Bacillus thuringiensis Spores and Vegetative Bacteria: Infection Capacity and Role of the Virulence Regulon PlcR Following Intrahaemocoel Injection of Galleria mellonella


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultures
2.2. Mortality Tests in Galleria mellonella
2.3. Statistical Analysis
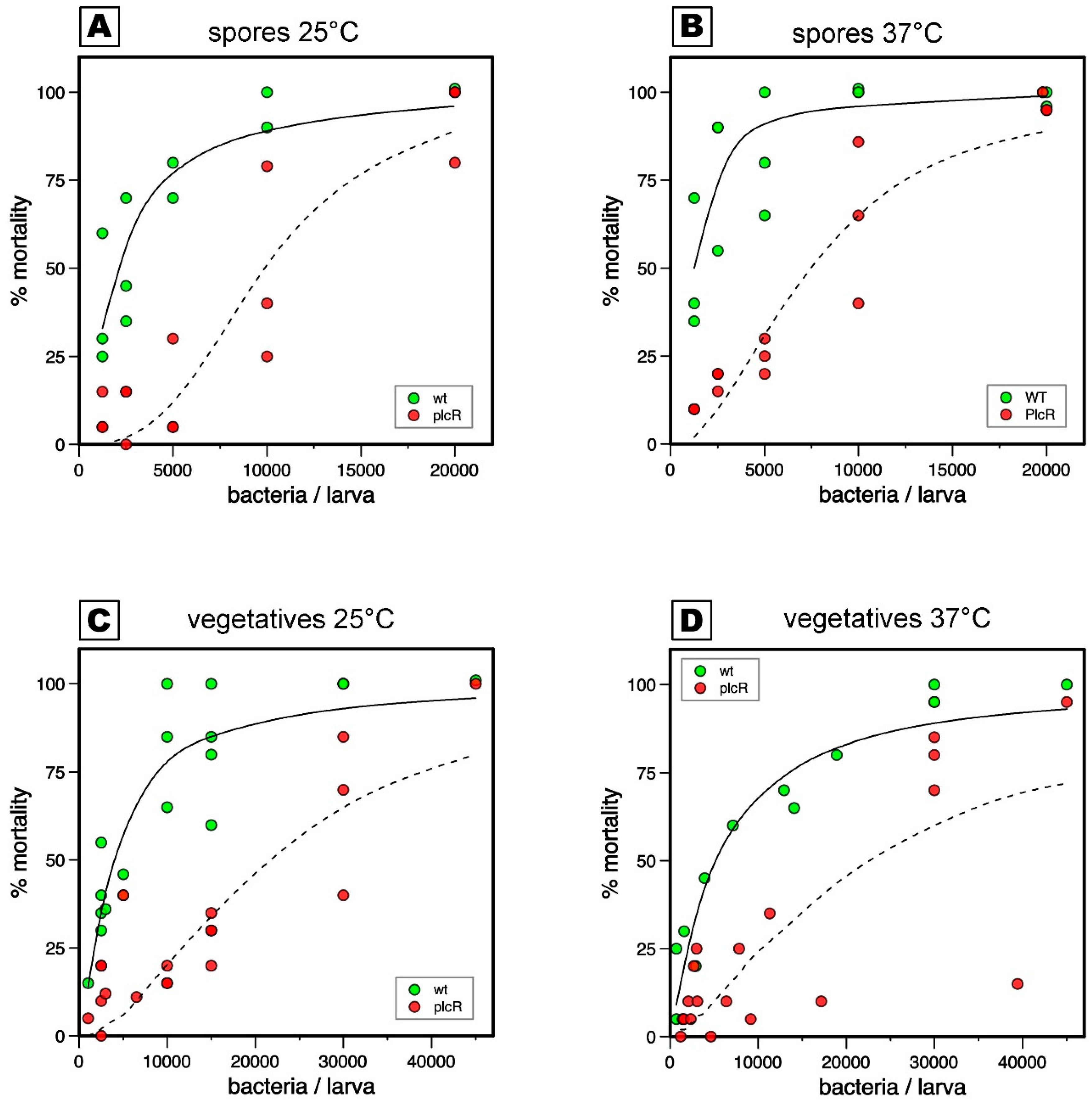
3. Results
3.1. Relative Roles of Temperature, Spores, and Vegetative Bacteria in Virulence Tests
3.2. The PlcR Regulon Is Required for Virulence When Spores or Vegetative Cells Are Mechanically Introduced into the Haemocoel
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: a century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [PubMed]
- Kolsto, A.B.; Tourasse, N.J.; Okstad, O.A. What sets Bacillus anthracis apart from other Bacillus species? Annu. Rev. Microbiol. 2009, 63, 451–476. [Google Scholar] [CrossRef]
- Patino-Navarrete, R.; Sanchis, V. Evolutionary processes and environmental factors underlying the genetic diversity and lifestyles of Bacillus cereus group bacteria. Res. Microbiol. 2017, 168, 309–318. [Google Scholar] [CrossRef]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberon, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachon, V.; Laprade, R.; Schwartz, J.L. Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: a critical review. J. Invertebr. Pathol. 2012, 111, 1–12. [Google Scholar] [CrossRef]
- Li, R.S.; Jarrett, P.; Burges, H.D. Importance of spores, crystals, and ∂-endotoxins in the pathogenicity of different varieties of Bacillus thuringiensis in Galleria mellonela and Pieris brassicae. J. Invertebr. Pathol. 1987, 50, 277–284. [Google Scholar] [CrossRef]
- Nielsen-LeRoux, C.; Gaudriault, S.; Ramarao, N.; Lereclus, D.; Givaudan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef]
- Dubois, T.; Faegri, K.; Perchat, S.; Lemy, C.; Buisson, C.; Nielsen-LeRoux, C.; Gohar, M.; Jacques, P.; Ramarao, N.; Kolsto, A.B.; et al. Necrotrophism is a quorum-sensing-regulated lifestyle in Bacillus thuringiensis. PLoS Pathog. 2012, 8, e1002629. [Google Scholar] [CrossRef] [PubMed]
- Slamti, L.; Lereclus, D. A cell-cell signaling peptide activates the PlcR virulence regulon in bacteria of the Bacillus cereus group. EMBO J. 2002, 21, 4550–4559. [Google Scholar] [CrossRef]
- Dubois, T.; Faegri, K.; Gelis-Jeanvoine, S.; Perchat, S.; Lemy, C.; Buisson, C.; Nielsen-LeRoux, C.; Gohar, M.; Jacques, P.; Ramarao, N.; et al. Correction: Necrotrophism Is a Quorum-Sensing-Regulated Lifestyle in Bacillus thuringiensis. PLoS Pathog. 2016, 12, e1006049. [Google Scholar] [CrossRef]
- Verplaetse, E.; Slamti, L.; Gohar, M.; Lereclus, D. Cell Differentiation in a Bacillus thuringiensis Population during Planktonic Growth, Biofilm Formation, and Host Infection. MBio. 2015, 6, e00138-15. [Google Scholar] [CrossRef]
- Salamitou, S.; Ramisse, F.; Brehelin, M.; Bourguet, D.; Gilois, N.; Gominet, M.; Hernandez, E.; Lereclus, D. The plcR regulon is involved in the opportunistic properties of Bacillus thuringiensis and Bacillus cereus in mice and insects. Microbiology 2000, 146, 2825–2832. [Google Scholar] [CrossRef]
- Gohar, M.; Faegri, K.; Perchat, S.; Ravnum, S.; Okstad, O.A.; Gominet, M.; Kolsto, A.B.; Lereclus, D. The PlcR virulence regulon of Bacillus cereus. PLoS ONE 2008, 3, e2793. [Google Scholar] [CrossRef]
- Slamti, L.; Perchat, S.; Huillet, E.; Lereclus, D. Quorum sensing in Bacillus thuringiensis is required for completion of a full infectious cycle in the insect. Toxins (Basel) 2014, 6, 2239–2255. [Google Scholar] [CrossRef] [PubMed]
- Bouillaut, L.; Ramarao, N.; Buisson, C.; Gilois, N.; Gohar, M.; Lereclus, D.; Nielsen-Leroux, C. FlhA influences Bacillus thuringiensis PlcR-regulated gene transcription, protein production, and virulence. Appl. Environ. Microbiol. 2005, 71, 8903–8910. [Google Scholar] [CrossRef]
- Candela, T.; Fagerlund, A.; Buisson, C.; Gilois, N.; Kolsto, A.B.; Okstad, O.A.; Aymerich, S.; Nielsen-Leroux, C.; Lereclus, D.; Gohar, M. CalY is a major virulence factor and a biofilm matrix protein. Mol. Microbiol. 2018. [Google Scholar] [CrossRef]
- Gominet, M.; Slamti, L.; Gilois, N.; Rose, M.; Lereclus, D. Oligopeptide permease is required for expression of the Bacillus thuringiensis plcR regulon and for virulence. Mol. Microbiol. 2001, 40, 963–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, B.; Elliot, S.L.; Ellis, R.J. Quantifying the reproduction of Bacillus thuringiensis HD1 in cadavers and live larvae of Plutella xylostella. J. Invertebr. Pathol. 2008, 98, 307–313. [Google Scholar] [CrossRef]
- Jensen, G.B.; Hansen, B.M.; Eilenberg, J.; Mahillon, J. The hidden lifestyles of Bacillus cereus and relatives. Environ. Microbiol. 2003, 5, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef]
- Raymond, B.; Johnston, P.R.; Nielsen-LeRoux, C.; Lereclus, D.; Crickmore, N. Bacillus thuringiensis: An impotent pathogen? Trends Microbiol. 2010, 18, 189–194. [Google Scholar] [CrossRef]
- Argolo-Filho, R.C.; Loguercio, L.L. Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches. Insects 2013, 5, 62–91. [Google Scholar] [CrossRef]
- Lecadet, M.M.; Blondel, M.O.; Ribier, J. Generalized transduction in Bacillus thuringiensis var. berliner 1715 using bacteriophage CP-54Ber. J. Gen. Microbiol. 1980, 121, 203–212. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Grizanova, E.V.; Whitten, M.M.; Mukherjee, K.; Greig, C.; Alikina, T.; Kabilov, M.; Vilcinskas, A.; Glupov, V.V.; Butt, T.M. Immuno-physiological adaptations confer wax moth Galleria mellonella resistance to Bacillus thuringiensis. Virulence 2016, 7, 860–870. [Google Scholar] [CrossRef]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Vogel, H.; Altincicek, B.; Glockner, G.; Vilcinskas, A. A comprehensive transcriptome and immune-gene repertoire of the lepidopteran model host Galleria mellonella. BMC Genom. 2011, 12, 308. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Krukova, N.A.; Glupov, V.V. Phagocytic activity and encapsulation rate of Galleria mellonella larval haemocytes during bacterial infection by Bacillus thuringiensis. J. Invertebr. Pathol. 2008, 98, 360–362. [Google Scholar] [CrossRef]
- Ramarao, N.; Lereclus, D. The InhA1 metalloprotease allows spores of the B. cereus group to escape macrophages. Cell. Microbiol. 2005, 7, 1357–1364. [Google Scholar] [CrossRef]
- Fedhila, S.; Nel, P.; Lereclus, D. The InhA2 metalloprotease of Bacillus thuringiensis strain 407 is required for pathogenicity in insects infected via the oral route. J. Bacteriol. 2002, 184, 3296–3304. [Google Scholar] [CrossRef]
- Peng, D.; Lin, J.; Huang, Q.; Zheng, W.; Liu, G.; Zheng, J.; Zhu, L.; Sun, M. A novel metalloproteinase virulence factor is involved in Bacillus thuringiensis pathogenesis in nematodes and insects. Environ. Microbiol. 2016, 18, 846–862. [Google Scholar] [CrossRef]
- Fang, S.; Wang, L.; Guo, W.; Zhang, X.; Peng, D.; Luo, C.; Yu, Z.; Sun, M. Bacillus thuringiensis bel protein enhances the toxicity of Cry1Ac protein to Helicoverpa armigera larvae by degrading insect intestinal mucin. Appl. Environ. Microbiol. 2009, 75, 5237–5243. [Google Scholar] [CrossRef] [PubMed]
- Kho, M.F.; Bellier, A.; Balasubramani, V.; Hu, Y.; Hsu, W.; Nielsen-LeRoux, C.; McGillivray, S.M.; Nizet, V.; Aroian, R.V. The pore-forming protein Cry5B elicits the pathogenicity of Bacillus sp. against Caenorhabditis elegans. PLoS ONE 2011, 6, e29122. [Google Scholar] [CrossRef]
- Fedhila, S.; Guillemet, E.; Nel, P.; Lereclus, D. Characterization of two Bacillus thuringiensis genes identified by in vivo screening of virulence factors. Appl. Environ. Microbiol. 2004, 70, 4784–4791. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ji, F.; Shang, H.; Zhu, Q.; Wang, P.; Xu, C.; Deng, Y.; Peng, D.; Ruan, L.; Sun, M. Gene clusters located on two large plasmids determine spore crystal association (SCA) in Bacillus thuringiensis subsp. finitimus strain YBT-020. PLoS ONE 2011, 6, e27164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Slamti, L.; Nielsen-LeRoux, C.; Lereclus, D.; Raymond, B. The social biology of quorum sensing in a naturalistic host pathogen system. Curr. Biol. 2014, 24, 2417–2422. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Temperature | Strain | LD50 * | [CI95%] | Ratio ∆plcR Mutant v.s. Bt407WT |
|---|---|---|---|---|
| 37 °C | Bt407WT | 1261 | [606–1757] | |
| ∆plcR mutant | 7344 | [5871–9050] | 5.8 | |
| 25 °C | Bt407WT | 2088 | [1391–2873] | |
| ∆plcR mutant | 9858 | [1015–7734] | 4.7 |
| Temperature | Strain | LD50 | [CI95%] | Ratio ∆plcR Mutant vs. Bt407WT |
|---|---|---|---|---|
| 37 °C | Bt407WT | 5148 | [3618–7087] | |
| ∆plcR mutant | 19,213 | [17,237–25,689] | 3.7 | |
| 25 °C | Bt407WT | 4032 | [2981–5260] | |
| ∆plcR mutant | 21,395 | [16,789–29,146] | 5.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buisson, C.; Gohar, M.; Huillet, E.; Nielsen-LeRoux, C. Bacillus thuringiensis Spores and Vegetative Bacteria: Infection Capacity and Role of the Virulence Regulon PlcR Following Intrahaemocoel Injection of Galleria mellonella. Insects 2019, 10, 129. https://doi.org/10.3390/insects10050129
Buisson C, Gohar M, Huillet E, Nielsen-LeRoux C. Bacillus thuringiensis Spores and Vegetative Bacteria: Infection Capacity and Role of the Virulence Regulon PlcR Following Intrahaemocoel Injection of Galleria mellonella. Insects. 2019; 10(5):129. https://doi.org/10.3390/insects10050129
Chicago/Turabian StyleBuisson, Christophe, Michel Gohar, Eugénie Huillet, and Christina Nielsen-LeRoux. 2019. "Bacillus thuringiensis Spores and Vegetative Bacteria: Infection Capacity and Role of the Virulence Regulon PlcR Following Intrahaemocoel Injection of Galleria mellonella" Insects 10, no. 5: 129. https://doi.org/10.3390/insects10050129
APA StyleBuisson, C., Gohar, M., Huillet, E., & Nielsen-LeRoux, C. (2019). Bacillus thuringiensis Spores and Vegetative Bacteria: Infection Capacity and Role of the Virulence Regulon PlcR Following Intrahaemocoel Injection of Galleria mellonella. Insects, 10(5), 129. https://doi.org/10.3390/insects10050129