The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards

 , ,
, ,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
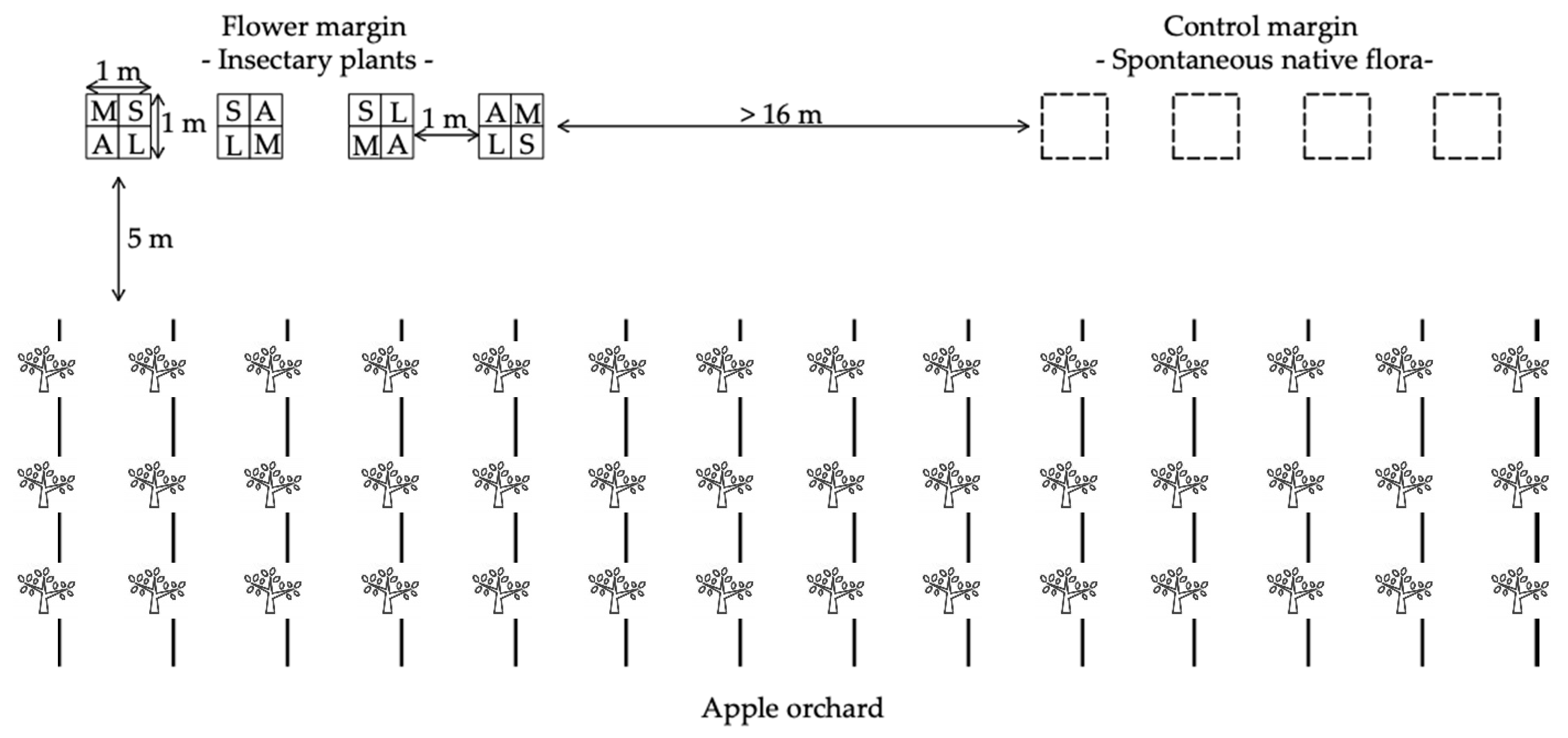
2.1. Study Area, Orchards and Treatments
2.2. Attractiveness of the Flower Margin to Natural Enemies and Phytophagous Insects
2.3. Assessment of D. plantaginea, E. lanigerum and Their Natural Enemies
2.4. Data Analysis
3. Results
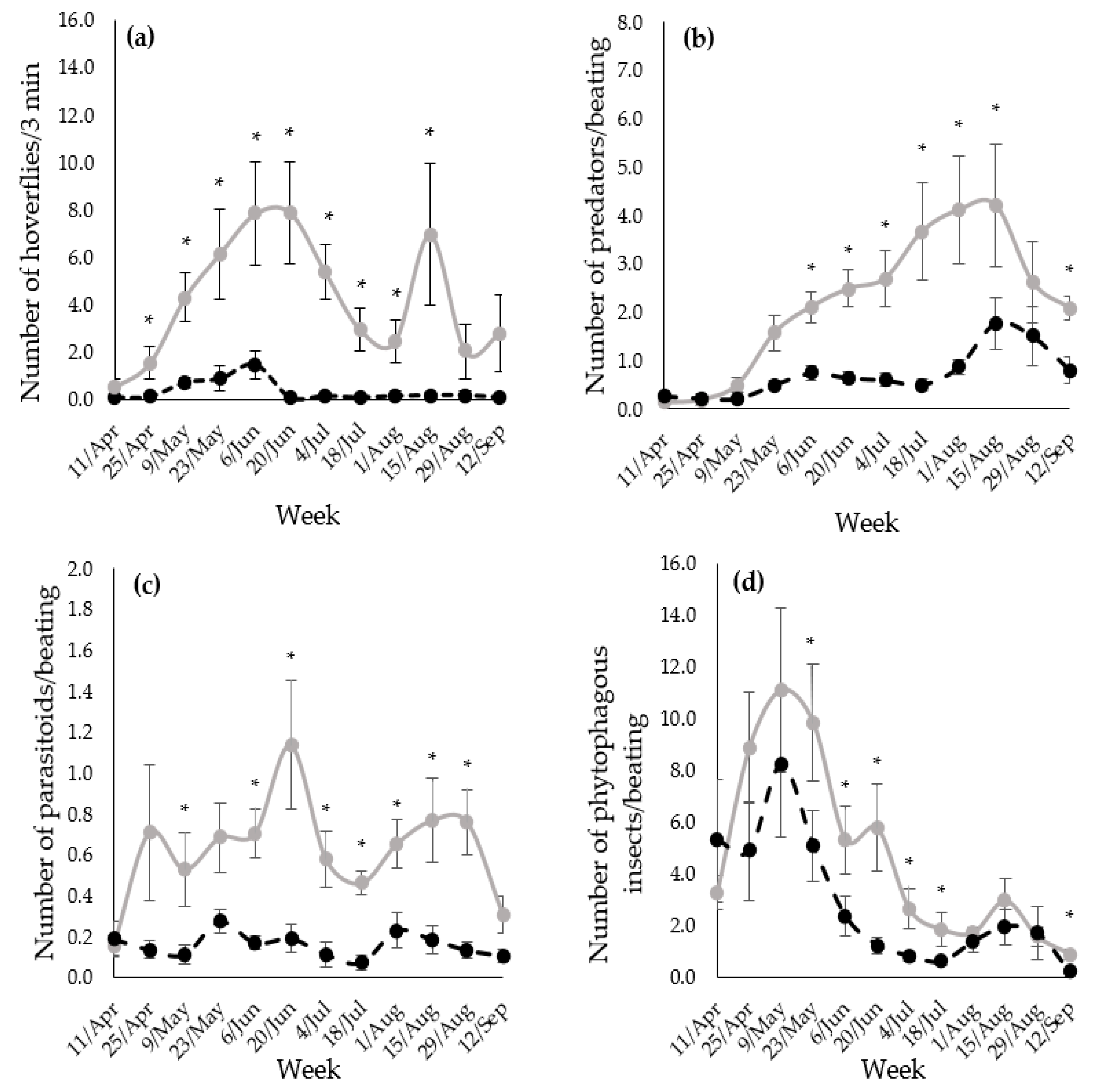
3.1. Attractiveness of the Margins to Natural Enemies and Phytophagous Insects
3.2. Assessment of D. plantaginea, E. lanigerum and Their Natural Enemies
3.3. Correlations between the Natural Enemies and D. plantaginea and E. lanigerum Colonies
3.3.1. Arthropods Found in the Margins
3.3.2. Arthropods Found in the D. plantaginea Colonies
3.3.3. Arthropods Found in the E. lanigerum Colonies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, O.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garibaldi, L.A.; Gemmill-Herren, B.; D’Annolfo, R.; Graeub, B.E.; Cunningham, S.A.; Breeze, T.D. Farming Approaches for Greater Biodiversity, Livelihoods, and Food Security. Trends Ecol. Evol. 2017, 32, 68–80. [Google Scholar] [CrossRef]
- Simon, S.; Bouvier, J.C.; Debras, J.F.; Sauphanor, B. Biodiversity and pest management in orchard systems. A review. Agron. Sustain. Dev. 2010, 30, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M.S. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A.; Nicholls, A.H. Biodiversity and Pest Management in Agroecosystems, 2nd ed.; Food Products Press, Haworth Press, Inc.: Binghamton, NY, USA, 2004. [Google Scholar]
- Boller, E.F.; Häni, F.; Poehling, H.M. Ecological Infrastructures: Ideabook on Functional Diversity at the Farm Level; IOBC WPRS: Winterthur, Switzerland, 2004. [Google Scholar]
- Rousselin, A.; Bevacqua, D.; Sauge, M.H.; Lescourret, F.; Mody, K.; Jordan, M.O. Harnessing the aphid life cycle to reduce insecticide reliance in apple and peach orchards. A review. Agron. Sustain. Dev. 2017, 37, 13. [Google Scholar] [CrossRef]
- Barbagallo, S.; Cocuzza, G.E.; Cravedi, P.; Komazaki, S. IPM Case Studies: Deciduous Fruit Tree. In Aphids as Crop Pests, 2nd ed.; CABI Publishing: Wallingford, UK, 2017; p. 700. [Google Scholar]
- Bonnemaison, L. Le puceron cendré du pommier (Dysaphis plantaginea Pass.). Morphologie et biologie. Méthodes de lutte. Annales des Épiphyties 1959, 3, 257–320. [Google Scholar]
- Brown, M.W.; Schmitt, J.J.; Ranger, S.; Hogmire, H.W. Yield reduction in apple by edaphic Wooly apple aphid (Homoptera, Aphididae) populations. J. Econ. Entomol. 1995, 88, 127–133. [Google Scholar] [CrossRef]
- Asante, S.K.; Danthanarayana, W.; Cairns, S.C. Spatial and temporal distribution patterns of Eriosoma lanigerum (Homoptera, Aphididae) on apple. Environ. Entomol. 1993, 22, 1060–1065. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.J.A.; Entling, M.H.; Moonen, A.C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Fitzgerald, J.D.; Solomon, M.G. Can flowering plants enhance numbers of beneficial arthropods in UK apple and pear orchards? Biocontrol Sci. Technol. 2004, 14, 291–300. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Beers, E.H.; Snyder, W.E. Flowers promote aphid suppression in apple orchards. Biol. Control 2013, 66, 8–15. [Google Scholar] [CrossRef]
- Albert, L.; Franck, P.; Gilles, Y.; Plantegenest, M. Impact of Agroecological Infrastructures on the Dynamics of Dysaphis plantaginea (Hemiptera: Aphididae) and Its Natural Enemies in Apple Orchards in Northwestern France. Environ. Entomol. 2017, 46, 528–537. [Google Scholar] [CrossRef]
- Miñarro, M.; Prida, E. Hedgerows surrounding organic apple orchards in north-west Spain: Potential to conserve beneficial insects. Agric. For. Entomol. 2013, 15, 382–390. [Google Scholar] [CrossRef]
- Odorizzi Santos, L.A.; Costa, M.B.; Lavigne, C.; Fernandes, O.A.; Bischoff, A.; Franck, P. Influence of the margin vegetation on the conservation of aphid biological control in apple orchards. J. Insect Conserv. 2018, 22, 465–474. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Plant Sociology: The Study of Plant Communities; MacGraw Hill: New York, NY, USA, 1932. [Google Scholar]
- Barahoei, H.; Madjdzadeh, S.M.; Mehrparvar, M. Aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) and their tritrophic relationships in Kerman province, Southeastern Iran. Iran. J. Anim. Biosyst. 2012, 8, 1–14. [Google Scholar]
- Rakhshani, E.; Kazemzadeh, S.; Stary, P.; Barahoei, H.; Kavallieratos, N.G.; Cetkovic, A.; Popovic, A.; Bodlah, I.; Tomanovic, Z. Parasitoids (Hymenoptera: Braconidae: Aphidiinae) of northeastern Iran: Aphidiine-aphid-plant associations, key and description of a new species. J. Insect Sci. 2012, 12. [Google Scholar] [CrossRef]
- Debras, J.F.; Torre, F.; Rieux, R.; Kreiter, S.; Garcin, M.S.; Van Helden, M.; Buisson, E.; Dutoit, T. Discrimination between agricultural management and the hedge effect in pear orchards (south-eastern France). Ann. Appl. Biol. 2006, 149, 347–355. [Google Scholar] [CrossRef]
- Horton, D.R.; Broers, D.A.; Lewis, R.R.; Granatstein, D.; Zack, R.S.; Unruh, T.R.; Moldenke, A.R.; Brown, J.J. Effects of mowing frequency on densities of natural enemies in three Pacific Northwest pear orchards. Entomol. Exp. Appl. 2003, 106, 135–145. [Google Scholar] [CrossRef]
- Wyss, E.; Niggli, U.; Nentwig, W. The impact of spiders on aphid populations in a strip-managed apple orchard. J. Appl. Entomol. 1995, 119, 473–478. [Google Scholar] [CrossRef]
- Wyss, E. The effects of weed strips on aphids and aphidophagous predators in an apple orchard. Entomol. Exp. Appl. 1995, 75, 43–49. [Google Scholar] [CrossRef]
- Dib, H.; Libourel, G.; Warlop, F. Entomological and functional role of floral strips in an organic apple orchard: Hymenopteran parasitoids as a case study. J. Insect Conserv. 2012, 16, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wackers, F. Getting more power from your flowers: Multi-functional flower strips enhance pollinators and pest control agents in apple orchards. Insects 2017, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, F.S. Foraging ecology of hoverflies—Morphology of the mouthparts in relation to feeding on nectar and pollen in some common urban species. Ecol. Entomol. 1981, 6, 245–262. [Google Scholar] [CrossRef]
- Schneider, F. Bionomics and Physiology of Aphidophagous Syrphidae. Annu. Rev. Entomol. 1969, 14, 103–124. [Google Scholar] [CrossRef]
- Wratten, S.D.; Bowie, M.H.; Hickman, J.M.; Evans, A.M.; Sedcole, J.R.; Tylianakis, J.M. Field boundaries as barriers to movement of hover flies (Diptera: Syrphidae) in cultivated land. Oecologia 2003, 134, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Miñarro, M.; Hemptinne, J.L.; Dapena, E. Colonization of apple orchards by predators of Dysaphis plantaginea: sequential arrival, response to prey abundance and consequences for biological control. BioControl 2005, 50, 403–414. [Google Scholar] [CrossRef]
- Dib, H.; Sauphanor, B.; Capowiez, Y. Effect of management strategies on arthropod communities in the colonies of rosy apple aphid, Dysaphis plantaginea Passerini (Hemiptera: Aphididae) in south-eastern France. Agric. Ecosyst. Environ. 2016, 216, 203–206. [Google Scholar] [CrossRef]
- Dib, H.; Simon, S.; Sauphanor, B.; Capowiez, Y. The role of natural enemies on the population dynamics of the rosy apple aphid, Dysaphis plantaginea Passerini (Hemiptera: Aphididae) in organic apple orchards in south-eastern France. Biol. Control 2010, 55, 97–109. [Google Scholar] [CrossRef]
- Stáry, P. Biology of Aphid Parasites (Hymenoptera: Aphidiidae) With Respect to Integrated Control, 6th ed.; Series Entomologica; Springer: Dordrecht, The Netherlands, 1970. [Google Scholar]
- Brown, M.W.; Mathews, C.R. Conservation biological control of rosy apple aphid, Dysaphis plantaginea (Passerini), in Eastern North America. Environ. Entomol. 2007, 36, 1131–1139. [Google Scholar] [CrossRef]
- Stewart-Jones, A.; Pope, T.W.; Fitzgerald, J.D.; Poppy, G.M. The effect of ant attendance on the success of rosy apple aphid populations, natural enemy abundance and apple damage in orchards. Agric. For. Entomol. 2008, 10, 37–43. [Google Scholar] [CrossRef]
- Cahenzli, F.; Pfiffner, L.; Daniel, C. Reduced crop damage by self-regulation of aphids in an ecologically enriched, insecticide-free apple orchard. Agron. Sustain. Dev. 2017, 37, 8. [Google Scholar] [CrossRef]
- Cross, J.V.; Solomon, M.G.; Babandreier, D.; Blommers, L.; Easterbrook, M.A.; Jay, C.N.; Jenser, G.; Jolly, R.L.; Kuhlmann, U.; Lilley, R.; et al. Biocontrol of pests of apples and pears in northern and central Europe: 2. Parasitoids. Biocontrol. Sci. Technol. 1999, 9, 277–314. [Google Scholar] [CrossRef]
- Peusens, G.; Buntinx, L.; Gobin, B. Parasitation of the parasitic wasp Ephedrus Persicae (Frogatt) on the rosy apple aphid Dysaphis Plantaginea (Passerini). Commun. Agric. Appl. Biol. Sci. 2006, 71, 369–374. [Google Scholar] [PubMed]
- Bribosia, E.; Bylemans, D.; Van Impe, G.; Migon, M. Assessing the suitability of alternative host aphids for Ephedrus persicae Froggatt (Hymenoptera: Braconidae), the main parasitoid attacking Dysaphis plantaginea (Passerini) (Homoptera: Aphididae) in Belgian apple orchards. In Proceedings of the Sixth International Symposium on Aphids, Rennes, France, 3–7 September 2001. [Google Scholar]
- Michelena, J.M.; Gonzalez, P.; Soler, E. Parasitoides afidiinos (Hymenoptera, Braconidae, Aphidiinae) de pulgones de cultivos agrícolas en la Comunidad Valenciana. Boletín de Sanidad Vegetal Plagas 2004, 30, 317–326. [Google Scholar]
- Aparicio, Y.; Gabarra, R.; Arno, J. Attraction of Aphidius ervi (Hymenoptera: Braconidae) and Aphidoletes aphidimyza (Diptera: Cecidomyiidae) to Sweet Alyssum and Assessment of Plant Resources Effects on their Fitness. J. Econ. Entomol. 2018, 111, 533–541. [Google Scholar] [CrossRef]
- Sadeghi, H.; Gilbert, F. Oviposition preferences of aphidophagous hoverflies. Ecol. Entomol. 2000, 25, 91–100. [Google Scholar] [CrossRef]
- James, D.G. Synthetic herbivore-induced plant volatiles as field attractants for beneficial insects. Environ. Entomol. 2003, 32, 977–982. [Google Scholar] [CrossRef]
- Almohamad, R.; Verheggen, F.J.; Haubruge, E. Searching and oviposition behavior of aphidophagous hoverflies (Diptera: Syrphidae): A review. Biotechnol. Agron. Soc. 2009, 13, 467–481. [Google Scholar]
- Almohamad, R.; Verheggen, F.J.; Francis, F.; Hance, T.; Haubruge, E. Discrimination of parasitized aphids by a hoverfly predator: Effects on larval performance, foraging, and oviposition behavior. Entomol. Exp. Appl. 2008, 128, 73–80. [Google Scholar] [CrossRef]
- Michaud, J.P. Sources of mortality in colonies of brown citrus aphid, Toxoptera citricida. BioControl 1999, 44, 347–367. [Google Scholar] [CrossRef]
- Mueller, T.F.; Blommers, L.H.M.; Mols, P.J.M. Earwig (Forficula auricularia) predation on the woolly apple aphid, Eriosoma lanigerum. Entomol. Exp. Appl. 1988, 47, 145–152. [Google Scholar] [CrossRef]
- Nicholas, A.H.; Spooner-Hart, R.N.; Vickers, R.A. Abundance and natural control of the woolly aphid Eriosoma lanigerum in an Australian apple orchard IPM program. BioControl 2005, 50, 271–291. [Google Scholar] [CrossRef]
- Lordan, J.; Alegre, S.; Moerkens, R.; Sarasua, M.J.; Alins, G. Phenology and interspecific association of Forficula auricularia and Forficula pubescens in apple orchards. Span. J. Agric. Res. 2015, 13. [Google Scholar] [CrossRef]
- Sauphanor, B.; Chabrol, L.; Darcier, F.F.; Sureau, F.; Lenfant, C. Side-effects of Diflubenzuron on a pear psilla predator—Forficula auricularia. Entomophaga 1993, 38, 163–174. [Google Scholar] [CrossRef]
- Bergh, J.C.; Short, B.D. Ecological and life-history notes on syrphid predators of woolly apple aphid in Virginia, with emphasis on Heringia calcarata. BioControl 2008, 53, 773–786. [Google Scholar] [CrossRef]
- Short, B.D.; Bergh, J.C. Feeding and egg distribution studies of Heringia calcarata (Diptera: Syrphidae), a specialized predator of woolly apple aphid (Homoptera: Eriosomatidae) in Virginia apple orchards. J. Econ. Entomol. 2004, 97, 813–819. [Google Scholar] [CrossRef]
- Rojo, S.; Gilbert, F.; Marcos-García, M.A.; Nieto, J.; Mier, M.P. A World Review of Predatory Hoverflies (Diptera, Syprhidae: Syrphinae) and Their Prey; CIBIO Ediciones: Alicante, Spain, 2003. [Google Scholar]
- Zhou, H.X.; Yu, Y.; Tan, X.M.; Chen, A.D.; Feng, J.G. Biological control of insect pests in apple orchards in China. Biol. Control 2014, 68, 47–56. [Google Scholar] [CrossRef]
- Monteiro, L.B.; Souza, A.; Belli, E.L. Parasitism on Eriosoma lanigerum (Homoptera: Aphididae) by Aphelinus mali (Hymenoptera: Encyrtidae) on apple orchards, in Fraiburgo county, state of Santa Catarina, Brazil. Rev. Bras. Frutic. 2004, 26, 550–551. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Beers, E.H.; Snyder, W.E. Complementary suppression of aphids by predators and parasitoids. Biol. Control 2015, 90, 83–91. [Google Scholar] [CrossRef]
- Bergh, J.C.; Stallings, J.W. Field evaluations of the contribution of predators and the parasitoid, Aphelinus mali, to biological control of woolly apple aphid, Eriosoma lanigerum, in Virginia, USA. BioControl 2016, 61, 155–165. [Google Scholar] [CrossRef]
- Lordan, J.; Alegre, S.; Gatius, F.; Sarasua, M.-J.; Alins, G. Woolly apple aphid Eriosoma lanigerum Hausmann ecology and its relationship with climatic variables and natural enemies in Mediterranean areas. Bull. Entomol. Res. 2015, 105, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Mackauer, M. Oviposition and host-feeding patterns in Aphelinus asychis (Hymenoptera, Aphelinidae) at different aphid densities. Ecol. Entomol. 1990, 15, 9–16. [Google Scholar] [CrossRef]
- Wu, Z.S.; Heimpel, G.E. Dynamic egg maturation strategies in an aphid parasitoid. Physiol. Entomol. 2007, 32, 143–149. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Sampling | Arthropod Group | Treatment | Year | Year*Treatment | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control Margin ± SE | Flower Margin ± SE | F1,12 | p | F1,12 | p | F1,12 | p | ||
| VO | Adult hoverflies | 0.384 ± 0.077 | 4.242 ± 0.463 | 109.052 | <0.001 | 0.968 | 0.348 | 3.182 | 0.114 |
| BTS | All predators | 0.724 ± 0.122 | 2.203 ± 0.332 | 39.991 | <0.001 | 1.323 | 0.278 | 0.773 | 0.405 |
| Thysanoptera | 0.035 ± 0.012 | 0.224 ± 0.039 | 42.784 | <0.001 | 6.663 | 0.026 | 2.811 | 0.128 | |
| Araneae | 0.321 ± 0.049 | 0.679 ± 0.140 | 6.340 | 0.034 | 0.175 | 0.684 | 0.314 | 0.589 | |
| Heteroptera | 0.280 ± 0.114 | 1.085 ± 0.307 | 17.482 | <0.001 | 4.676 | 0.057 | 0.230 | 0.644 | |
| Coleoptera | 0.045 ± 0.009 | 0.139 ± 0.048 | 6.100 | 0.038 | 7.036 | 0.023 | 5.506 | 0.046 | |
| Other predators | 0.044 ± 0.015 | 0.074 ± 0.007 | 3.810 | 0.081 | 2.003 | 0.185 | 3.446 | 0.094 | |
| Parasitoid wasps | 0.161 ± 0.015 | 0.623 ± 0.083 | 30.017 | <0.001 | 0.282 | 0.606 | 0.799 | 0.394 | |
| Phytophagous insects | 2.828 ± 0.604 | 4.66 ± 0.598 | 7.655 | 0.025 | 1.654 | 0.229 | 0.002 | 0.968 | |
| Type of Sampling | Arthropod Group | Syrphidae in the FM | Syrphidae in the CM | Predators in the FM | Predators in the CM | Parasitoids in the FM | Parasitoids in the CM | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ρ | p | ρ | p | ρ | p | ρ | p | ρ | p | ρ | p | |||
| FM | VO | Syrphidae | − | − | 0.192 | 0.06 | 0.344 | <0.001 | −0.025 | 0.806 | 0.38 | <0.001 | 0.01 | 0.92 |
| BTS | Predators | 0.344 | <0.001 | 0.038 | 0.716 | − | − | 0.446 | <0.001 | 0.514 | <0.001 | 0.08 | 0.937 | |
| Parasitoids | 0.38 | <0.001 | 0.185 | 0.072 | 0.514 | <0.001 | 0.214 | 0.036 | − | − | 0.32 | 0.002 | ||
| Phytophagous insects | 0.434 | <0.001 | 0.183 | 0.075 | −0.124 | 0.229 | −0.383 | <0.001 | 0.273 | 0.007 | 0.122 | 0.236 | ||
| CM | VO | Syrphidae | 0.192 | 0.06 | − | − | 0.038 | 0.716 | 0.027 | 0.793 | 0.184 | 0.072 | 0.126 | 0.221 |
| BTS | Predators | −0.025 | 0.806 | 0.027 | 0.793 | 0.446 | <0.001 | − | − | 0.214 | 0.036 | 0.176 | 0.09 | |
| Parasitoids | 0.01 | 0.92 | 0.126 | 0.221 | 0.008 | 0.937 | 0.176 | 0.086 | 0.32 | 0.002 | − | − | ||
| Phytophagous insects | 0.302 | 0.003 | 0.158 | 0.125 | −0.125 | 0.227 | −0.193 | 0.06 | 0.129 | 0.212 | 0.234 | 0.022 | ||
| Type of Sampling | Arthropod Group | Syrphidae Predators in the RAA Colonies | Non-Syrphidae Predators in the RAA Colonies | Parasiotids in the RAA Colonies | ||||
|---|---|---|---|---|---|---|---|---|
| ρ | p | ρ | p | ρ | p | |||
| FM | VO | Syrphidae | 0.312 | 0.138 | 0.333 | 0.104 | 0.621 | 0.001 |
| BTS | Predators | 0.309 | 0.142 | 0.146 | 0.498 | 0.488 | 0.016 | |
| Parasitoids | 0.173 | 0.418 | 0.272 | 0.188 | 0.281 | 0.183 | ||
| Phytophagous insects | 0.312 | 0.138 | 0.025 | 0.908 | 0.226 | 0.289 | ||
| CM | VO | Syrphidae | 0.253 | 0.234 | −0.039 | 0.854 | 0.105 | 0.626 |
| BTS | Predators | 0.175 | 0.414 | 0.155 | 0.471 | 0.428 | 0.037 | |
| Parasitoids | 0.449 | 0.028 | 0.244 | 0.250 | 0.483 | 0.017 | ||
| Phytophagous insects | 0.116 | 0.589 | 0.414 | 0.040 | 0.031 | 0.887 | ||
| RAA | RAA colonies | % shoots infested with RAA colonies | 0.536 | 0.007 | 0.233 | 0.273 | 0.606 | 0.002 |
| Syrphidae predators | − | − | −0.057 | 0.793 | 0.658 | <0.001 | ||
| Non-Syrphidae predators | −0.057 | 0.793 | − | − | 0.191 | 0.373 | ||
| Parasitoids | 0.658 | <0.001 | 0.191 | 0.373 | − | − | ||
| Type of Sampling | Arthropod Group | Predators in the WAA Colonies | Parasitoids in the WAA Colonies | |||
|---|---|---|---|---|---|---|
| ρ | p | ρ | p | |||
| FM | VO | Syrphidae | 0.298 | 0.078 | 0.181 | 0.292 |
| BST | Predators | 0.139 | 0.419 | 0.055 | 0.752 | |
| Parasitoids | 0.295 | 0.081 | 0.223 | 0.190 | ||
| Phytophagous insects | 0.022 | 0.898 | 0.079 | 0.647 | ||
| CM | VO | Syrphidae | 0.107 | 0.536 | −0.141 | 0.413 |
| BTS | Predators | 0.031 | 0.857 | −0.156 | 0.363 | |
| Parasitoids | 0.093 | 0.588 | −0.101 | 0.557 | ||
| Phytophagous insects | 0.154 | 0.369 | −0.026 | 0.881 | ||
| WAA | WAA colonies | % shoots infested with WAA colonies | 0.227 | 0.183 | 0.597 | <0.001 |
| Predators | − | − | 0.253 | 0.136 | ||
| Parasitoids | 0.253 | 0.136 | − | − | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Gasol, N.; Avilla, J.; Aparicio, Y.; Arnó, J.; Gabarra, R.; Riudavets, J.; Alegre, S.; Lordan, J.; Alins, G. The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards. Insects 2019, 10, 148. https://doi.org/10.3390/insects10050148
Rodríguez-Gasol N, Avilla J, Aparicio Y, Arnó J, Gabarra R, Riudavets J, Alegre S, Lordan J, Alins G. The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards. Insects. 2019; 10(5):148. https://doi.org/10.3390/insects10050148
Chicago/Turabian StyleRodríguez-Gasol, Neus, Jesús Avilla, Yahana Aparicio, Judit Arnó, Rosa Gabarra, Jordi Riudavets, Simó Alegre, Jaume Lordan, and Georgina Alins. 2019. "The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards" Insects 10, no. 5: 148. https://doi.org/10.3390/insects10050148
APA StyleRodríguez-Gasol, N., Avilla, J., Aparicio, Y., Arnó, J., Gabarra, R., Riudavets, J., Alegre, S., Lordan, J., & Alins, G. (2019). The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards. Insects, 10(5), 148. https://doi.org/10.3390/insects10050148