Identification and Virulence Characterization of Two Akanthomyces attenuatus Isolates Against Megalurothrips usitatus (Thysanoptera: Thripidae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Soil and Isolation of Fungi
2.2. Insect Rearing
2.3. Morphological Characterization
2.4. Radial Growth and Conidial Yield
2.5. DNA Extraction, PCR Amplification, and Sequence Analysis
2.6. Virulence of the Fungi in the Laboratory
2.7. Transmission Electron Microscopy
2.8. Statistical Analysis
3. Results
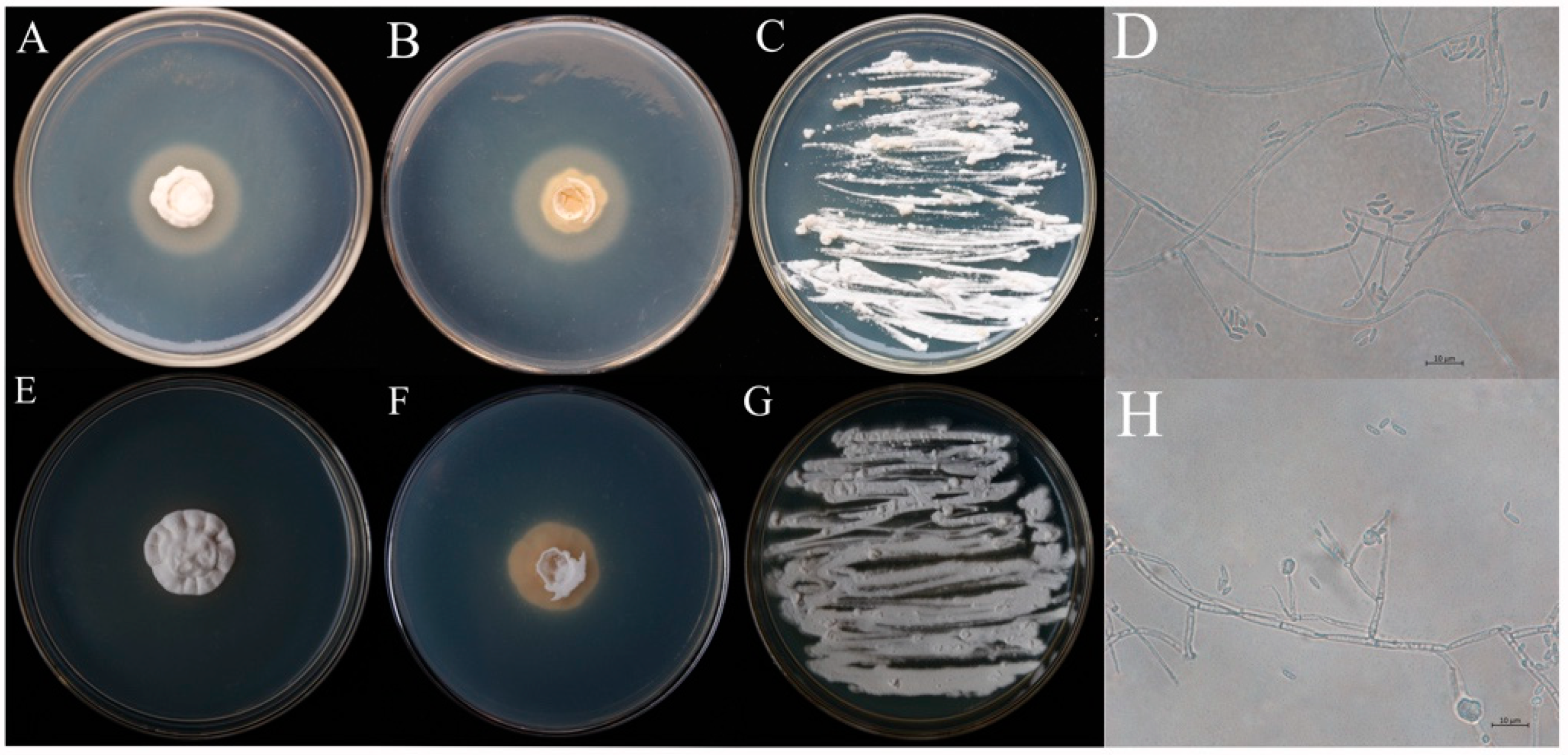
3.1. Morphological Identification of Fungi
3.2. Molecular Analyses
3.2.1. BLASTN Comparisons
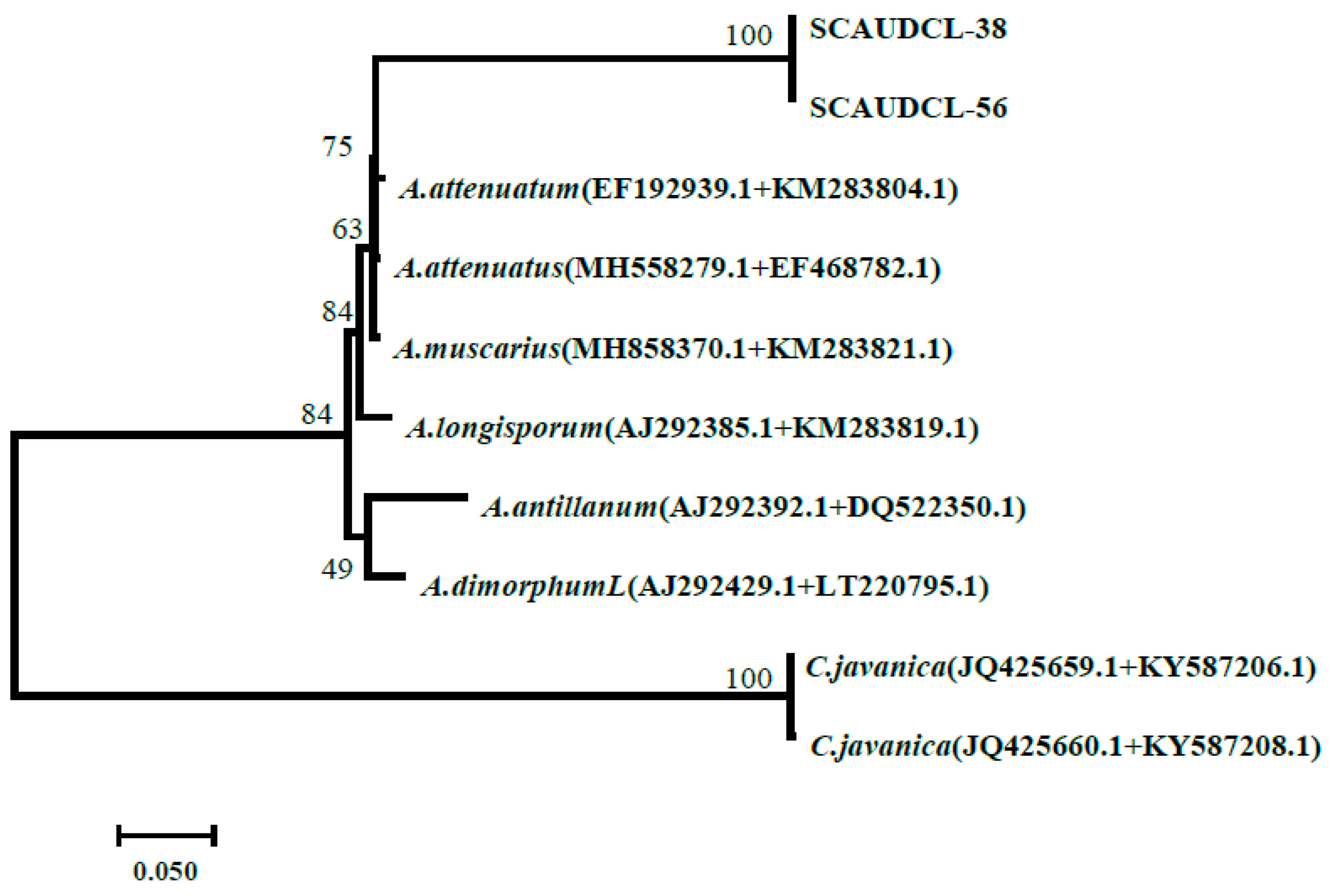
3.2.2. Phylogenetic Analysis
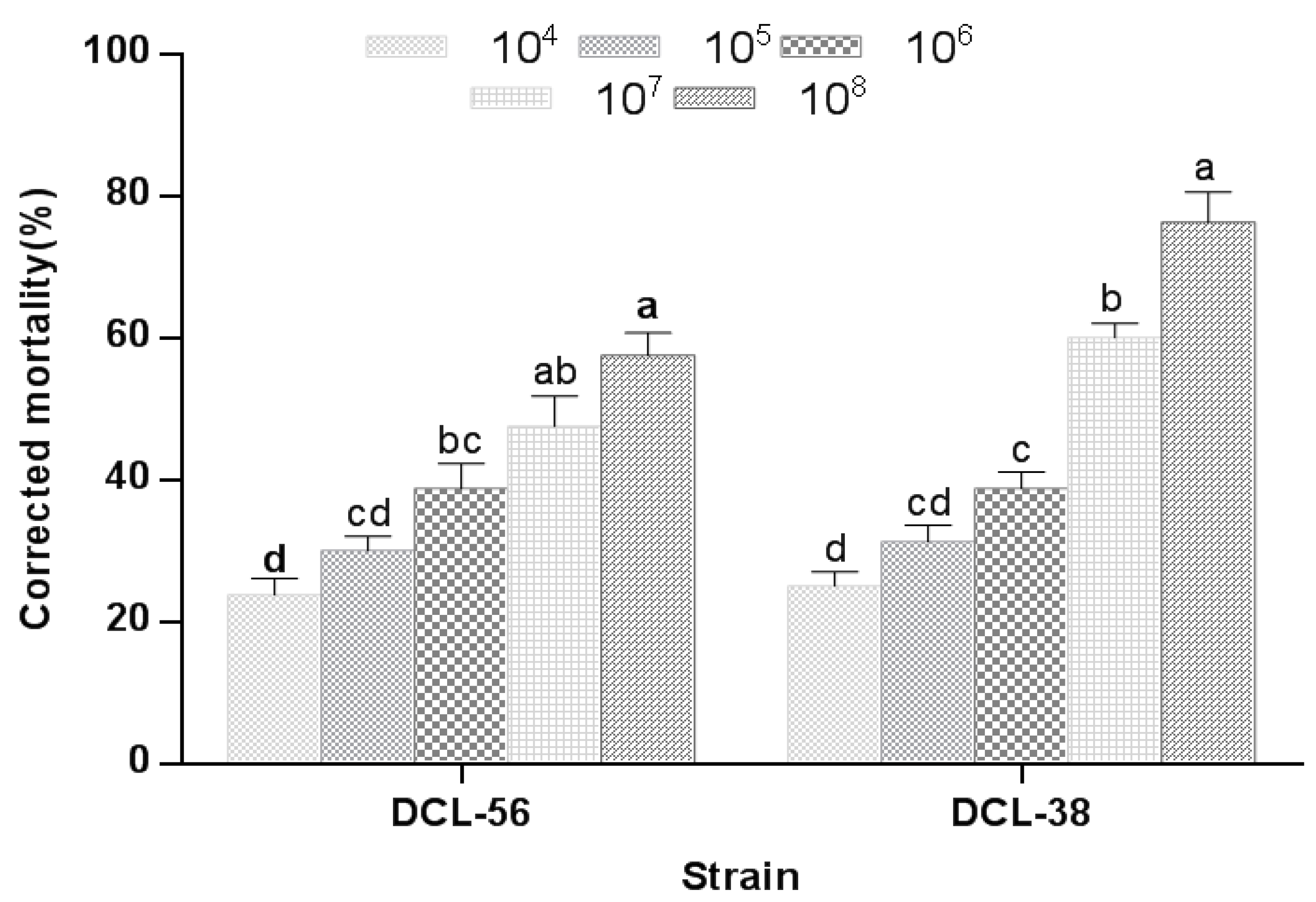
3.3. Virulence of A. attenuatus against M. usitatus
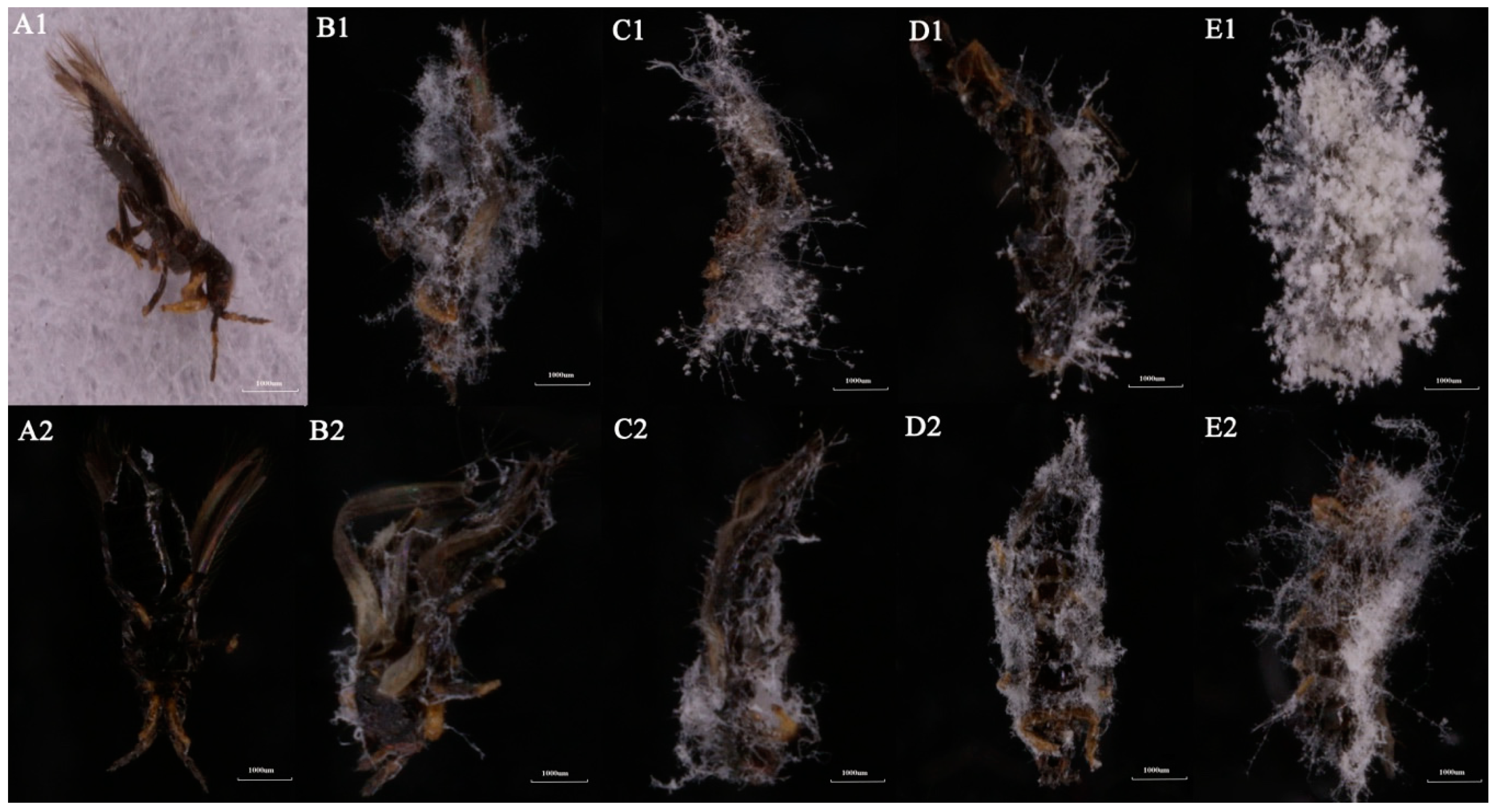
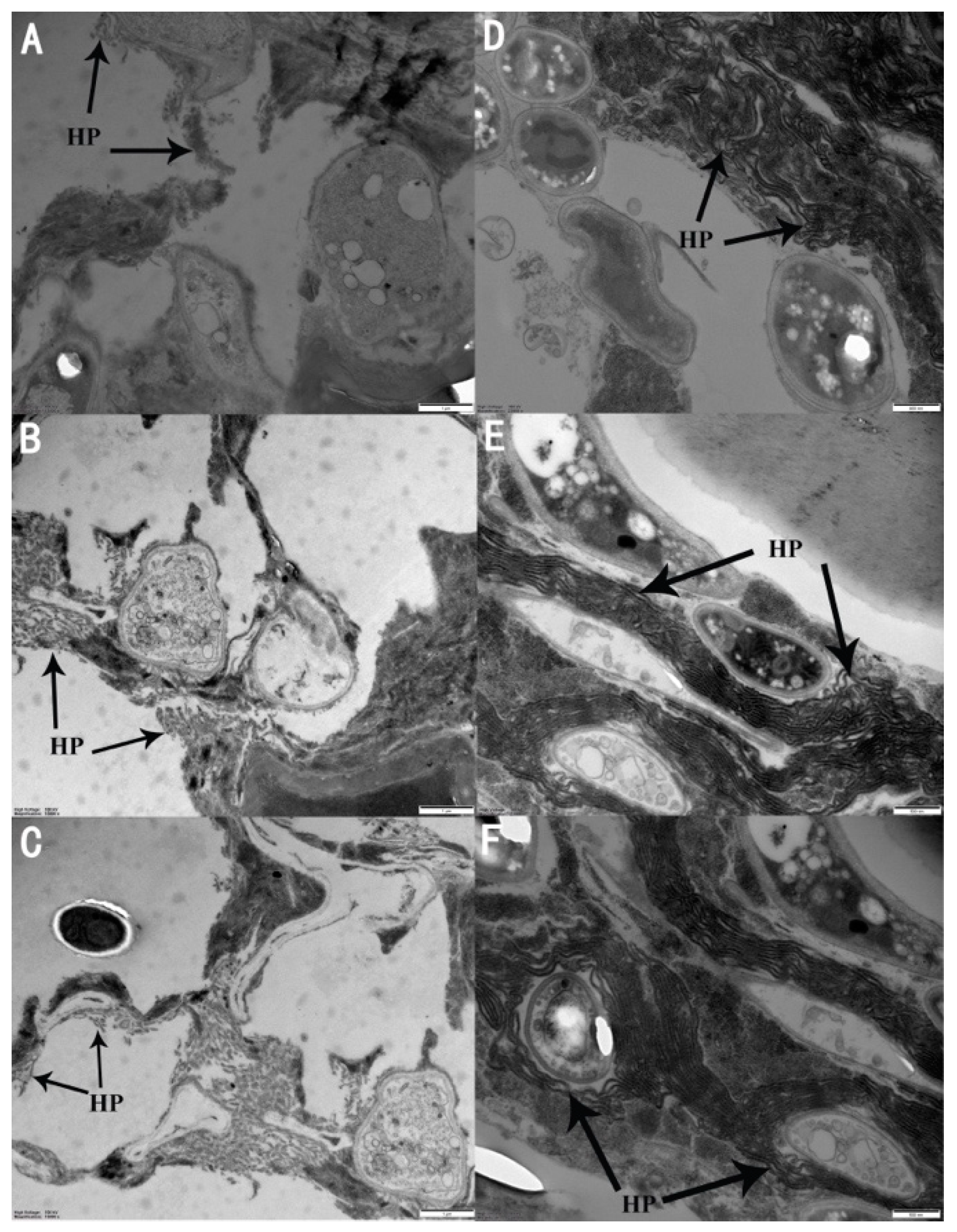
3.4. Microscopic Examination of A. attenuatus Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tang, L.D.; Yan, K.L.; Fu, B.L. The life table parameters of Megalurothrips usitatus (Thysanopera: Thripidea) on four leguminous crops. Fla. Entomol. 2015, 98, 620–625. [Google Scholar] [CrossRef]
- Shipp, J.L.; Wang, K.; Binns, M.R. Economic injury levels for western flower thrips (Thysanoptera: Thrippidae) on greenhouse cucumber. J. Econ. Entomol. 2000, 93, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.P.; Jia, W.T.; Zheng, X.; Zhang, L.; Sangbaramou, R.; Tan, S.Q.; Liu, Y.Q.; Shi, W.P. Predation functional response and life table parameters of Orius sauter (Hemiptera: Antocoridae) feeding on Megalurothrips usitatus (Thysanoptera: Thripidae). Fla. Entomol. 2018, 101, 254–259. [Google Scholar] [CrossRef]
- Chen, W.S. A study on the relationship between thrips and the yield of peanut [Taeniothrips distalis Karny, Thrips spp. and Scirtothrips dorsalis]. Res. Bull. Taiwan Dais. 1980, 14, 51–57. [Google Scholar]
- Gao, Y.L.; Lei, Z.R.; Reitz, S.R. Western flower thrips resistance to insecticides: Detection, mechanisms and management strategies. Pest Manag. Sci. 2012, 68, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Brownbridge, M.; Nelson, T.L.; Hackell, D.L. Field application of biopolymer-coated Beauveria bassiana F418 clover root weevil (Sitona Lepidus) control in Waikato and Manawatu. N. Z. Plant Prot. 2006, 59, 304–311. [Google Scholar]
- Jensen, S.E. Mechanisms associated with methiocarb resistance in Franklinie occidentalis (Thysanoptera: Thripidae). J. Econ. Entomol. 2000, 93, 464–471. [Google Scholar] [CrossRef]
- Fan, Y.M.; Tong, X.L.; Gao, L.I.; Wang, M.; Liu, Z.Q.; Zhang, Y.; Yang, Y. The spatial aggregation pattern of dominant species of thrips on cowpeain Hainan. J. Environ. Entomol. 2013, 35, 737–743. [Google Scholar]
- Wright, S.P.; Filotas, M.J.; Sanderson, J.P. Comparative efficacy of emulsifiable-oil, wettable-powder, and unformulated-powder preparations of Beauveria bassiana against the melon aphid Aphis gossypii. Biocontrol Sci. Technol. 2016, 26, 894–914. [Google Scholar] [CrossRef]
- Ansari, M.A.; Brownbridge, M.; Shah, F.A.; Butt, T.M. Efficacy of entomopathogenic fungi against soil-dwelling life stages of western flower thrips, Frankliniella occidentalis, in plant-growing media. Entomol. Exp. Appl. 2008, 127, 80–87. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, M.A.; Whittaker, M.; Prasad, M.; Butt, T.M. Control of western flower thrips (Frankliniella occidentalis) pupae with Metarhizium anisopliae in peat and pear alternative growing media. Biol. Control 2007, 40, 293–297. [Google Scholar] [CrossRef]
- Saito, T.; Brownbrideg, M. Compatibility of soil-dwelling predators and microbial agents and their efficacy in controlling soil-dwelling stages of western flower thrips Frankliniella occdentalis. Biol. Control 2016, 92, 92–100. [Google Scholar] [CrossRef]
- Brownbridge, M.; Butt, T.M. Fungal Pathogens of Thrips, 1st ed.; CABI: Wallingford, UK, 1997; pp. 399–433. [Google Scholar]
- Skinner, M.; Gouli, S.; Frank, C.E.; Parker, B.L.; Kim, J.S. Management of Frankliniella occidentalis (Thysanoptera: Thripidae) with granular formulations of entomopathogenic fungi. Biol. Control 2012, 63, 246–252. [Google Scholar] [CrossRef]
- Kivett, J.M.; Cloyd, R.A.; Bello, N.M. Evaluation of entomopathogenic fungi against the western flower thrips (Thysanoptera: Thripidae) under laboratory conditions. J. Entomol. Sci. 2016, 51, 274–291. [Google Scholar] [CrossRef]
- Pong, K.K.; Ramle, M.; Wahizatul, A.A.; Norman, K.; Siti, R.A. Genetic variation of entomopathogenic fungi, Metarhizium anisopliae and Isaria amoenerosea and their pathogenicity against subterranean termite, Coptotermes curvignathus. J. Oil Palm Res. 2017, 29, 35–46. [Google Scholar]
- Ainsworth, G.C.; Bisby, G.R.; Kirk, P.M. Ainsworth & Bisby’s Dictionary of the Fungi, 5th ed.; CABI: Wallingford, UK, 2008; p. 771. [Google Scholar]
- Wang, T.J.; Huang, J.; You, M.; Guan, X.; Liu, B. Toxicity and feeding deterrence of crude toxin extracts of Lecanicillium (Verticillium) lecanii (Hyphomycetes) against sweet potato whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). Pest Manag. Sci. 2007, 63, 381–387. [Google Scholar] [CrossRef]
- Gottel, M.S.; Koike, M.; Kim, J.J.; Aiuchi, D.; Shinya, R.; Brodeur, J. Potential of Leacanicillium spp. For management of insects nematodes and plant diseases. J. Invert. Pathol. 2008, 98, 256–261. [Google Scholar] [CrossRef]
- Lu, L.; Cheng, B.; Du, D.; Hu, X.; Peng, A.; Pu, Z.; Zhang, X.; Huang, Z.; Chen, G. Morphological, molecular and virulence characterization of three Lecanicillium species infecting Asian citrus psyllids in Huangyuan citrus groves. J. Invert. Pathol. 2015, 125, 45–55. [Google Scholar] [CrossRef]
- Inglis, G.D.; Johnson, D.L.; Kawchuk, L.M.; Goettel, M.S. Effect of soil texture and soil sterilization on susceptibility of ovipositing grasshoppers to Beauveria bassiana. J. Invert. Pathol. 1998, 71, 73–78. [Google Scholar] [CrossRef]
- Imoulan, A.; Alaoui, A.; Meziane, A. Natural occurrence of soil-borne entomopathogenic fungi in the Moroccan Endemic forest of Argania spinose and their pathogenicity to Ceratitis capitata. World J. Microbiol. Biotechnol. 2011, 27, 2619–2628. [Google Scholar] [CrossRef]
- Imoulan, A.; Wu, H.J.; Lu, W.L.; Li, Y.; Li, B.B.; Yand, R.H.; Wand, W.J.; Wand, X.J.; Kirk, P.M.; Yao, Y.J. Beauveria meogensis sp. nov., a new fungus of the entomopathogenic genus from China. J. Invert. Pathol. 2016, 139, 74–81. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Media composition influences on growth, enzyme activity and virulence of the entomopathogen hyphomycete Isaria fumosorosea. Entomol. Exp. Appl. 2009, 131, 30–38. [Google Scholar] [CrossRef]
- Agrawal, Y.; Mual, P.; Shenoy, B.D. Multi-gene genealogies reveal cryptic species Beauveria rudraprayagi sp. nov. from India. Mycosphere 2014, 5, 719–736. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: Genes for Phylogenetics; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF 1—alpha sequences: Evidence for cryrtic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar]
- Bischoff, J.F.; Rehne, S.A.; Humber, R.A. A multilocus phylogeny of the M. anisopliae. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladisitics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Ali, S.; Zhang, C.; Wang, Z.Q.; Wang, X.M.; Wu, J.H.; Cuthbertson, A.G.S.; Shao, Z.F.; Qiu, B.L. Toxicological and biochemical basis of synergism between the entomopathogenic fungus Lecanicillium muscarium and the insecticide matrine against Bemisia tabaci (Gennadius). Sci. Reports 2017, 7, 46558. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lei, Z.R.; Zhang, Q.W. Observation of infection process of Metarhizium anisopliae on Plutella xylostella larvae with transmission electron microscopy. Acta Ent. Sin. 2006, 49, 1042–1045. [Google Scholar]
- Finney, J.G.; Smith, D.F.; Skeeters, D.E.; Auvenshine, C.D. MMPI alcoholism scales: Factor structure and content analysis. J. Stud. Alcohol 1971, 32, 1055–1060. [Google Scholar]
- SAS, T.C.J.; Gerver, W.J.M.; Bruin, R.D.; Mulder, P.G.H.; Cole, T.J.; Wall, W.D.; Hokken-Koelega, A.C.S. Body proportions during 6 years of GH treatment in children with short born small for gestational age participating in a randomised, double-blind, dose-response trial. Clin. Endocrinol. 2000, 53, 675–681. [Google Scholar] [CrossRef]
- Rehner, S.A.; Minnis, D.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Wang, D.K.; Deng, J.X.; Pei, Y.F.; Li, T.; Jin, Z.Y.; Liang, L.; Wang, W.K.; Li, L.D.; Dong, X.L. Identification and virulence characterization of entomopathogenic fungus Lecanicillium attenuatus against the pea aphid Acyrthos iphonpisum (Hemiptera: Aphididae). Appl. Entomol. Zool. 2017, 52, 511–518. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. Sequence Matrix: Concatenation software for the fast assembly of multi-gene data sets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. Metarhizium frigidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. flavoviride complex. Mycologia 2006, 98, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Montalva, C.; Valenzuela, E.; Barta, M.; Rojas, E.; Arismendi, N.; Juscelino, R.; Humber, R.A. Lecanicillium attenuatus isolates affecting the invasive cypress aphid (Cinaracupressi) in Chile. Biol. Control 2017, 62, 625–637. [Google Scholar]
- Kim, J.J.; Goettel, M.S.; Gillespie, D.R. Potential of Lecanicillium species for dual microbial control of aphids and the cucumber powdery mildew fungus, Sphaerotheca fuliginea. Biol. Control 2007, 40, 327–332. [Google Scholar] [CrossRef]
- Jandricic, S.E.; Filotas, M.; Sanderson, J.P.; Wraight, S.P. Pathogenicity of conidia-based preparations of entomopathogenic fungi against the greenhouse pest aphids Myzus persicae, Aphis gossypii, and Aulacorthum solani (Hemiptera: Aphididae). J. Invertebr. Pathol. 2014, 118, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Vu, V.; Hong, S.; Kim, K. Selection of entomopathogenic fungi for aphid control. Bio. Sci. Bioeng. 2007, 104, 498–504. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr No. | Genes | Primer | Primer Sequence | Reference |
|---|---|---|---|---|
| 1 | ITS | ITS4F | TCCTCCGCTTATTGATATGC | White et al., 1990 [26] |
| ITS5R | GGAAGTAAAAGTCGTAACAAGG | White et al., 1990 [26] | ||
| 2 | EF 1-α | 983F | GCYCCYGGHCAYGGTGAYTTYAT | Rehner and Buckley, 2005 [27] |
| 2218R | ATGACACCRACRGCRACRGTYTG | Bischoff et al., 2009 [28] |
| Species | ITS | EF 1-α | ||||||
|---|---|---|---|---|---|---|---|---|
| Accession Number | Strain No. | Host | Location | Accession Number | Strain No. | Host | Location | |
| A. attenuatum | EF192939 | CNU-23 | Green peach aphid | Korea | KM283804 | KACC 42493 | / | Korea |
| A. attenuatum | MH558279 | MO315369 | Leaf Roller | USA | EF468782 | CBS 402.78 | / | USA |
| Akanthomyces muscarius | MH858370 | CBS 641.63 | / | Albania | KM283821 | CBS 143.62 | Trialeurodes vaporariorum | Korea |
| Akanthomyces longisporum | AJ292385 | IMI 021167 | Verticillium | United Kingdom | KM283819 | CBS 102072 | T. vaporariorum | Korea |
| Akanthomyces antillanum | AJ292392 | CBS 350.85 | Verticillium | United Kingdom | DQ522350 | CBS 350.85 | Animal pathogen | USA |
| Akanthomyces dimorphum | AJ292429 | CBS 363.86 | Verticillium | United Kingdom | LT220795 | TMSL132 | Soils | Portugal |
| Cordyceps javanica | JQ425659 | BCC24976 | Spider | Thailand | KY587206 | CHE-CNRCB 357 | Diaphorina citri | Mexico |
| Cordyceps javanica | JQ425660 | BCC26304 | Spider | Thailand | KY587208 | CHE-CNRCB 363 | D. citri | Mexico |
| Isolates | Regression Equation | LC50 (Conidia/mL) | 95% Confidence Limit |
|---|---|---|---|
| SCAUDCL-38 | Y = 0.352X–2.217 | 1.9 × 106 | (3.2 × 105–1.8 × 107) |
| SCAUDCL-56 | Y = 0.229X–1.651 | 1.5 × 107 | (1.2 × 106–9.5 × 1011) |
| Isolates | Regression Equation | LT50 (Days) | 95% Confidence Limit |
|---|---|---|---|
| SCAUDCL-38 | Y = 2.901X–5.910 | 3.52 | (2.84–4.75) |
| SCAUDCL-56 | Y = 2.832X–5.864 | 4.90 | (3.82–8.64) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, C.; Yang, B.; Wu, J.; Ali, S. Identification and Virulence Characterization of Two Akanthomyces attenuatus Isolates Against Megalurothrips usitatus (Thysanoptera: Thripidae). Insects 2019, 10, 168. https://doi.org/10.3390/insects10060168
Du C, Yang B, Wu J, Ali S. Identification and Virulence Characterization of Two Akanthomyces attenuatus Isolates Against Megalurothrips usitatus (Thysanoptera: Thripidae). Insects. 2019; 10(6):168. https://doi.org/10.3390/insects10060168
Chicago/Turabian StyleDu, Cailian, Bo Yang, Jianhui Wu, and Shaukat Ali. 2019. "Identification and Virulence Characterization of Two Akanthomyces attenuatus Isolates Against Megalurothrips usitatus (Thysanoptera: Thripidae)" Insects 10, no. 6: 168. https://doi.org/10.3390/insects10060168
APA StyleDu, C., Yang, B., Wu, J., & Ali, S. (2019). Identification and Virulence Characterization of Two Akanthomyces attenuatus Isolates Against Megalurothrips usitatus (Thysanoptera: Thripidae). Insects, 10(6), 168. https://doi.org/10.3390/insects10060168