soil bacteria. Front. Microbiol. 2014, 5, 289. [Google Scholar] [CrossRef] [Green Version]
- Piechulla, B.; Lemfack, M.C.; Kai, M. Effects of discrete bioactive microbial volatiles on plants and fungi. Plant Cell Environ. 2017, 40, 2042–2067. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Zweers, H.; de Boer, W.; Garbeva, P. A fragrant neighborhood: Volatile mediated bacterial interactions in soil. Front. Microbiol. 2015, 6, 1212. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyc, O.; Wolf, AB.; Garbeva, P. The effect of phylogenetically different bacteria on the fitness of Pseudomonas fluorescens in sand microcosms. PLoS ONE 2015, 10, e0119838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyc, O.; de Jager, V.C.L.; van den Berg, M.; Gerards, S.; Janssens, T.K.S.; Zaagman, N.; Kai, M.; Svatos, A.; Zweers, H.; Hordijk, C.; et al. Exploring bacterial interspecific interactions for discovery of novel antimicrobial compounds. Microb. Biotechnol. 2017, 10, 910–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Cordovez, V.; Etalo, D.W.; van der Voort, M.; Raaijmakers, J.M. Role of the GacS sensor kinase in the regulation of volatile production by plant growth-promoting Pseudomonas fluorescens SBW25. Front. Plant Sci. 2016, 7, 1706. [Google Scholar] [CrossRef] [PubMed]
- Lapouge, K.; Schubert, M.; Allain, F.H.; Haas, D. Gac/Rsm signal transduction pathway of gamma-proteobacteria: From RNA recognition to regulation of social behaviour. Mol. Microbiol. 2008, 67, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Heeb, S.; Haas, D. Regulatory roles of the GacS/GacA two-component system in plant-associated and other gram-negative bacteria. Mol. Plant Microb. Interact. 2001, 14, 1351–1363. [Google Scholar] [CrossRef] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calcagnile, M.; Tredici, S.M.; Talà, A.; Alifano, P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects 2019, 10, 441. https://doi.org/10.3390/insects10120441
Calcagnile M, Tredici SM, Talà A, Alifano P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects. 2019; 10(12):441. https://doi.org/10.3390/insects10120441
Chicago/Turabian StyleCalcagnile, Matteo, Salvatore Maurizio Tredici, Adelfia Talà, and Pietro Alifano. 2019. "Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants" Insects 10, no. 12: 441. https://doi.org/10.3390/insects10120441
APA StyleCalcagnile, M., Tredici, S. M., Talà, A., & Alifano, P. (2019). Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects, 10(12), 441. https://doi.org/10.3390/insects10120441