Can Anastatus bifasciatus Be Used for Augmentative Biological Control of the Brown Marmorated Stink Bug in Fruit Orchards?

 ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Parasitoid Rearing
2.2. Release Sites
2.3. Egg Exposure
2.3.1. Parasitoid Efficacy and Persistence
2.3.2. Non-target parasitism
2.4. Parasitoid Releases
2.5. Treatment of Recollected Eggs
2.6. Statistical Analyses
3. Results
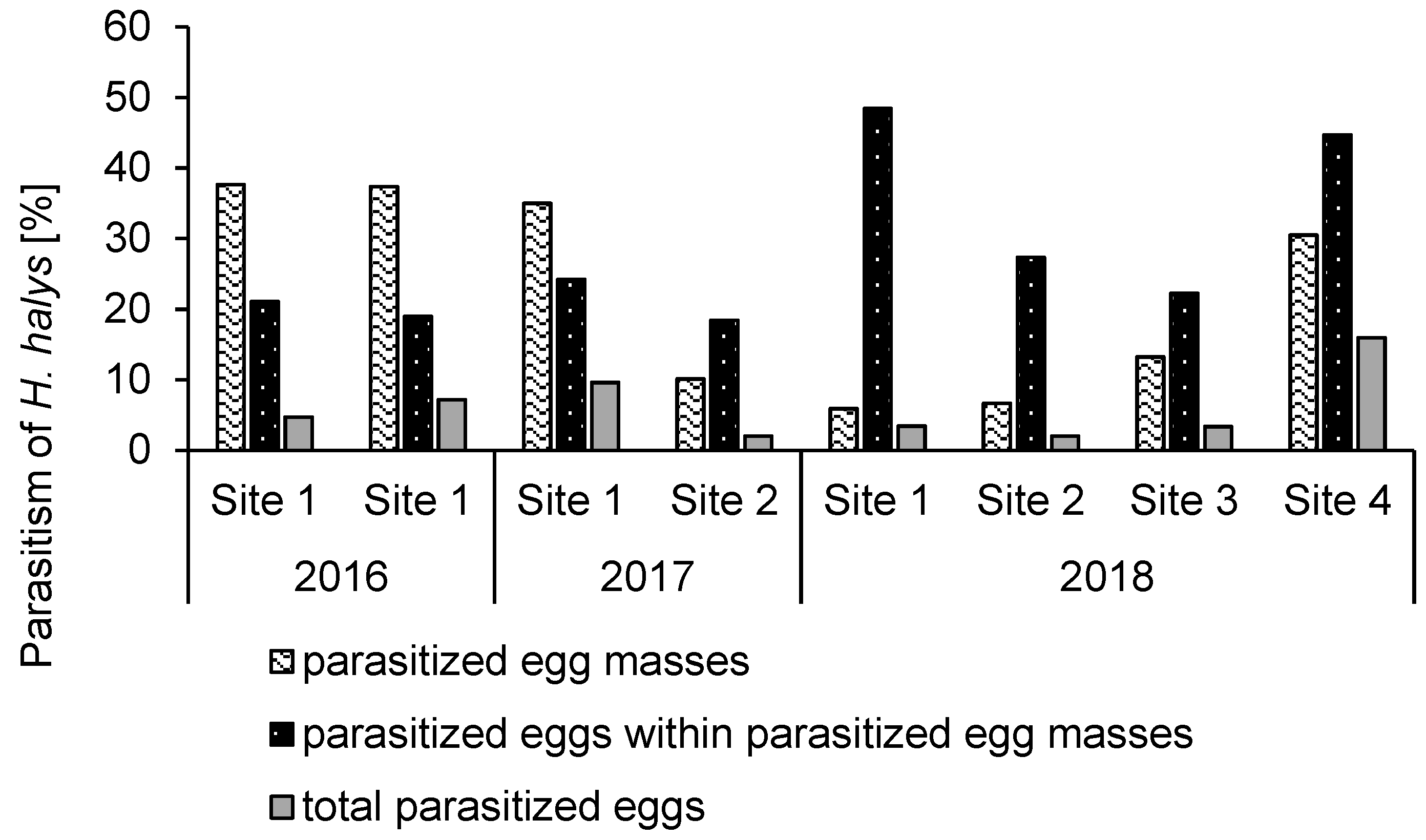
3.1. Parasitoid Efficacy and Persistence
3.2. Non-target Parasitism
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bigler, F.; Wajnberg, E.; Hassan, S.A. Quality control in Trichogramma production. In Biological Control with Egg Parasitoids; CAB International: Wallingford, UK, 1994. [Google Scholar]
- Smith, S.M. Biological control with Trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef] [PubMed]
- Narasimham, A.U.; Sankaran, T. Biology of Anastatus umae (Hymenoptera: Eupelmidae), an oothecal parasite of Neostylopyga rhombifolia. Colemania 1982, 1, 135–140. [Google Scholar]
- Jones, W.A. World review of the parasitoids of the southern green stink bug, Nezara viridula (L.)(Heteroptera: Pentatomidae). Annu. Entomol. Soc. Am. 1988, 81, 262–273. [Google Scholar] [CrossRef]
- Askew, A. A new European species of Anastatus Motschulsky (Hym., Eupelmidae). Entomol. Mag. 2005, 141, 1697–1699. [Google Scholar]
- Fay, H.; De Faveri, S. Egg parasitoids of fruitspotting bugs (Amblypelta spp.): Potential for their mass-rearing and strategic release. In Final Report HG308; Queensland Department of Primary Industries: Mareeba, Australia, 1997; p. 23. [Google Scholar]
- Huwer, R.; Maddox, C.D.; Fay, H.; Llewellyn, R.; Danne, A.; Hickey, M.; Melville, P.; Burnett, J.; Allen, A.; Groves, I. A multi target approach to fruitspotting bug management. In Proceedings of the VII World Avocado Congress 2011, Cairns, Australia, 5–9 September 2011. Research agreement 2011, MT10049:50. [Google Scholar]
- Govender, A.W. Australian Fruitspotting Bugs, Amblypelta nitida Stål and A. lutescens lutescens Distant (Hemiptera: Coreidae), and the Potential for Their Biologically Based Management in Macadamia Orchards. Ph.D. Thesis, University of Queensland, Brisbane, Australia, 2015. [Google Scholar]
- Shrestha, R. Productivity Improvement of Citrus Fruits through Effective Fruit Drop Management Technique in the Mid and Far Western Development Region of Nepal; Technical Report; National Citrus Research Program: Dhankuta, Nepal, 2011; p. 7. [Google Scholar]
- Chen, Y.; Lin, L.; Chen, J. Preliminary report of study on the release of Anastatus for the control of the stink bug (Tessaratoma papillosa). Fujian Agric. Sci. Technol. 1990, 2, 15–16. [Google Scholar]
- He, J.; Guo, L.; Tang, F.; Zhang, F.; Li, F.; Zhao, Z. The study of control Tessaratoma papillosa by releasing Anastatus japonicus. Guangxi Zhiwu 2001, 21, 163–165. [Google Scholar]
- Huang, M.; Mai, S.; Wu, W.; Poo, C. The bionomics of Anastatus sp. and its utilization for the control of lichee stink bug Tessaratoma papillosa Drury. Acta Entomol. Sin. 1974, 17, 362–375. [Google Scholar]
- Hou, Z.; Liang, H.; Chen, Q.; Hu, Y.; Tian, H. Application of Anastatus sp. against Halyomorpha halys. For. Pests Dis. 2009, 4, 39. [Google Scholar]
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stǻl) (Heteroptera: Pentatomidae): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. [Google Scholar]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stal (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitt. Schweiz. Entomol. Ges. 2008, 81, 1–8. [Google Scholar]
- Faúndez, E.I.; Rider, D.A. The brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Chile. Arq. Entomolóxicos 2017, 17, 305–307. [Google Scholar]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef]
- Kobayashi, T. The developmental stages of some species of the Japanese Pentatomoidea (Hemiptera): XVI. Homalogonia and an allied genus of Japan (Pentatomidae). Appl. Entomol. Zool. 1967, 2, 1–8. [Google Scholar]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Leskey, T.C.; Hamilton, G.C.; Nielsen, A.L.; Polk, D.F.; Rodriguez-Saona, C.; Bergh, J.C.; Herbert, D.A.; Kuhar, T.P.; Pfeiffer, D.; Dively, G.P.; et al. Pest status of the brown marmorated stink bug, Halyomorpha halys in the USA. Outlooks Pest Manag. 2012, 23, 218–226. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest Sci. 2017, 91, 661–670. [Google Scholar] [CrossRef]
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.P.; Nannini, R.; et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy. J. Pest Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Chabert, S.; Allemand, R.; Poyet, M.; Eslin, P.; Gibert, P. Ability of European parasitoids (Hymenoptera) to control a new invasive Asiatic pest, Drosophila suzukii. Biol. Control 2013, 63, 40–47. [Google Scholar] [CrossRef]
- Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Anfora, G. Augmentative releases of Trichopria drosophilae for the suppression of early season Drosophila suzukii populations. BioControl 2019, 64, 9–19. [Google Scholar] [CrossRef]
- Haye, T.; Fischer, S.; Zhang, J.; Gariepy, T. Can native egg parasitoids adopt the invasive brown marmorated stink bug, Halyomorpha halys (Heteroptera: Pentatomidae), in Europe? J. Pest Sci. 2015, 88, 693–705. [Google Scholar] [CrossRef]
- Roversi, P.F.; Marianelli, L.; Costi, E.; Maistrello, L.; Sabbatini, P.G. Searching for native egg-parasitoids of the invasive alien species Halyomorpha halys (Stål) (Heteroptera Pentatomidae) in Southern Europe. Redia 2016, 99, 63–70. [Google Scholar]
- Costi, E.; Haye, T.; Maistrello, L. Surveying native egg parasitoids and predators of the invasive Halyomorpha halys in Northern Italy. J. Appl. Entomol. 2019, 143, 299–307. [Google Scholar] [CrossRef]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef]
- Noyes, J.; Universal Chalcidoidea Database—World Wide Web Electronic Publication. Natural History Museum Update March. Available online: http://www.nhm.ac.uk/our-science/data/chalcidoids/ (accessed on 15 October 2018).
- Stahl, J.M.; Babendreier, D.; Haye, T. Using the egg parasitoid Anastatus bifasciatus against the invasive brown marmorated stink bug in Europe: Can non-target effects be ruled out? J. Pest Sci. 2018, 91, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Google Site 1, Canton of Zurich, Switzerland. Available online: http://maps.google.com (accessed on 15 December 2018).
- Morrison, W.R.; Lee, D.H.; Short, B.D.; Khrimian, A.; Leskey, T.C. Establishing the behavioral basis for an attract-and-kill strategy to manage the invasive Halyomorpha halys in apple orchards. J. Pest Sci. 2016, 89, 81–96. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 20 June 2015).
- RStudio Team. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA. 2016. Available online: http://www.rstudio.com/ (accessed on 20 June 2015).
- Komsta, L. Median-Based Linear Models, R Package Version; 2005. Available online: https://cran.r-project.org/web/packages/mblm/index.html (accessed on 20 July 2015).
- Stahl, J.M.; Babendreier, D.; Haye, T. Life history of Anastatus bifasciatus, a potential biological control agent of the brown marmorated stink bug in Europe. Biol. Control 2019. [Google Scholar] [CrossRef]
- Mills, N.J.; Wajnberg, E. Optimal foraging behavior and efficient biological control methods. In Behavioral Ecology of Insect Parasitoids: From Theoretical Approaches to Field Applications; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 3–30. [Google Scholar]
- Andow, D.A.; Prokrym, D.R. Release density, efficiency and disappearance of Trichogramma nubilale for control of European corn borer. Entomophaga 1991, 36, 105–113. [Google Scholar] [CrossRef]
- Vinson, S.B. Habitat location. In Semiochemicals: Their Role in Pest Control; Wiley: New York, NY, USA, 1981. [Google Scholar]
- Weseloh, R.M.; Anderson, J.F. Inundative release of Apanteles melanoscelus against the gypsy moth. Environ. Entomol. 1975, 4, 33–36. [Google Scholar] [CrossRef]
- Colazza, S.; Salerno, G.; Wajnberg, E. Volatile and contact chemicals released by Nezara viridula (Heteroptera: Pentatomidae) have a kairomonal effect on the egg parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). Biol. Control 1999, 16, 310–317. [Google Scholar] [CrossRef]
- Rondoni, G.; Bertoldi, V.; Malek, R.; Foti, M.C.; Peri, E.; Maistrello, L.; Haye, T.; Conti, E. Native egg parasitoids recorded from the invasive Halyomorpha halys successfully exploit volatiles emitted by the plant–herbivore complex. J. Pest Sci. 2017, 90, 1087–1095. [Google Scholar] [CrossRef]
- Os, N.; Varshney, R. Utilization of Trichogrammatid egg parasitoids in pest management. Acta Sci. Agric. 2018, 2, 49–53. [Google Scholar]
- Li, D.S.; Liao, C.; Zhang, B.X.; Song, Z.W. Biological control of insect pests in litchi orchards in China. Biol. Control 2014, 68, 23–36. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Kuhlmann, U.; Gillott, C.; Erlandson, M. A large-scale comparison of conventional and molecular methods for the evaluation of host–parasitoid associations in non-target risk-assessment studies. J. Appl. Ecol. 2008, 45, 708–715. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Haye, T.; Zhang, J. A molecular diagnostic tool for the preliminary assessment of host–parasitoid associations in biological control programmes for a new invasive pest. Mol. Ecol. 2014, 23, 3912–3924. [Google Scholar] [CrossRef]
- Stahl, J.M.; Gariepy, T.G.; Beukeboom, L.W.; Haye, T. A molecular tool to identify Anastatus parasitoids of the brown marmorated stink bug. Entomol. Exp. Appl. 2019; in press. [Google Scholar]
- Konopka, J.K.; Haye, T.; Gariepy, T.D.; McNeil, J.N. Possible coexistence of native and exotic parasitoids and their impact on control of Halyomorpha halys. J. Pest Sci. 2017, 90, 1119–1125. [Google Scholar] [CrossRef]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.; Shrewsbury, P.M. Sentinel eggs underestimate rates of parasitism of the exotic brown marmorated stink bug, Halyomorpha halys. Biol. Control 2014, 78, 61–66. [Google Scholar] [CrossRef]
- Risch, S.J.; Andow, D.; Altieri, M.A. Agroecosystem diversity and pest control: Data, tentative conclusions, and new research directions. Environ. Entomol. 1983, 12, 625–629. [Google Scholar] [CrossRef]
- Kozár, F. Organization of arthropod communities in agroecosystems. Acta Phytopathol. Entomol. Hung. 1992, 27, 365–373. [Google Scholar]
- Simon, S.; Bouvier, J.C.; Debras, J.F.; Sauphanor, B. Biodiversity and pest management in orchard systems. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2011; Volume 2, pp. 693–709. [Google Scholar]
- Rand, T.A.; Tylianakis, J.M.; Tscharntke, T. Spillover edge effects: The dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecol. Lett. 2006, 9, 603–614. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Location (Municipality, Canton/Region, Country) | GPS Coordinates | Management | Host Plants in Plot (Species, Variety) | H. halys Presence | Release Plot Size [m2] | Number of Releases |
|---|---|---|---|---|---|---|---|
| 1 | Lindau, Zurich, Switzerland | 47°26’52.0″N 8°40’47.6″E | IPM | Malus pumila, Golden Delicious/Diwa/Braeburn | no | 210 | 4 |
| 2 | Bellinzona, Ticino, Switzerland | 46°09’42.1″N 8°58’12.2″E | IPM | Malus pumila, Golden Delicious/Braeburn | yes | 288 | 2 |
| 3 | Manno, Ticino, Switzerland | 46°01’52.8″N 8°55’20.4″E | Organic | Malus pumila, unknown varieties | yes | 212 | 1 |
| 4 | Carpi, Emilia-Romagna, Italy | 44°43’46.8″N 10°52’30.0″E | Organic | Pyrus communis, Abate Fetel | yes | 480 | 1 |
| Exposure Date | Location | Treatment | Mean Temperature (Min – Max) [°C] | Egg Masses (Eggs) Exposed | Egg Masses (Eggs) Recovered 1 |
|---|---|---|---|---|---|
| 2016 | |||||
| 23.–28. Jul | 1 – Lindau | Pre-release monitoring | 22.4 (12.9–36.0) | 25 (657) | 22 (575) |
| 28. Jul – 2. Aug | Release | 21.7 (12.4–38.3) | 97 (2653) | 93 (2409) | |
| 11.–16. Aug | Post-release monitoring | 21.8 (9.6–36.6) | 55 (1480) | 54 (1459) | |
| 16.–21. Aug | Release | 20.4 (12.1–36.6) | 90 (2410) | 83 (2258) | |
| 2017 | |||||
| 17.–21. Jul | 1 – Lindau | Pre-release monitoring | 23.0 (13.8–34.5) | 25 (623) | 15 (298) |
| 29. Jul–3. Aug | Release | 23.2 (15.3–35.0) | 90 (2273) | 20 (479) | |
| 10.–17. Aug | Post-release monitoring | 18.8 (10.2–32.7) | 55 (1410) | 23 (562) | |
| 2.–10. Aug | 2 – Bellinzona | Pre-release monitoring | 23.4 (15.3–38.5) | 25 (637) | 20 (416) |
| 14.–21. Aug | Release | 23.1 (10.9–35.2) | 90 (2327) | 89 (1897) | |
| 28. Aug–4. Sep | Post-release monitoring | 21.2 (12.3–33.5) | 55 (1387) | 53 (774) | |
| 2018 | |||||
| 19.–25. Jul | 2 – Bellinzona | Pre-release monitoring | 25.0 (13.9–38.3) | 25 (685) | 22 (1371) |
| 25.–30. Jul | Release | 26.1 (15.5–37.9) | 90 (2418) | 90 (2205) | |
| 8.–13. Aug | Post-release monitoring | 23.7 (16.8–36.4) | 55 (2697) | 51 (1940) | |
| 19.–25. Jul | 4 – Carpi | Pre-release monitoring | 25.0 (16.0–35.0) | 25 (694) | 25 (363) |
| 25.–30. Jul | Release | 26.5 (17.0–36.5) | 90 (2392) | 82 (1780) | |
| 9.–14. Aug | Post-release monitoring | 25.4 (17.0–35.5) | 55 (1469) | 55 (958) | |
| 25.–30. Jul | 3 – Manno | Pre-release monitoring | 26.6 (15.3–44.5) | 25 (752) | 20 (545) |
| 30. Jul–3. Aug | Release | 28.0 (17.5–39.4) | 90 (2347) | 68 (957) | |
| 13.–17. Aug | Post-release monitoring | 23.1 (14.6–34.7) | 55 (1677) | 51 (1345) | |
| 6.–10. Aug | 1 – Lindau | Pre-release monitoring | Na | 25 (611) | 10 (58) |
| 11.–15. Aug | Release | Na | 90 (2303) | 85 (1026) | |
| 24.–28. Aug | Post-release monitoring | Na | 55 (1760) | 51 (1536) | |
| Total | 1287 (35,662) | 1082 (25,340) |
| Species | Site | Year | Egg Batch Size | Total # Eggs Exposed/Site |
|---|---|---|---|---|
| Samia cynthia (Drury) (Saturniidae) | 1 | 2016 | 6 | 42 |
| Euthrix potatoria (L.) (Lasiocampidae) | 1 | 2016 | 6 | 252 |
| Odonestis pruni (L.) (Lasiocampidae) | 1 | 2016 | 4 | 168 |
| Dendrolimus pini (L.) (Lasiocampidae) | 1, 2 | 2017 | 4 | 192 |
| Lasiocampa quercus (L.) (Lasiocampidae) | 1, 2, 3, 4 | 2018 | 6 (3 in site 4) | 288 (144) |
| Macrothylacia rubi (L.) (Lasiocampidae) | 4 | 2018 | 3 | 144 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stahl, J.M.; Babendreier, D.; Marazzi, C.; Caruso, S.; Costi, E.; Maistrello, L.; Haye, T. Can Anastatus bifasciatus Be Used for Augmentative Biological Control of the Brown Marmorated Stink Bug in Fruit Orchards? Insects 2019, 10, 108. https://doi.org/10.3390/insects10040108
Stahl JM, Babendreier D, Marazzi C, Caruso S, Costi E, Maistrello L, Haye T. Can Anastatus bifasciatus Be Used for Augmentative Biological Control of the Brown Marmorated Stink Bug in Fruit Orchards? Insects. 2019; 10(4):108. https://doi.org/10.3390/insects10040108
Chicago/Turabian StyleStahl, Judith M., Dirk Babendreier, Cristina Marazzi, Stefano Caruso, Elena Costi, Lara Maistrello, and Tim Haye. 2019. "Can Anastatus bifasciatus Be Used for Augmentative Biological Control of the Brown Marmorated Stink Bug in Fruit Orchards?" Insects 10, no. 4: 108. https://doi.org/10.3390/insects10040108
APA StyleStahl, J. M., Babendreier, D., Marazzi, C., Caruso, S., Costi, E., Maistrello, L., & Haye, T. (2019). Can Anastatus bifasciatus Be Used for Augmentative Biological Control of the Brown Marmorated Stink Bug in Fruit Orchards? Insects, 10(4), 108. https://doi.org/10.3390/insects10040108