Effects of Detritivores on Nutrient Dynamics and Corn Biomass in Mesocosms

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study System
2.2. Experimental Design
2.3. Mesocosm Construction and Maintenance
2.4. Soil Collection and Preparation
2.5. Synthetic Rainwater and Storm Design
2.6. Invertebrate Assemblages
2.7. Soil Solution Nutrient Analysis
2.8. Mesocosm Harvest
2.9. Soil Nutrient Analysis
2.10. Corn and Invertebrate Nutrient Analysis
2.11. Statistical Analysis
3. Results
3.1. Invertebrate Mortality and Nutrients
3.2. Soil Nutrients and Ratios
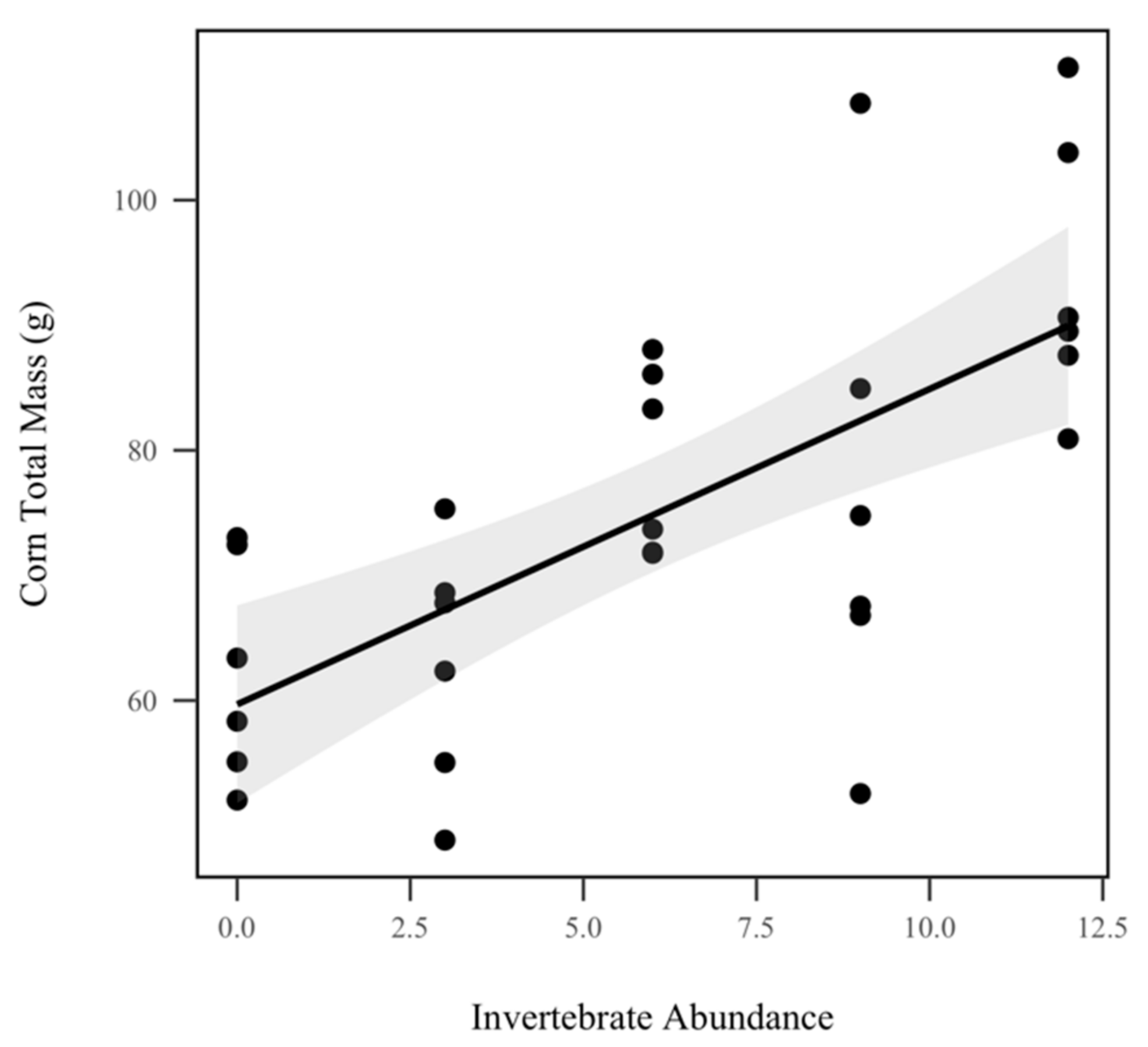
3.3. Corn Biomass and Nutrients
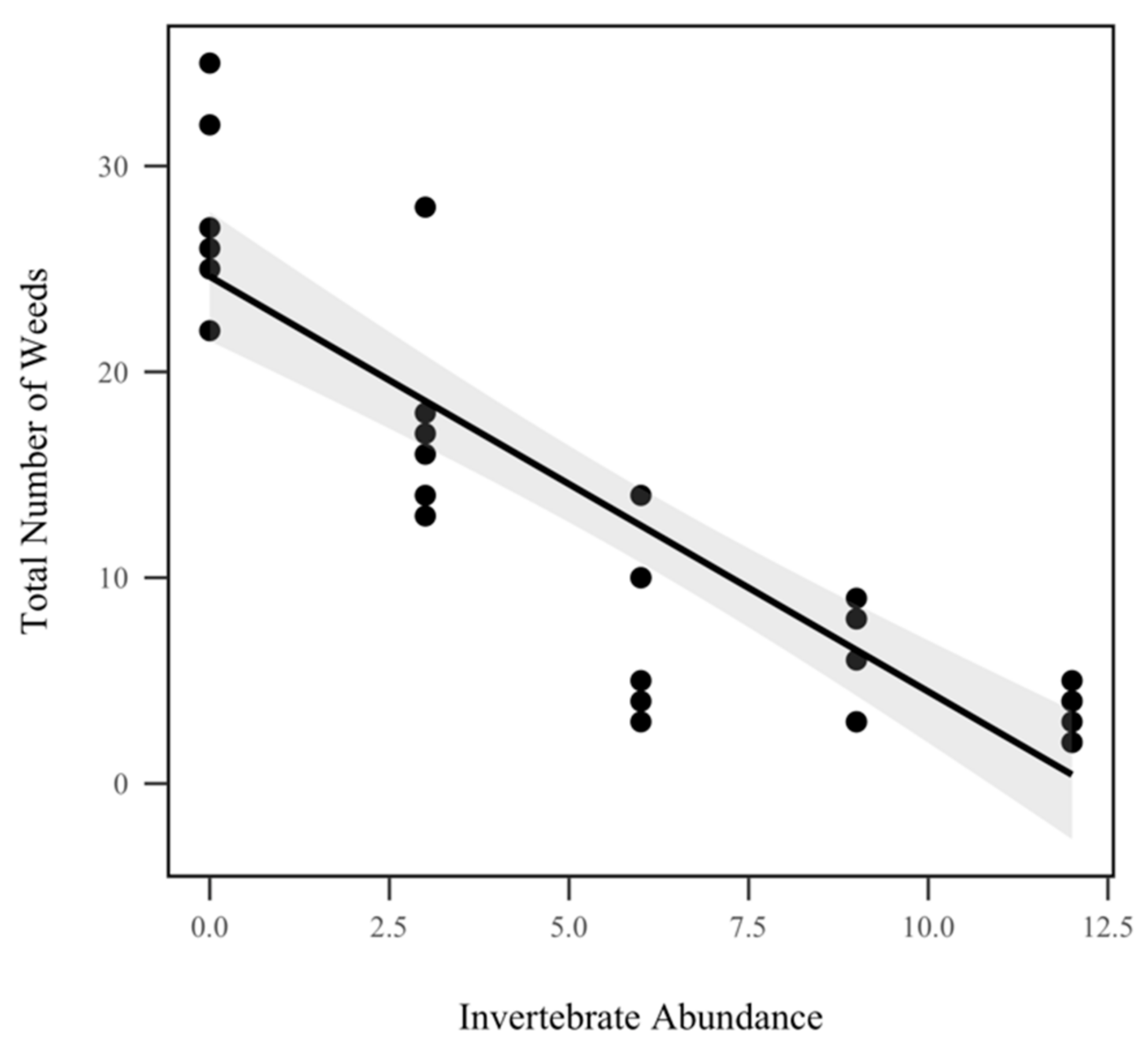
3.4. Weed Abundance
3.5. Soil Solution Volume and Nutrients
4. Discussion
4.1. Total Soil Nutrients
4.2. Soil Solution Nutrients
4.3. Weed Abundance
4.4. Corn Biomass
4.5. Future Work and Implications for Agriculture Management Strategies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
- Digestion Reagent Preparation:
- 1.
- Sodium Hydroxide 2.3 M
- a.
- Dissolve 92 g of sodium hydroxide in 800 mL of DI water in volumetric or graduated flask. CAUTION! When NaOH dissolves, heat is released so be careful handling this flask. Allow the resulting solution to cool and dilute to total volume of 1 L. Transfer reagent to plastic bottle. Solution will be stable at room temperature for 6 months.
- 2.
- Alkaline Persulfate Digestion Reagent (can adjust volume you prepare to volume needed by using same ratios persulfate and sodium hydroxide solutions to total volume)
- a.
- For 100 mL add 4.0 g of potassium persulfate and 10 mL of 2.3 M sodium hydroxide solution to 70 mL of DI water in a graduated flask. Mix with a magnetic stirrer until dissolution is complete, as seen in Table A1.
- b.
- Add enough DI water to bring volume to 100 mL. Swirl bottle to mix contents.
- c.
- Prepare this reagent same-day.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solution Volume (mL) | 2.3 M Sodium Hydroxide (mL) | DI Water (mL) | Potassium Persulfate (g) |
|---|---|---|---|
| 100 | 10 | 70 | 4 |
| 200 | 20 | 140 | 8 |
| 300 | 30 | 210 | 12 |
| 400 | 40 | 280 | 16 |
| 500 | 50 | 350 | 20 |
| 600 | 60 | 420 | 24 |
| 700 | 70 | 490 | 28 |
| 800 | 80 | 560 | 32 |
| 900 | 90 | 630 | 36 |
| 1000 | 100 | 700 | 40 |
- Sample Preparation:
- 1.
- Dispense sample and digestion reagent into Pyrex, round-bottom culture tubes at a ratio of 2:1 for sample to reagent for liquid samples (i.e., 10 mL of sample with 5 mL of digestion reagent) or a ratio of 5:1 for sample to reagent for solid samples (i.e., 100 mg of sample with 20 mL of digestion reagent).
- 2.
- Loosen cap on tube just a little before placing in autoclave.
- 3.
- Place capped tubes in autoclave and digest at 121 °C and 17 psi for 1 h. Follow proper autoclave instructions.
- 4.
- When digestion cycle is complete and pressure and temperature gages on the autoclave indicate 0 psi and less than 80 °C, remove alkaline persulfate digests from the autoclave and allow them to cool sufficiently.
- 5.
- Dilute digested samples with DI water if needed for SEAL AQ2 discrete analyzer. Make sure to weigh and record volumes of digested sample and DI water.
- 6.
- Digests can be stored for up to four days at room temperature if they are tightly capped.
- Analysis:
Appendix C
- TOC-LSM Soil Standard Preparation:
- 1.
- Bake 40 mL glass vials in the furnace at 600 °C for two hours to rid of any residual carbon.
- 2.
- Measure out five samples of the liquid standard to create a standard curve, as seen in Table A2.
- a.
- Liquid Standard: Nonpurgable Organic Carbon Standard (NPOC) R1848000 Ricca Chemical Company.
- i.
- 2000ppm +/− 5ppm Carbon.
| ID | Standard Target (mL) | Theoretical % Carbon (mg/L) |
|---|---|---|
| STD 1 | 0.0 | 0.0000 |
| STD 2 | 0.25 | 24.96 |
| STD 3 | 0.50 | 50.51 |
| STD 4 | 0.75 | 75.30 |
| STD 5 | 0.99 | 100.5 |
- TOC-LSM Quality Control:
- 1.
- Measure out target samples of NPOC into 40 mL glass vials.
- 2.
- Bring samples as close to 20 mL as possible by adding DI water to 40 mL glass vials.
- 3.
- If quality control measurement is not within 10% of theoretical carbon content, measure new standards and repeat Quality Control.
- TOC-LSM Sample Preparation:
- 1.
- Measured out 20.0 mL of all unknown samples into 40 mL glass vials.
- TOC-LSM Analysis:
- 1.
- Dissolved organic carbon was determined using high temperature oxidation followed by infrared detection of CO2 (Shimadzu TOC-VCSH) equipped with a liquid auto sampler (Shimadzu ASI-L).
- 2.
- Turn on the air/oxygen at 3 bars of pressure.
- 3.
- Turn on TOC-L and allow for 10 s start up.
- 4.
- Look at lights on the front for indications of machine status:
- a.
- (Red light = error (check gas))
- b.
- (Yellow light = warming up)
- c.
- (Green light = ready to use)
- d.
- (Blue light = measuring)
- 5.
- Turn on the Shimadzu SSM-5000A
- 6.
- Allow to fully heat to 900 °C. Once the Shimadzu TOC-L light is green, you’re able to start.
- TOC-L Sample Table Editor Software:
- 1.
- Using a zero shift, linear regression set up the calibration curve with the units as parts per million (PPM)
- 2.
- Set up the method settings for the Shimadzu TOC-L/ASI-L.
- a.
- Set to manual dilution 1×.
- b.
- Set determination by volume.
- c.
- Set measure in mg/L.
- d.
- Set to only 1 injection.
- 3.
- Hit “Connect” to connect the computer to the Shimadzu TOC-L/ASI-L.
- 4.
- Click “Start”.
- 5.
- Measure samples in the order of standards, quality control, and unknown samples.
- 6.
- With the ASI-L in the “Initial Position”, place the first sample in the ASI-L and lock tight.
- 7.
- Allow for a 2-minute purge.
- 8.
- Move ASI-L to the “measure” position.
- 9.
- After reading is complete, move to the “cooling” position and allow 30 s for cool down.
- 10.
- Move SSM-TC back to the “initial position”.
- 11.
- Repeat steps 6–10 for each subsequent sample.
Appendix D
- TOC-SSM Soil Standard Preparation:
- 1.
- Bake Shimadzu ceramic boats in the furnace at 600 °C for two hours to rid of any residual carbon.
- 2.
- Using a VWR A-Series balance, weigh out five samples of the standards to create a standard curve, as seen in Table A3, Table A4 and Table A5.
- a.
- Soil Standard: Leco soil certified reference material (LCRM)
- i.
- 3.82% +/− 0.07% Carbon
| ID | Target (mg) | Theoretical % Carbon (mg) |
|---|---|---|
| STD 1 | 0.00 | 0.0000 |
| STD 2 | 6.00 | 0.2292 |
| STD 3 | 12.00 | 0.4584 |
| STD 4 | 30.00 | 1.1460 |
| STD 5 | 60.00 | 2.2920 |
- b.
- Invertebrate Standard: Lab Grade Dextrose S25295A (Fisher Science Education)
| ID | Dextrose Sample (mg) | Carbon Concentration (mg) |
|---|---|---|
| STD 1 | 0.00 | 0.00 |
| STD 2 | 5.2 | 0.2080 |
| STD 3 | 15.3 | 0.6120 |
| STD 4 | 30.8 | 0.1232 |
| STD 5 | 64.6 | 0.2584 |
- c.
- Corn Standard: BBOT Leco Certified Reference Material (LCRM)
- 72.48% +/− 0.25% Carbon
| ID | Target (mg) | Theoretical Carbon (mg) |
|---|---|---|
| STD 1 | 0.00 | 0.000 |
| STD 2 | 5.00 | 3.624 |
| STD 3 | 13.00 | 9.422 |
| STD 4 | 34.00 | 24.643 |
| STD 5 | 60.00 | 43.488 |
- TOC-SSM Quality Control:
- 1.
- Weigh one 20 mg sample of LCRM into Shimadzu ceramic boat.
- 2.
- If quality control measurement is not within 10% of theoretical carbon content, measure new standards and repeat Quality Control.
- TOC-SSM Sample Preparation:
- 1.
- Using a VWR A-Series balance, weigh 20.0 mg of all unknown samples into Shimadzu ceramic boats.
- TOC-SSM Analysis:
- 1.
- Use a Shimadzu TOC-L/Shimadzu SSM-5000A, which uses high temperature oxidation combustion followed by CO2 infrared detection.
- 2.
- Turn on the air/oxygen at 3 bars of pressure.
- 3.
- Turn on TOC-L and allow for 10 s start up.
- 4.
- Look at lights on the front for indications of machine status:
- a.
- (Red light = error (check gas))
- b.
- (Yellow light = warming up)
- c.
- (Green light = ready to use)
- d.
- (Blue light = measuring)
- 5.
- Turn on the Shimadzu SSM-5000A
- 6.
- Allow to fully heat to 900 °C. Once the Shimadzu TOC-L light is green, you’re able to start.
- TOC-L Sample Table Editor Software:
- 1.
- Using a zero shift, linear regression set up the calibration curve with the units as parts per million (PPM).
- a.
- Set up the method settings for the Shimadzu TOC-L/SSM-5000A.
- b.
- Set to manual dilution 1×.
- c.
- Set determination by weight.
- d.
- Set measure in PPM if needed.
- e.
- Set to only 1 injection.
- 2.
- Hit “Connect” to connect the computer to the Shimadzu TOC-L/SSM-5000A.
- 3.
- Click “Start”.
- 4.
- Measure samples in the order of standards, quality control, and unknown samples.
- 5.
- With the SSM in the “Initial Position”, place the first sample in the SSM-TC and lock tight.
- 6.
- Allow for a 2-minute purge.
- 7.
- Move SSM-TC to the “measure” position.
- 8.
- After reading is complete, move to the “cooling” position and allow 30 s for cool down.
- 9.
- Move SSM-TC back to the “initial position”.
- 10.
- Repeat steps 6–10 for each subsequent sample.
References
- Richards, R.P.; Baker, D.B.; Crumrine, J.P. Improved water quality in Ohio tributaries to Lake Erie: A consequence of conservation practices. J. Soil Water Conserv. 2009, 64, 200–211. [Google Scholar] [CrossRef]
- Schindler, D.W.; Carpenter, S.R.; Chapra, S.C.; Hecky, R.E.; Orihel, D.M. Reducing Phosphorus to Curb Lake Eutrophication is a Success. Environ. Sci. Technol. 2016, 50, 8923–8929. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for Control of Anthropogenic Nitrogen and Phosphorus to Reduce Eutrophication of Inland Waters. Environ. Sci. Technol. 2011, 45, 10300–10305. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Bridgeman, T.B.; Bade, D.L. Nitrogen constrains the growth of late summer cyanobacterial blooms in Lake Erie. Adv. Microbiol. 2013, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Scott, J.T. Throwing Fuel on the Fire: Synergistic Effects of Excessive Nitrogen Inputs and Global Warming on Harmful Algal Blooms; ACS Publications: Washington, DC, USA, 2010. [Google Scholar]
- NOAA. Experimental Lake Erie Harmful Algal Bloom Bulletin #35; NOAA: Silver Spring, MD, USA, 2017; p. 1. [Google Scholar]
- Davis, T.W.; Harke, M.J.; Marcoval, M.A.; Goleski, J.; Orano-Dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Joosse, P.J.; Baker, D.B. Context for re-evaluating agricultural source phosphorus loadings to the Great Lakes. Can. J. Soil Sci. 2011, 91, 317–327. [Google Scholar] [CrossRef]
- Jarvie, H.P.; Sharpley, A.N.; Withers, P.J.A.; Scott, J.T.; Haggard, B.E.; Neal, C. Phosphorus Mitigation to Control River Eutrophication: Murky Waters, Inconvenient Truths, and “Postnormal” Science. J. Environ. Qual. 2013, 42, 295–304. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Edwards, C.A. Earthworm effects on N dynamics and soil respiration in microcosms receiving organic and inorganic nutrients. Soil Biol. Biochem. 1995, 27, 341–348. [Google Scholar] [CrossRef]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.-P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Crumsey, J.M.; Capowiez, Y.; Goodsitt, M.M.; Larson, S.; Moine, J.M.L.; Bird, J.A.; Kling, G.W.; Nadelhoffer, K.J. Exotic earthworm community composition interacts with soil texture to affect redistribution and retention of litter-derived C and N in northern temperate forest soils. Biogeochemistry 2015, 126, 379–395. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.; Ineson, P.; Heal, O.W.; Dhillion, S. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil Biol. 1997, 33, 159–193. [Google Scholar]
- Jouquet, P.; Dauber, J.; Lagerlöf, J.; Lavelle, P.; Lepage, M. Soil invertebrates as ecosystem engineers: Intended and accidental effects on soil and feedback loops. Appl. Soil Ecol. 2006, 32, 153–164. [Google Scholar] [CrossRef]
- Dempsey, M.A.; Fisk, M.C.; Yavitt, J.B.; Fahey, T.J.; Balser, T.C. Exotic earthworms alter soil microbial community composition and function. Soil Biol. Biochem. 2013, 67, 263–270. [Google Scholar] [CrossRef]
- Groffman, P.M.; Fahey, T.J.; Fisk, M.C.; Yavitt, J.B.; Sherman, R.E.; Bohlen, P.J.; Maerz, J.C. Earthworms increase soil microbial biomass carrying capacity and nitrogen retention in northern hardwood forests. Soil Biol. Biochem. 2015, 87, 51–58. [Google Scholar] [CrossRef]
- Jégou, D.; Schrader, S.; Diestel, H.; Cluzeau, D. Morphological, physical and biochemical characteristics of burrow walls formed by earthworms. Appl. Soil Ecol. 2001, 17, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Kautz, T. Research on subsoil biopores and their functions in organically managed soils: A review. Renew. Agric. Food Syst. 2015, 30, 318–327. [Google Scholar] [CrossRef]
- Bender, S.F.; van der Heijden, M.G.A. Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. J. Appl. Ecol. 2015, 52, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Blankinship, J.C.; Niklaus, P.A.; Hungate, B.A. A meta-analysis of responses of soil biota to global change. Oecologia 2011, 165, 553–565. [Google Scholar] [CrossRef]
- Briones, M.J.I.; Ineson, P.; Piearce, T.G. Effects of climate change on soil fauna; responses of enchytraeids, Diptera larvae and tardigrades in a transplant experiment. Appl. Soil Ecol. 1997, 6, 117–134. [Google Scholar] [CrossRef]
- Melillo, J.M.; Richmond, T.C.; Yohe, G.W. Ch. 2: Our Changing Climate. In Climate Change Impacts in the United States: The Third National Climate Assessment; National Climate Assessment Report: Washington, DC, USA, 2014; pp. 19–67. [Google Scholar]
- Vuorenmaa, J.; Rekolainen, S.; Lepistö, A.; Kenttämies, K.; Kauppila, P. Losses of Nitrogen and Phosphorus from Agricultural and Forest Areas in Finland during the 1980s and 1990s. Environ. Monit. Assess. 2002, 76, 213–248. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, R.W.; Dobb, D.E.; Raab, G.A.; Nocerino, J.M. Gy sampling theory in environmental studies. 1. Assessing soil splitting protocols. J. Chemom. 2002, 16, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Gibson, C.A.; O’Reilly, C.M.; Conine, A.L.; Jobs, W.; Belli, S. Organic matter carbon, nitrogen, and phosphorous from a single persulfate digestion. Limnol. Oceanogr. Methods 2015, 13, 202–211. [Google Scholar] [CrossRef]
- O’Dell, J. Method 365.1, Revision 2.0: Determination of Phosphorus by Semi-Automated Colorimetry; EPA: Washington, DA, USA, 1993. [Google Scholar]
- Nelson, N.S. An acid-persulfate digestion procedure for determination of phosphorus in sediments. Commun. Soil Sci. Plant Anal. 1987, 18, 359–369. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Luke, S.G. Evaluating significance in linear mixed-effects models in R. Behav. Res. Methods 2017, 49, 1494–1502. [Google Scholar] [CrossRef]
- Thakur, M.P.; Reich, P.B.; Hobbie, S.E.; Stefanski, A.; Rich, R.; Rice, K.E.; Eddy, W.C.; Eisenhauer, N. Reduced feeding activity of soil detritivores under warmer and drier conditions. Nat. Clim. Chang. 2018, 8, 75–78. [Google Scholar] [CrossRef]
- Joly, F.-X.; Weibel, A.K.; Coulis, M.; Throop, H.L. Rainfall frequency, not quantity, controls isopod effect on litter decomposition. Soil Biol. Biochem. 2019, 135, 154–162. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Cheng, W. Moisture modulates rhizosphere effects on C decomposition in two different soil types. Soil Biol. Biochem. 2007, 39, 2264–2274. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Kouno, K.; Wu, J.; Brookes, P.C. Turnover of biomass C and P in soil following incorporation of glucose or ryegrass. Soil Biol. Biochem. 2002, 34, 617–622. [Google Scholar] [CrossRef]
- Setälä, H.; Laakso, J.; Mikola, J.; Huhta, V. Functional diversity of decomposer organisms in relation to primary production. Appl. Soil Ecol. 1998, 9, 25–31. [Google Scholar] [CrossRef]
- O’Neill, R.V. Population Energies of the Millipede, Narceus Americanus (Beauvois). Ecology 1968, 49, 803–809. [Google Scholar] [CrossRef]
- Ausmus, B.S. Regulation of Wood Decomposition Rates by Arthropod and Annelid Populations. Ecol. Bull. 1977, 25, 180–192. [Google Scholar]
- Coleman, D.C.; Callaham, M.A.; Crossley, D.A. Secondary Production: Activities of Heterotrophic Organisms—The Soil Fauna. In Fundamentals of Soil Ecology, 3rd ed.; Coleman, D.C., Callaham, M.A., Crossley, D.A., Eds.; Academic Press: Cambridge, MA, USA, 2018; Chapter 4; pp. 77–171. ISBN 978-0-12-805251-8. [Google Scholar]
- Dudgeon, D.; Ma, H.H.T.; Lam, P.K.S. Differential palatability of leaf litter to four sympatric isopods in a Hong Kong forest. Oecologia 1990, 84, 398–403. [Google Scholar] [CrossRef]
- Hendriksen, N.B. Leaf litter selection by detritivore and geophagous earthworms. Biol. Fertil. Soils 1990, 10, 17–21. [Google Scholar]
- Ashwini, K.M.; Sridhar, K.R. Leaf litter preference and conversion by a saprophagous tropical pill millipede, Arthrosphaera magna Attems. Pedobiologia 2005, 49, 307–316. [Google Scholar] [CrossRef]
- Rincón, J.; Martínez, I. Food quality and feeding preferences of Phylloicus sp. (Trichoptera: Calamoceratidae). J. N. Am. Benthol. Soc. 2006, 25, 209–215. [Google Scholar] [CrossRef]
- Catalán, T.P.; Lardies, M.A.; Bozinovic, F. Food selection and nutritional ecology of woodlice in Central Chile. Physiol. Entomol. 2008, 33, 89–94. [Google Scholar] [CrossRef]
- Ihnen, K.; Zimmer, M. Selective consumption and digestion of litter microbes by Porcellio scaber (Isopoda: Oniscidea). Pedobiologia 2008, 51, 335–342. [Google Scholar] [CrossRef]
- Sarabi, V. Factors that influence the level of weed seed predation: A review. Weed Biol. Manag. 2019, 19, 61–74. [Google Scholar] [CrossRef]
- Warrell, D.A. 136—Injurious Arthropods. In Hunter’s Tropical Medicine and Emerging Infectious Disease, 9th ed.; Magill, A.J., Hill, D.R., Solomon, T., Ryan, E.T., Eds.; W.B. Saunders: London, UK, 2013; pp. 969–985. ISBN 978-1-4160-4390-4. [Google Scholar]
- Dettner, K. 4.09—Chemical Defense and Toxins of Lower Terrestrial and Freshwater Animals. In Comprehensive Natural Products II; Liu, H.-W.B., Mander, L., Eds.; Elsevier: Oxford, UK, 2010; pp. 387–410. ISBN 978-0-08-045382-8. [Google Scholar]
- Shear, W.A. The chemical defenses of millipedes (diplopoda): Biochemistry, physiology and ecology. Biochem. Syst. Ecol. 2015, 61, 78–117. [Google Scholar] [CrossRef] [Green Version]
- Oka, S. Mechanism of Antimicrobial Effect of Quinone Compounds. Agric. Biol. Chem. 1962, 26, 387–398. [Google Scholar]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2012, 186, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Chen, H.Y.H. Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nat. Clim. Chang. 2015, 5, 465–469. [Google Scholar] [CrossRef]
- Li, Y.; Niu, S.; Yu, G. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Glob. Chang. Biol. 2016, 22, 934–943. [Google Scholar] [CrossRef]
- Sardans, J.; Grau, O.; Chen, H.Y.H.; Janssens, I.A.; Ciais, P.; Piao, S.; Peñuelas, J. Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Chang. Biol. 2017, 23, 3849–3856. [Google Scholar] [CrossRef]
- Soons, M.B.; Hefting, M.M.; Dorland, E.; Lamers, L.P.M.; Versteeg, C.; Bobbink, R. Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biol. Conserv. 2017, 212, 390–397. [Google Scholar] [CrossRef]
- Yue, K.; Fornara, D.A.; Yang, W.; Peng, Y.; Li, Z.; Wu, F.; Peng, C. Effects of three global change drivers on terrestrial C: N: P stoichiometry: A global synthesis. Glob. Chang. Biol. 2017, 23, 2450–2463. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Sprent, J.I.; Raven, J.A.; Eady, P.E. Relationships between shoot to root ratio, growth and leaf soluble protein concentration of Pisum sativum, Phaseolus vulgaris and Triticum aestivum under different nutrient deficiencies. Plant Cell Environ. 2002, 22, 949–958. [Google Scholar] [CrossRef]
- de Groot, C.C.; Marcelis, L.F.M.; van den Boogaard, R.; Kaiser, W.M.; Lambers, H. Interaction of nitrogen and phosphorus nutrition in determining growth. Plant Soil 2003, 248, 257–268. [Google Scholar] [CrossRef]
- Güsewell, S.; Bollens, U. Composition of plant species mixtures grown at various N:P ratios and levels of nutrient supply. Basic Appl. Ecol. 2003, 4, 453–466. [Google Scholar]
- Güsewell, S. Responses of wetland graminoids to the relative supply of nitrogen and phosphorus. Plant Ecol. 2005, 176, 35–55. [Google Scholar] [CrossRef]
- Barberis, E.; Ajmone Marsan, F.; Scalenghe, R.; Lammers, A.; Schwertmann, U.; Edwards, A.C.; Maguire, R.; Wilson, M.J.; Delgado, A.; Torrent, J. European soils overfertilized with phosphorus: Part 1. Basic properties. Fertil. Res. 1995, 45, 199–207. [Google Scholar] [CrossRef]
- Balota, E.L.; Colozzi Filho, A.; Andrade, D.S.; Dick, R.P. Long-term tillage and crop rotation effects on microbial biomass and C and N mineralization in a Brazilian Oxisol. Soil Tillage Res. 2004, 77, 137–145. [Google Scholar] [CrossRef]
- Clapperton, M.J. Increasing soil biodiversity through conservation agriculture: Managing the soil as a habitat. In Proceedings of the II World Congress on Sustainable Agriculture: Biomass an in Harmony with Nature, Iguaçu, Brazil, 10–15 August 2003. [Google Scholar]








| χ2 of Predictor Variables | ||||
|---|---|---|---|---|
| Response Variables | Invertebrate Abundance | Rainfall Treatment | Invertebrate Abundance and Rainfall Interaction | |
| Mesocosm n = 30 | Ending Invertebrate Abundance | 54.5 * | 0 | 1.1 |
| Total Weed Abundance | 186.6 * | 0.2 | 1.5 | |
| Corn n = 30 | Aboveground Biomass | 21.9 * | 0.6 | 0.03 |
| Belowground Biomass | 13.0 * | 0.5 | 0.09 | |
| Corn Ear Biomass | 1.6 | 1.3 | 0.03 | |
| Total Biomass | 24.3 * | 0.6 | 0.1 | |
| Corn Nutrient Concentrations n = 30 | Total P | 0.5 | 0.02 | 0.4 |
| Total N | 0.6 | 0.01 | 0.02 | |
| Total C | −0.01 | 0.8 | 0.06 | |
| Corn Mass Balance Nutrients n = 30 | Total P | 16.1 * | 0.3 | 0.4 |
| Total N | 9.1 * | 0.2 | 0.0004 | |
| Total C | 14.7 * | 0.1 | 0.03 | |
| Soil n = 30 | Total P | 1.7 | 5.3 * | 9.6 * |
| Total N | 0.5 | 3.7 | 12.6 * | |
| Total C | 4.3 * | 0.008 | 6.0 * | |
| C:P | 5.7 * | 0.8 | 0.7 | |
| C:N | 5.3 * | 0.6 | 0.4 | |
| N:P | 0.009 | 0.002 | 0.01 | |
| Millipedes n = 4 | Total P | 0.02 | 0.9 | - |
| Total N | 4.8 * | 0.22 | - | |
| Total C | 0.9 | 0.9 | - | |
| Pill bugs n = 4 | Total P | 4.49 *,1 | 8.37 | - |
| Total N | 0.3 | 1.1 | - | |
| Total C | 4.2 *,1 | 0.7 | - | |
| Soil Solution Nutrient Concentrations n = 30 | Total P | 1.4 | 0.2 | 1.2 |
| Total N | 0.3 | 0.1 | 2.5 | |
| Total Organic C | 0.2 | 1.3 | 2.6 | |
| Total NH4+ | 0.0004 | 0.09 | 0.7 | |
| Total PO43− | 1.7 | 0.3 | 1.5 | |
| Total NO3+/NO2− | 0.7 | 0.8 | 1.4 | |
| Volume | 20.9 * | 3.4 | 0.006 | |
| Soil Solution Mass Balance Nutrients n = 30 | Total N | 7.3 * | 3.7 | 0.02 |
| Total P | 0.004 | 0.8 | 0.07 | |
| Total Organic C | 4.2 *,1 | 0.9 | 0.3 | |
| Total NH4+ | 1.1 | 2.7 | 0.6 | |
| Total PO43− | 0.09 | 0.2 | 0.05 | |
| Total NO3+/NO2− | 0.02 | 0.1 | 0.1 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindsey-Robbins, J.; Vázquez-Ortega, A.; McCluney, K.; Pelini, S. Effects of Detritivores on Nutrient Dynamics and Corn Biomass in Mesocosms. Insects 2019, 10, 453. https://doi.org/10.3390/insects10120453
Lindsey-Robbins J, Vázquez-Ortega A, McCluney K, Pelini S. Effects of Detritivores on Nutrient Dynamics and Corn Biomass in Mesocosms. Insects. 2019; 10(12):453. https://doi.org/10.3390/insects10120453
Chicago/Turabian StyleLindsey-Robbins, Josephine, Angélica Vázquez-Ortega, Kevin McCluney, and Shannon Pelini. 2019. "Effects of Detritivores on Nutrient Dynamics and Corn Biomass in Mesocosms" Insects 10, no. 12: 453. https://doi.org/10.3390/insects10120453
APA StyleLindsey-Robbins, J., Vázquez-Ortega, A., McCluney, K., & Pelini, S. (2019). Effects of Detritivores on Nutrient Dynamics and Corn Biomass in Mesocosms. Insects, 10(12), 453. https://doi.org/10.3390/insects10120453