Efficacy of Two Entomopathogenic Fungi, Metarhizium brunneum, Strain F52 Alone and Combined with Paranosema locustae against the Migratory Grasshopper, Melanoplus sanguinipes, under Laboratory and Greenhouse Conditions




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setting
2.2. Fungal and Baits Preparations
2.3. Treatments Application to the Hosts
2.4. Data Collection
2.5. Statistical Analysis
3. Results
3.1. Treatments Exposure in the Laboratory
3.1.1. Single Pathogen Treatments
3.1.2. Combination of Treatments
3.2. Treatments Exposure in the Greenhouse
3.3. Virulence
3.4. Interactions between the Pathogens
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sergeev, M.G. Ecogeographical distribution of Orthoptera. In The Bionomics of Grasshoppers, Katydids, and their Kin; Gangwere, S.K., Muralirangan, M.C., Muralirangan, M., Eds.; CAB International: Wallingford, UK, 1997; pp. 129–146. [Google Scholar]
- Branson, D.H.; Joern, A.; Sword, G.A. Sustainable management of insect herbivores in grassland ecosystems: New perspectives in grasshopper control. Bioscience 2006, 56, 743–755. [Google Scholar] [CrossRef]
- Pfadt, R.E. Field Guide to Common Western Grasshoppers; Wyoming Agricultural Experiment Station Bulletin 912: Laramie, WY, USA, 1988. [Google Scholar]
- Pfadt, R.E. A Field Guide to Common Western Grasshoppers, 3rd ed.; Wyoming Agricultural Experiment Station Bulletin 912: Laramie, WY, USA, 2002; p. 228. [Google Scholar]
- Lockwood, J.A.; McNary, T.J.; Larsen, J.C.; Zimmerman, K.; Shambaugh, B.A.; Latchininsky, A.V.; Herring, B.; Legg, C. Distribution Atlas for Grasshoppers and Mormon Crickets in Wyoming, 1988-92. Available online: http://www.uwyo.edu/entomology/grasshoppers/ghatlas.htm (accessed on 8 March 2019).
- Fielding, D.J.; Brusven, M.A.; Shafii, B.; Price, W.J. Spatial heterogeneity of low-density populations of Melanoplus sanguinipes (Orthoptera: Acrididae) associated with grazing and vegetation treatments. J. Can. Entomol. 2001, 133, 843–855. [Google Scholar] [CrossRef]
- Lockwood, J.A.; Anderson-Sprecher, R.; Schell, S.P. When less is more: Optimization of reduced agent-area treatments (RAATs) for management of rangeland grasshoppers. Crop Prot. 2002, 21, 551–562. [Google Scholar] [CrossRef]
- McNary, T.J.; Shambaugh, B.A.; Elliston, R.J.; Brown, C.L. Cooperative rangeland grasshopper suppression in Wyoming (USA) in 2010. Metaleptea 2011, 31, 10–12. [Google Scholar]
- USDA APHIS. Rangeland Grasshopper and Mormon Cricket Suppression Program. Final Environmental Impact Statement; United States Department of Agriculture, Marketing and Regulatory Programs: Riverdale, MD, USA, 2002; p. 336.
- Smith, D.I.; Lockwood, J.A.; Latchininsky, A.V.; Legg, D.E. Changes in non-target arthropod populations following application of liquid bait formulations of insecticides for control of rangeland grasshoppers. J. Int. Pest Manag. 2006, 52, 125–139. [Google Scholar] [CrossRef]
- Hajek, A.E. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2004; p. 378. [Google Scholar]
- Foster, R.N.; Jaronski, S.; Reuter, K.C.; Black, L.R.; Schlothauer, R.; Harper, J.; Jech, L.E. Simulated aerial sprays for field cage evaluation of Beauveria bassiana and Metarhizium brunneum (Ascomycetes: Hypocreales) against Anabrus simplex (Orthoptera: Tettigoniidae) in Montana. Biocontrol Sci. Technol. 2011, 21, 1331–1350. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Jackson, M.A. Efficacy of Metarhizium anisopliae microsclerotial granules. Biocontrol Sci. Technol. 2008, 18, 849–863. [Google Scholar] [CrossRef]
- Furlong, M.J.; Pell, J.K.; Reddy, G.V.P. Premortality effects of Zoophthora radicans infection in Plutella xylostella. J. Invertebr. Pathol. 1997, 70, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.V.P.; Furlong, M.J.; Pell, J.K.; Poppy, G.M. Zoophthora radicans infection inhibits the response to and production of sex pheromone in the diamondback moth (Note). J. Invertebr. Pathol. 1998, 72, 167–169. [Google Scholar] [CrossRef] [PubMed]
- USEPA. Reregistration Eligibility Document (RED) Facts: Nosema locustae; US Environmental Protection Agency, EPA: Washington, DC, USA, 1992; 738-F-92-011.
- Lockwood, J.A.; Bomar, C.R.; Ewen, A.B. The history of biological control with Nosema locustae: Lessons for locust management. J. Int. Tro. Insect Sci. 1999, 19, 333–350. [Google Scholar] [CrossRef]
- Lange, C.E. The host and geographical range of the grasshopper pathogen Paranosema (Nosema) locustae revisited. J. Orthoptera Res. 2005, 14, 137–141. [Google Scholar] [CrossRef]
- Solter, L.F.; Maddox, J.V. Timing of an early sporulation sequence of microsporidia in the Genus Vairimorpha (Microsporidia: Burenellidae). J. Invertebr. Pathol. 1998, 72, 323–329. [Google Scholar] [CrossRef]
- Milner, R.J.; Hunter, D.M. Recent developments in the use of fungi as biopesticides against locusts and grasshoppers in Australia. J. Orthoptera Ress. 2001, 10, 271–276. [Google Scholar] [CrossRef]
- Milner, R.J.; Lozano, L.B.; Driver, F.; Hunter, D. A comparative study of two Mexican isolates with an Australian isolate of Metarhizium anisopliae var. acridum–strain characterisation, temperature profile and virulence for wingless grasshopper, Phaulacridium vittatum. Biocontrol 2003, 48, 335–348. [Google Scholar] [CrossRef]
- Tounou, A.K.; Kooyman, C.; Douro-Kpindou, O.K.; Poehling, H.M. Interaction between Paranosema locustae and Metarhizium anisopliae var. acridum, two pathogens of the desert locust, Schistocerca gregaria under laboratory conditions. J. Invertebr. Pathol. 2008, 97, 203–210. [Google Scholar] [CrossRef]
- Tounou, A.K.; Kooyman, C.; Douro-Kpindou, O.K.; Poehling, H.M. Combined field efficacy of Paranosema locustae and Metarhizium anisopliae var. acridum for the control of sahelian grasshoppers. Biocontrol 2008, 53, 813–828. [Google Scholar] [CrossRef]
- Roxburgh, L.; Pinshow, B.; Prange, H.D. Temperature regulation by evaporative cooling in a desert grasshopper, Calliptamus barbarous. J. Therm. Biol. 1996, 21, 331–337. [Google Scholar] [CrossRef]
- Ouedraogo, R.M.; Cusson, M.; Goettel, M.S.; Brodeur, J. Inhibition of fungal growth in thermoregulating locusts, Locusta migratoria, infected by the fungus Metarhizium anisopliae var acridum. J. Invertebr. Pathol. 2003, 82, 103–109. [Google Scholar] [CrossRef]
- Ouedraogo, R.M.; Goettel, M.S.; Brodeur, J. Behavioral thermoregulation in the migratory locust: A therapy to overcome fungal infection. Oecologia 2004, 138, 312–319. [Google Scholar] [CrossRef]
- USEPA (United States Environmental Protection Administration). Nosema locustae (117001) Fact Sheet. 2000. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/fs_PC-117001_01-Oct-00.pdf (accessed on 13 February 2019).
- Hinks, C.F.; Erlandson, M.A. Rearing grasshoppers and locusts: Review, rationale and update. J. Orthoptera Res. 1994, 1, 1–10. [Google Scholar] [CrossRef]
- Elliot, S.L.; Blanford, S.; Thomas, M.B. Host–pathogen interactions in a varying environment: Temperature, behavioral fever and fitness. Proc. R. Soc. London B. 2002, 269, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill Book Company: New York, NY, USA, 1980. [Google Scholar]
- Milliken, G.A.; Johnson, D.E. Analysis of Messy Data; Chapman and Hall: New York, NY, USA, 1992. [Google Scholar]
- SAS Institute. SAS/STAT User’s Guide, 2nd ed.; Version 13.2 for Windows; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Nishimatsu, T.; Jackson, J.J. Interaction of insecticides, entomopathogenic nematodes, and larvae of the western corn root worm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1998, 91, 410–418. [Google Scholar] [CrossRef]
- Feinberg, S.E. The Analysis of Cross-Classified Categorical Data, 2nd ed.; The MIT Press: Cambridge, MA, USA, 1980; p. 198. [Google Scholar]
- Cox, F.E.G. Concomitant infections, parasites and immune responses. Parasitology 2001, 122, 23–38. [Google Scholar] [CrossRef]
- Hajek, A.E.; St. Leger, R.J. Interactions between fungal pathogens and insect hosts. Annu. Rev. Entomol. 1994, 39, 293–322. [Google Scholar] [CrossRef]
- Tounou, A.K.; Kooyman, C.; Douro-Kpindou, O.K.; Gumedzoe, Y.M.; Poehlingn, H.M. Laboratory assessment of the potential of Paranosema locustae to control immature stages of Schistocerca gregaria and Oedaleus senegalensis and vertical transmission of the pathogen in host populations. Biocontrol Sci. Technol. 2011, 21, 605–617. [Google Scholar] [CrossRef]
- Solter, L.F.; Becnel, J.J.; Oi, D.H. Microsporidian entomopathogens. In Insect Pathology, 2nd ed.; Vega, F.E., Kaya, H.K., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 221–263. [Google Scholar]
- Henry, J.E. Epizootiology of infections by Nosema locustae Canning (Microsporida: Nosematidae) in grasshoppers. Acrida 1972, 1, 111–120. [Google Scholar]
- Tounou, A.K. The potential of Paranosema (Nosema) locustae (Microsporidia: Nosematidae) and its combination with Metarhizium anisopliae var. acridum (Deuteromycotina: Hyphomycetes) for the control of locusts and grasshoppers in West Africa. Ph.D. Thesis, Leibniz University of Hanover, Hanover, Germany, September 2007; p. 126. [Google Scholar]
- Ericsson, J.D.; Kabaluk, J.T.; Goettel, M.S.; Myers, J.H. Spinosad interacts synergistically with the insect pathogen Metarhizium anisopliae against two exotic wireworms, Agriotes lineatus and Agriotes obscurus (Coleoptera: Elateridae). J. Econ. Entomol. 2007, 100, 31–38. [Google Scholar] [CrossRef]
- Inglis, G.D.; Johnson, D.L.; Goettel, M.S. Effects of temperature and thermoregulation on Mycosis by Beauveria bassiana in Grasshoppers. Biol. Control 1996, 7, 131–139. [Google Scholar] [CrossRef]
- Ewen, A.B.; Mukerji, M.K. Evaluation of Nosema locustae (Microsporida) as a control agent of grasshopper populations in Saskatchewan. J. Invertebr. Pathol. 1980, 35, 295–303. [Google Scholar] [CrossRef]
- Johnson, D.L.; Henry, J.E. Low rates of insecticides and Nosema locustae (Microsporidia: Nosematidae) on baits applied to roadsides for grasshopper (Orthoptera: Acrididae) control. J. Econ. Entomol. 1987, 80, 685–689. [Google Scholar] [CrossRef]
- Sieglaff, D.H.; Pereira, R.M.; Capinera, J.L. Pathogenicity of Beauveria bassiana and Metarhizium flavoviride (Deuteromycotina) to Schistocerca americana (Orthoptera: Acrididae). J. Econ. Entomol. 1997, 90, 1539–1545. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
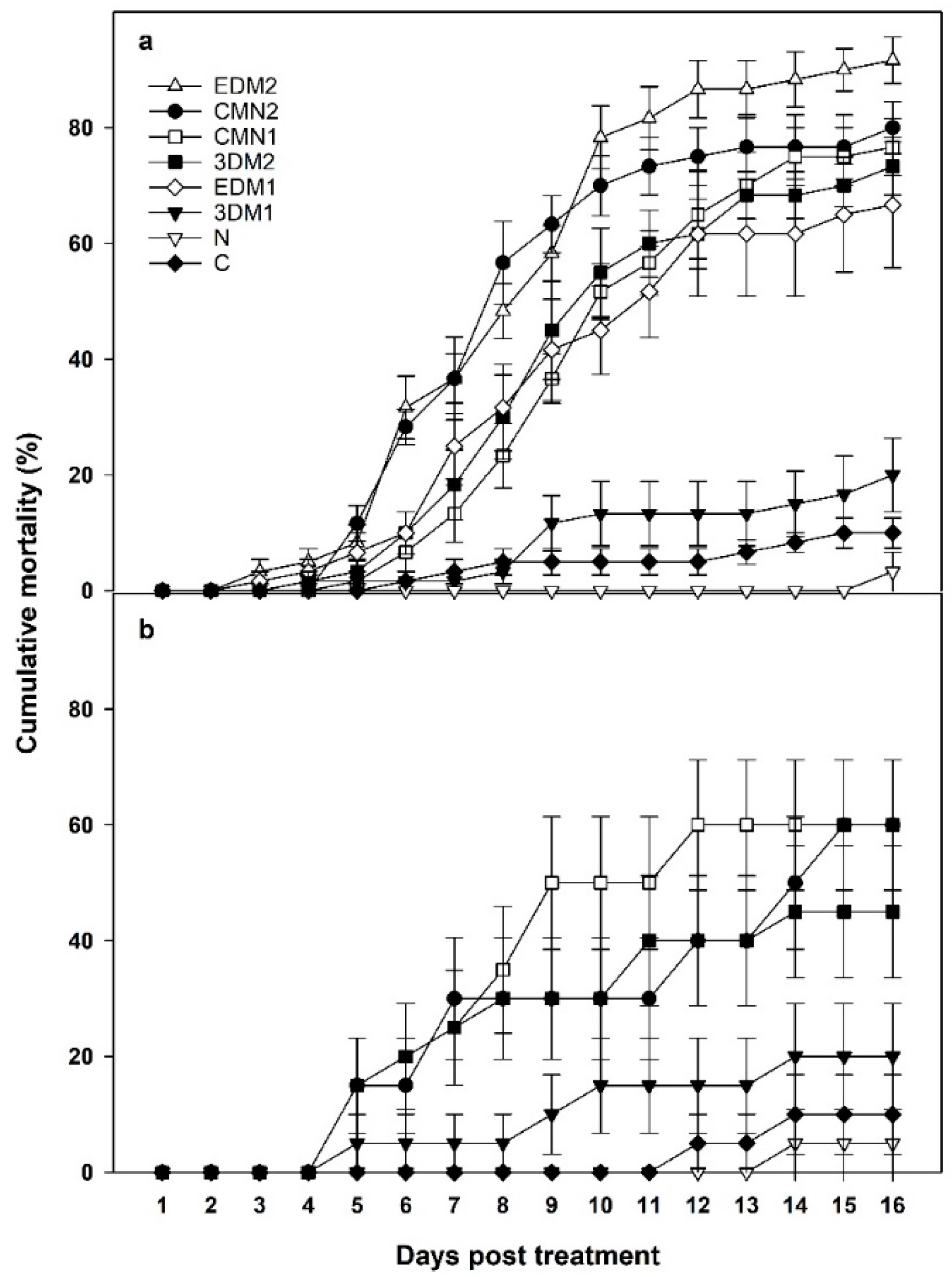{kind=link}
| Treatment † | Material | Rate |
|---|---|---|
| 3DM1 | Feeding low concentration of M. brunneum F52 for only 3 days | 0.15 × 109 conidia g−1 |
| 3DM2 | Feeding high concentration of M. brunneum F52 for only 3 days | 0.49 × 109 conidia g−1 |
| EDM1 | Feeding low concentration of M. brunneum F52 continuously for 14 days | 0.15 × 109 conidia g−1 |
| EDM2 | Feeding high concentration of M. brunneum F52 continuously for 14 days | 0.49 × 109 conidia g−1 |
| CMN1 | Feeding low concentration of both M. brunneum F52 and P. locustae for only 3 days | 0.15 × 109 spores g−1 |
| CMN2 | Feeding high concentration of both M. brunneum F52 and P. locustae for only 3 days | 0.49 × 109 spores g−1 |
| N | Exposure grasshoppers for P. locustae by itself | 2.2 × 106 spores g−1 |
| C | Untreated control | Untreated control |
| Treatment † | 7 Days | 10 Days | 14 Days | |||
|---|---|---|---|---|---|---|
| Laboratory | Greenhouse | Laboratory | Greenhouse | Laboratory | Greenhouse | |
| EDM2 | 37 a | 25 ab | 78 a | 30 ab | 88 a | 45 ab |
| CMN2 | 37 a | 30 a | 70 ab | 30 ab | 77 ab | 50 a |
| CMN1 | 13 bc | 25 ab | 52 c | 50 a | 75 ab | 60 a |
| 3DM2 | 18 b | -- | 55 bc | -- | 68 b | -- |
| EDM1 | 25 ab | 5 b,c | 45 c | 15 bc | 62 b | 20 bc |
| 3DM1 | 2 d | -- | 13 d | -- | 15 c | -- |
| C | 3 cd | 0 c | 5 de | 0 c | 8 c | 10 c |
| N | 0 d | 0 c | 0 e | 0 c | 0 d | 5 c |
| Treatment † | MST(days) § | Lower 95% | Upper 95% | |||
|---|---|---|---|---|---|---|
| Laboratory | Greenhouse | Laboratory | Greenhouse | Laboratory | Greenhouse | |
| EDM2 | 8.2 | -- | 7.2 | -- | 10.3 | -- |
| CMN2 | 8.2 | 13.5 | 7.1 | 8.4 | 9.2 | 18.4 |
| CMN1 | 10.5 | 10.1 | 9.1 | 7.3 | 11.9 | 12.8 |
| 3DM2 | 10.3 | -- | 9.1 | -- | 11.5 | -- |
| EDM1 | 10.7 | -- | 9.6 | -- | 11.9 | -- |
| Variables † | Coefficient | Standard Error | Z £ | p | R.R ¥ |
|---|---|---|---|---|---|
| Laboratory | |||||
| EDM2 vs. CMN2 | −0.08449 | 0.10067 | −0.84 | 0.4013 | 0.92 |
| EDM2 vs. untreated (C) | 1.04220 | 0.15864 | 6.57 | 0.0000 | 2.84 |
| CMN2 vs. untreated (C) | 0.54848 | 0.09494 | 5.78 | 0.0000 | 1.73 |
| Greenhouse | |||||
| CMN1 vs. CMN2 | −0.12217 | 0.40944 | −0.30 | 0.7654 | 0.88 |
| CMN1 vs. untreated (C) | 2.18975 | 0.76731 | 2.85 | 0.0043 | 8.93 |
| CMN2 vs. untreated (C) | 1.04986 | 0.38277 | 2.74 | 0.0061 | 2.86 |
| Treatment † | Pc% ζ | Pe% § | χ2 | Interaction |
|---|---|---|---|---|
| Laboratory | ||||
| CMN1 vs. 3DM1, and N | 75 | 22 | 97.838 | Synergism |
| CMN2 vs. 3DM2, and N | 77 | 71 | 0.965 | Additive |
| CMN1 vs. EDM1, and N | 75 | 65 | 2.691 | Additive |
| CMN2 vs. EDM2, and N | 77 | 89 | 9.909 | Antagonism |
| Greenhouse | ||||
| CMN1 vs. EDM1, and N | 60 | 32 | 7.463 | Synergism |
| CMN2 vs. EDM2, and N | 50 | 53 | 0.071 | Additive |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dakhel, W.H.; Latchininsky, A.V.; Jaronski, S.T. Efficacy of Two Entomopathogenic Fungi, Metarhizium brunneum, Strain F52 Alone and Combined with Paranosema locustae against the Migratory Grasshopper, Melanoplus sanguinipes, under Laboratory and Greenhouse Conditions. Insects 2019, 10, 94. https://doi.org/10.3390/insects10040094
Dakhel WH, Latchininsky AV, Jaronski ST. Efficacy of Two Entomopathogenic Fungi, Metarhizium brunneum, Strain F52 Alone and Combined with Paranosema locustae against the Migratory Grasshopper, Melanoplus sanguinipes, under Laboratory and Greenhouse Conditions. Insects. 2019; 10(4):94. https://doi.org/10.3390/insects10040094
Chicago/Turabian StyleDakhel, Wahid H., Alexandre V. Latchininsky, and Stefan T. Jaronski. 2019. "Efficacy of Two Entomopathogenic Fungi, Metarhizium brunneum, Strain F52 Alone and Combined with Paranosema locustae against the Migratory Grasshopper, Melanoplus sanguinipes, under Laboratory and Greenhouse Conditions" Insects 10, no. 4: 94. https://doi.org/10.3390/insects10040094
APA StyleDakhel, W. H., Latchininsky, A. V., & Jaronski, S. T. (2019). Efficacy of Two Entomopathogenic Fungi, Metarhizium brunneum, Strain F52 Alone and Combined with Paranosema locustae against the Migratory Grasshopper, Melanoplus sanguinipes, under Laboratory and Greenhouse Conditions. Insects, 10(4), 94. https://doi.org/10.3390/insects10040094