Olfactory Preference of Drosophila suzukii Shifts between Fruit and Fermentation Cues over the Season: Effects of Physiological Status

 ,
,  and
and 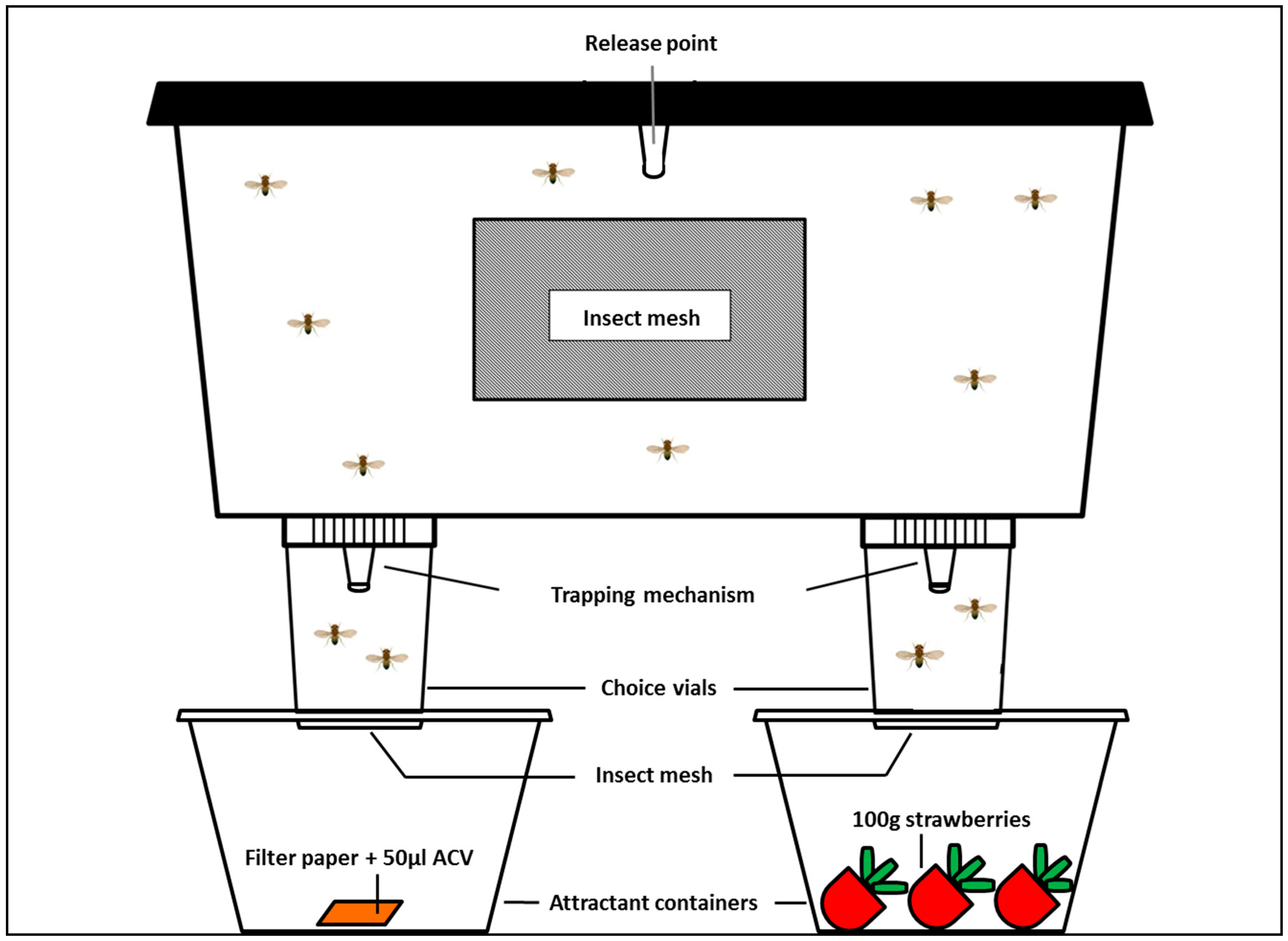{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
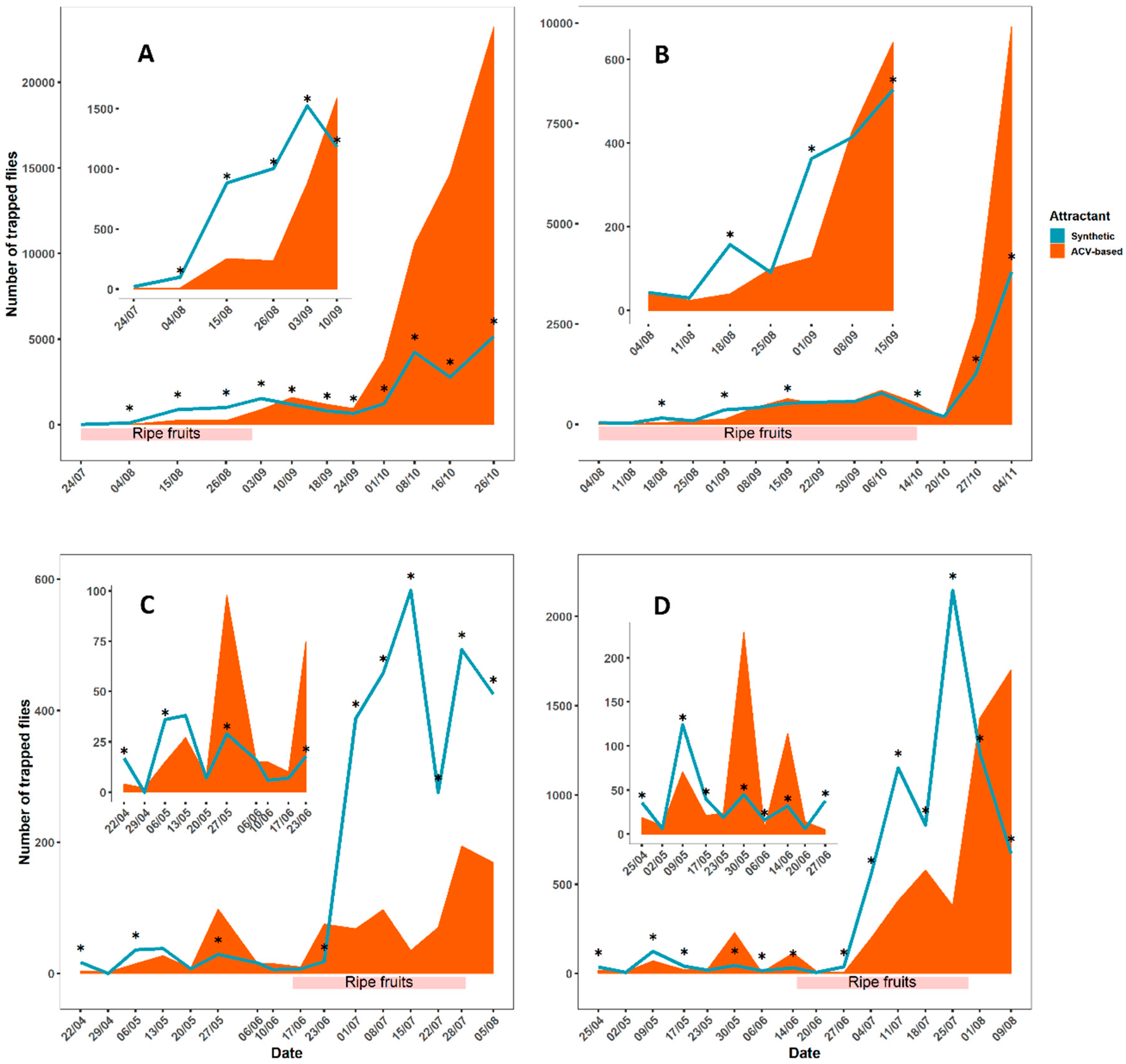
2.1. Field Comparisons of ACV Baits with Synthetic Lures
2.1.1. Experimental Sites
2.1.2. Experimental Design
2.1.3. Assessments
2.2. Olfactometer Experiments with ACV versus Strawberries: The Effect of Physiology
2.2.1. Insects
Stock Culture
Feeding Status
Mating Status
Reproductive Status
Seasonal Morphology
2.2.2. Experimental Set-up
Olfactometer
Procedure
2.3. Year-Round Field Olfactory Preference Experiment with ACV versus Strawberries
2.3.1. Experimental Site
2.3.2. Experimental Design
2.3.3. Assessments
2.4. Data Analysis
3. Results
3.1. Field Comparisons of ACV Baits with Synthetic Lures
3.2. Olfactometer Experiments with ACV versus Strawberries: The Effect of Physiology
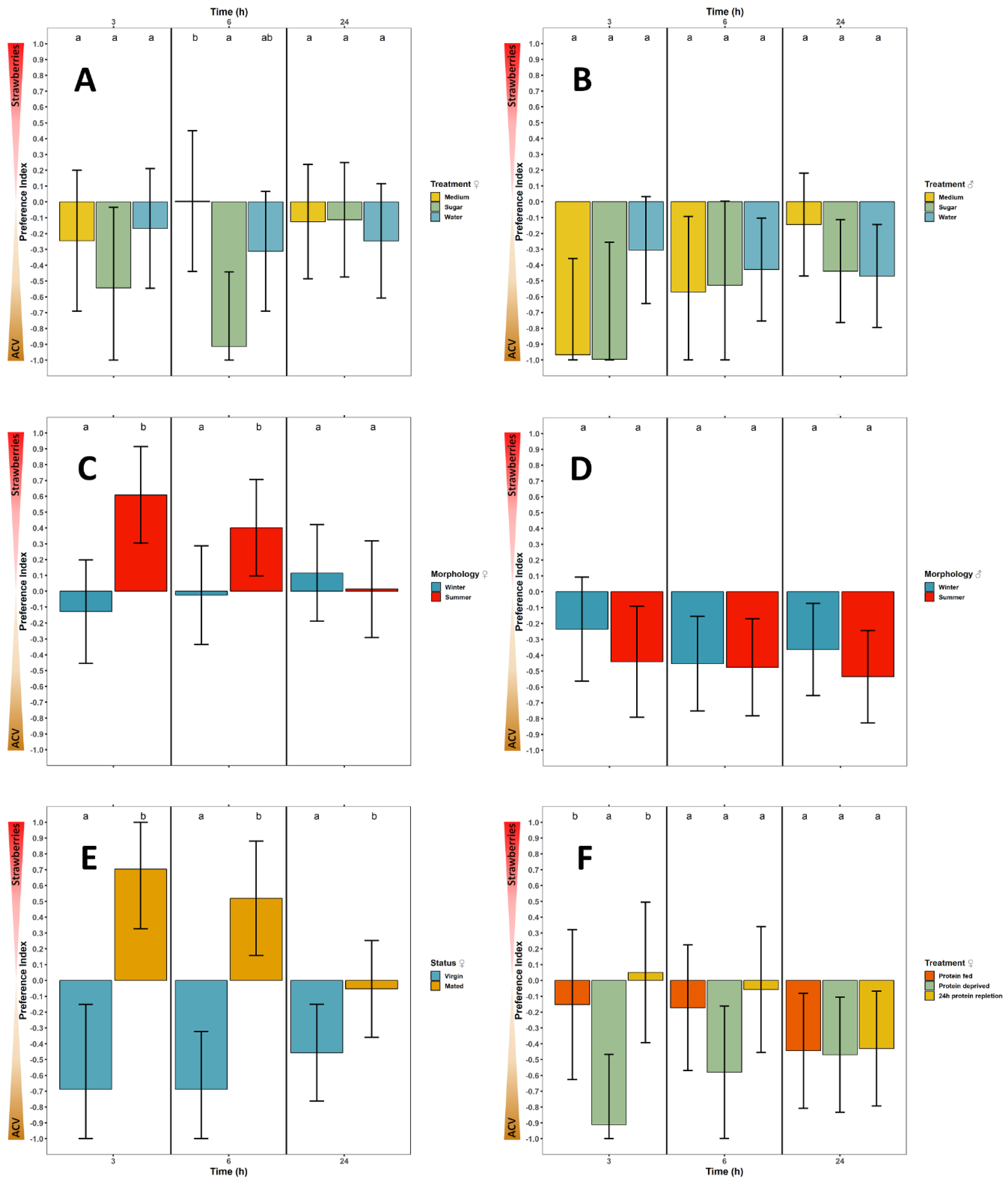
3.2.1. Feeding Status
3.2.2. Seasonal Morphology
3.2.3. Mating Status
3.2.4. Reproductive Status
3.2.5. Oogenic Status
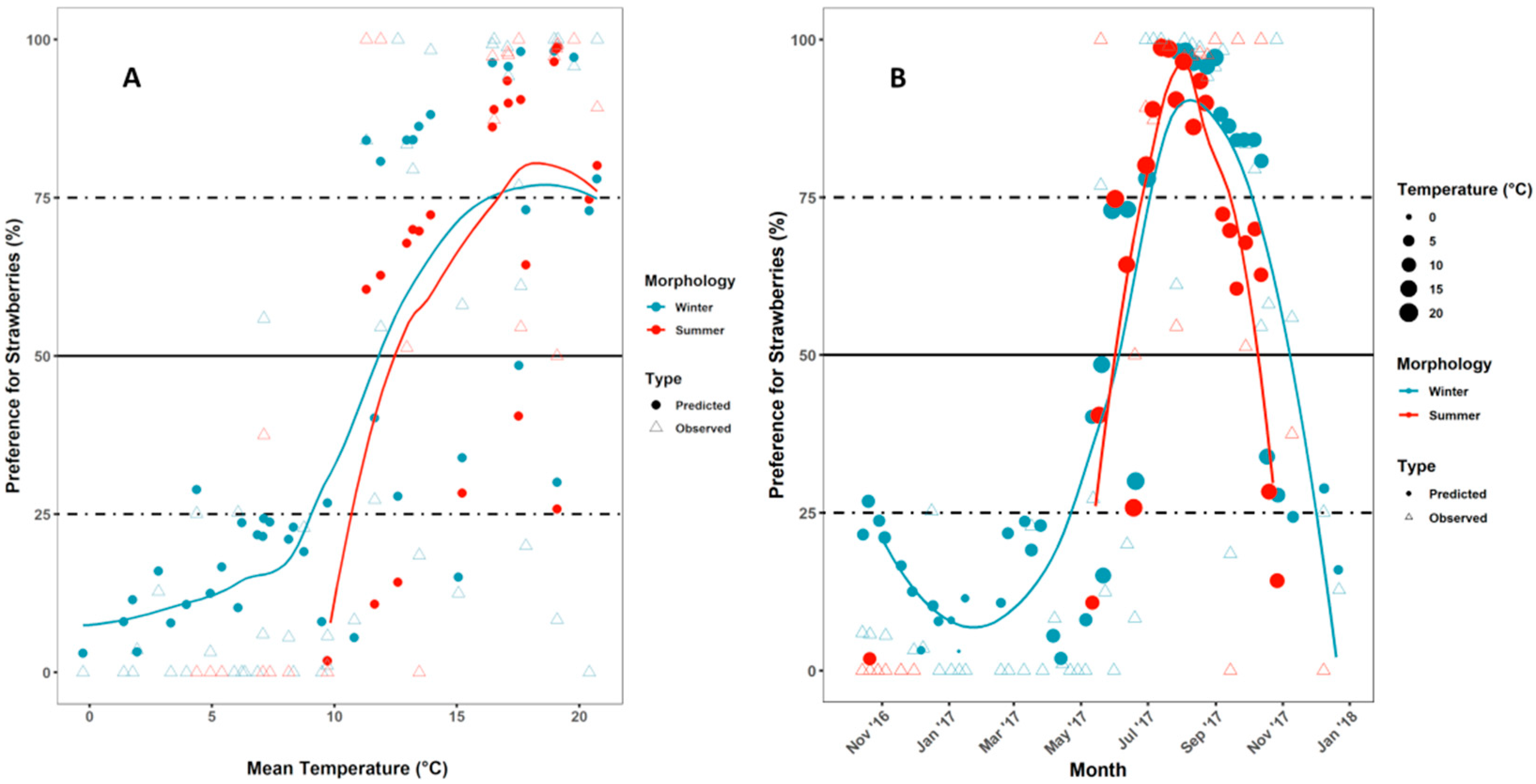
3.3. Year-Round Field Olfactory Preference Experiment with ACV versus Strawberries
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hamby, K.A.; Bolda, M.P.; Sheehan, M.E.; Zalom, F.G. Seasonal monitoring for Drosophila suzukii (Diptera: Drosophilidae) in California commercial raspberries. Environ. Entomol. 2014, 43, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Ørsted, I.V.; Ørsted, M. Species distribution models of the spotted wing Drosophila (Drosophila suzukii, Diptera: Drosophilidae) in its native and invasive range reveal an ecological niche shift. J. Appl. Ecol. 2019, 56, 423–435. [Google Scholar] [CrossRef]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Andreazza, F.; Bernardi, D.; Baronio, C.A.; Pasinato, J.; Nava, D.E.; Botton, M. Toxicities and effects of insecticidal toxic baits to control Drosophila suzukii and Zaprionus indianus (Diptera: Drosophilidae). Pest Manag. Sci. 2017, 73, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Belien, T.; Thys, T.; Fassotte, C.; Walrant, C.; Tomme, M.; Bolen, M.; Bylemans, D. Population dynamics of Drosophila suzukii (Diptera: Drosophilidae) in Belgium in 2013. Commun. Agric. Appl. Biol. Sci. 2014, 79, 169–175. [Google Scholar]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In Focus: Spotted wing Drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef]
- Mazzi, D.; Bravin, E.; Meraner, M.; Finger, R.; Kuske, S. Economic impact of the introduction and establishment of Drosophila suzukii on sweet cherry production in Switzerland. Insects 2017, 8, 18. [Google Scholar] [CrossRef]
- Mortelmans, J.; Casteels, H.; Belien, T. Drosophila suzukii (Diptera: Drosophilidae): A pest species new to Belgium. Belg. J. Zool. 2012, 142, 143–146. [Google Scholar]
- Stephens, A.R.; Asplen, M.K.; Hutchison, W.D.; Venette, R.C. Cold hardiness of winter-acclimated Drosophila suzukii (Diptera: Drosophilidae) adults. Environ. Entomol. 2015, 44, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, T.; Colinet, H. Basal tolerance to heat and cold exposure of the spotted wing Drosophila, Drosophila suzukii. PeerJ 2017, 5, e3112. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, A.K.; Loeb, G.M. Developmental acclimation of Drosophila suzukii (Diptera: Drosophilidae) and its effect on diapause and winter stress tolerance. Environ. Entomol. 2016, 45, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Stockton, D.G.; Wallingford, A.K.; Loeb, G.M. Phenotypic plasticity promotes overwintering survival in a globally invasive crop pest, Drosophila suzukii. Insects 2018, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- De Ro, M.; Devos, T.; Berkvens, N.; Casteels, H.; Goffin, J.; Belien, T.; De Clercq, P. Overwintering capacity of Drosophila suzukii (Diptera: Drosophilidae) in Belgium. IOBC-WPRS Bull. 2017, 123, 178–179. [Google Scholar]
- Chabert, S.; Allemand, R.; Poyet, M.; Eslin, P.; Gibert, P. Ability of European parasitoids (Hymenoptera) to control a new invasive Asiatic pest, Drosophila suzukii. Biol. Control 2012, 63, 40–47. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Grassi, A.; Dalton, D.T.; Miller, B.; Ouantar, M.; Loni, A.; Ioriatti, C.; Walton, V.M.; Anfora, G. First field records of Pachycrepoideus vindemiae as a parasitoid of Drosophila suzukii in European and Oregon small fruit production areas. Entomologia 2013, 1, 11–16. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Audsley, N. Efficacy of commercially available invertebrate predators against Drosophila suzukii. Insects 2014, 5, 952–960. [Google Scholar] [CrossRef]
- Papadopoulos, N.; Lux, S.A.; Köppler, K.; Belien, T. Invertebrate and vertebrate pests: Biology and management. In Cherries: Botany, Production and Uses; Quero-Garcia, J., Iezzoni, A., Lang, G., Pulawska, J., Eds.; CABI: Wallingford, UK, 2017; pp. 305–337. [Google Scholar]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the Asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef]
- Kenis, M.; Tonina, L.; Eschen, R.; van der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar] [CrossRef]
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The making of a pest: The evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. R. Soc. B 2014, 281, 20132840. [Google Scholar] [CrossRef] [PubMed]
- Alhmedi, A.; Clymans, R.; Van Kerckvoorde, V.; Bylemans, D.; Beliën, T. Preference and performance of Drosophila suzukii on Prunus species: A potential eco-friendly pest management tool. Crop Prot. 2019, 122, 35–41. [Google Scholar] [CrossRef]
- Adrion, J.R.; Kousathanas, A.; Pascual, M.; Burrack, H.J.; Haddad, N.M.; Bergland, A.O.; Machado, H.; Sackton, T.B.; Schlenke, T.A.; Watada, M.; et al. Drosophila suzukii: The genetic footprint of a recent, worldwide invasion. Mol. Biol. Evol. 2014, 31, 3148–3163. [Google Scholar] [CrossRef]
- Wiman, N.G.; Dalton, D.T.; Anfora, G.; Biondi, A.; Chiu, J.C.; Daane, K.M.; Gerdeman, B.; Gottardello, A.; Hamby, K.A.; Isaacs, R.; et al. Drosophila suzukii population response to environment and management strategies. J. Pest Sci. 2016, 89, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Van Timmeren, S.; Isaacs, R. Control of spotted wing Drosophila, Drosophila suzukii, by specific insecticides and by conventional and organic crop protection programs. Crop Prot. 2013, 54, 126–133. [Google Scholar] [CrossRef]
- Cha, D.H.; Hesler, S.P.; Wallingford, A.K.; Zaman, F.; Jentsch, P.; Nyrop, J.; Loeb, G.M. Comparison of commercial lures and food baits for early detection of fruit infestation risk by Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2018, 111, 645–652. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; Gut, L.J.; Miller, J.R. Estimating monitoring trap plume reach and trapping area for Drosophila suzukii (Diptera: Drosophilidae) in Michigan tart cherry. J. Econ. Entomol. 2018, 111, 1285–1289. [Google Scholar] [CrossRef]
- Jaffe, B.D.; Avanesyan, A.; Bal, H.K.; Feng, Y.; Grant, J.; Grieshop, M.J.; Lee, J.C.; Liburd, O.E.; Rhodes, E.; Rodriguez-Saona, C.; et al. Multistate comparison of attractants and the impact of fruit development stage on trapping Drosophila suzukii (Diptera: Drosophilidae) in raspberry and blueberry. Environ. Entomol. 2018, 47, 935–945. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Baroffio, C.A.; Richoz, P.; Fischer, S.; Kuske, S.; Linder, C.; Kehrli, P. Monitoring Drosophila suzukii in Switzerland in 2012. J. Berry Res. 2014, 4, 47–52. [Google Scholar] [CrossRef]
- Arnó, J.; Solà, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Briem, F.; Dominic, A.R.; Golla, B.; Hoffmann, C.; Englert, C.; Herz, A.; Vogt, H. Explorative data analysis of Drosophila suzukii trap catches from a seven-year monitoring program in Southwest Germany. Insects 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Mazzetto, F.; Pansa, M.G.; Ingegno, B.L.; Tavella, L.; Alma, A. Monitoring of the exotic fly Drosophila suzukii in stone, pome and soft fruit orchards in NW Italy. J. Asia Pac. Entomol. 2015, 18, 321–329. [Google Scholar] [CrossRef]
- Ioriatti, C.; Walton, V.; Dalton, D.; Anfora, G.; Grassi, A.; Maistri, S.; Mazzoni, V. Drosophila suzukii (Diptera: Drosophilidae) and its potential impact to wine grapes during harvest in two cool climate wine grape production regions. J. Econ. Entomol. 2015, 108, 1148–1155. [Google Scholar] [CrossRef]
- Tait, G.; Grassi, A.; Pfab, F.; Crava, C.M.; Dalton, D.T.; Magarey, R.; Ometto, L.; Vezzulli, S.; Rossi-Stacconi, M.V.; Gottardello, A.; et al. Large-scale spatial dynamics of Drosophila suzukii in Trentino, Italy. J. Pest Sci. 2018, 91, 1213–1224. [Google Scholar] [CrossRef]
- Wiman, N.G.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Burrack, H.J.; Chiu, J.C.; Daane, K.M.; Grassi, A.; Miller, B.; Tochen, S.; et al. Integrating temperature-dependent life table data into a matrix projection model for Drosophila suzukii population estimation. PLoS ONE 2014, 9, e106909. [Google Scholar] [CrossRef]
- Pelton, E.; Gratton, C.; Isaacs, R.; Van Timmeren, S.; Blanton, A.; Guédot, C. Earlier activity of Drosophila suzukii in high woodland landscapes but relative abundance is unaffected. J. Pest Sci. 2016, 89, 725–733. [Google Scholar] [CrossRef]
- Huang, J.; Gut, L.; Grieshop, M. Evaluation of food-based attractants for Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2017, 46, 878–884. [Google Scholar] [CrossRef]
- Wang, X.G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Harris, D.W.; Hamby, K.A.; Wilson, H.E.; Zalom, F.G. Seasonal monitoring of Drosophila suzukii (Diptera: Drosophilidae) in a mixed fruit production system. J. Asia. Pac. Entomol. 2014, 17, 857–864. [Google Scholar] [CrossRef]
- Drummond, F.; Ballman, E.; Collins, J. Population dynamics of spotted wing Drosophila (Drosophila suzukii Matsumara) in Maine wild blueberry (Vaccinium angustifolium Aiton). Preprints 2019, 2019050034. [Google Scholar]
- Santoiemma, G.; Trivellato, F.; Caloi, V.; Mori, N.; Marini, L. Habitat preference of Drosophila suzukii across heterogeneous landscapes. J. Pest Sci. 2019, 92, 485–494. [Google Scholar] [CrossRef]
- Tonina, L.; Mori, N.; Sancassani, M.; Dall’Ara, P.; Marini, L. Spillover of Drosophila suzukii between noncrop and crop areas: Implications for pest management. Agric. For. Entomol. 2018, 20, 575–581. [Google Scholar] [CrossRef]
- Burrack, H.J.; Asplen, M.; Bahder, L.; Collins, J.; Drummond, F.A.; Guédot, C.; Isaacs, R.; Johnson, D.; Blanton, A.; Lee, J.C.; et al. Multistate comparison of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae) in blueberries and caneberries. Environ. Entomol. 2015, 44, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, K.R. Advances in the chemical ecology of the spotted wing Drosophila (Drosophila suzukii) and its applications. J. Chem. Ecol. 2018, 44, 922–939. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.A.; Luo, N.; Yamaguchi, A.; Kapahi, P. A role for S6 kinase and serotonin in postmating dietary switch and balance of nutrients in D. melanogaster. Curr. Biol. 2010, 20, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Kim, J.S.; Min, K.J. Sexual dimorphism in nutrient intake and life span is mediated by mating in Drosophila melanogaster. Anim. Behav. 2013, 86, 987–992. [Google Scholar] [CrossRef]
- Walker, S.J.; Corrales-Carvajal, V.M.; Ribeiro, C. Postmating circuitry modulates salt taste processing to increase reproductive output in Drosophila. Curr. Biol. 2015, 25, 2621–2630. [Google Scholar] [CrossRef]
- Toshima, N.; Tanimura, T. Taste preference for amino acids is dependent on internal nutritional state in Drosophila melanogaster. J. Exp. Biol. 2012, 215, 2827–2832. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, S.; Becher, P.G.; Hansson, B.S.; Witzgall, P. Attraction of Drosophila melanogaster males to food-related and fly odours. J. Insect Physiol. 2012, 58, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.G.; Bengtsson, M.; Hansson, B.S.; Witzgall, P. Flying the fly: Long-range flight behavior of Drosophila melanogaster to attractive odors. J. Chem. Ecol. 2010, 36, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Guo, W.-Y.; Muhammad, S.A.; Chen, R.-R.; Mu, L.-L.; Li, G.-Q. Mating experience and food deprivation modulate odor preference and dispersal in Drosophila melanogaster males. J. Insect Sci. 2016, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, D.M.; Leach, H.L.; Xu, P.; Dong, K.; Isaacs, R.; Gut, L.J. Comparative antennal and behavioral responses of summer and winter morph Drosophila suzukii (Diptera: Drosophilidae) to ecologically relevant Volatiles. Environ. Entomol. 2018, 47, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Mori, B.A.; Whitener, A.B.; Leinweber, Y.; Revadi, S.; Beers, E.H.; Witzgall, P.; Becher, P.G. Enhanced yeast feeding following mating facilitates control of the invasive fruit pest Drosophila suzukii. J. Appl. Ecol. 2017, 54, 170–177. [Google Scholar] [CrossRef]
- Wong, J.S.; Wallingford, A.K.; Loeb, G.M.; Lee, J.C. Physiological status of Drosophila suzukii (Diptera: Drosophilidae) affects their response to attractive odours. J. Appl. Entomol. 2018, 142, 473–482. [Google Scholar] [CrossRef]
- Rendon, D.; Buser, J.; Tait, G.; Lee, J.C.; Walton, V.M. Survival and fecundity parameters of two Drosophila suzukii (Diptera: Drosophilidae) morphs on variable diet under suboptimal temperatures. J. Insect Sci. 2018, 18, 8. [Google Scholar] [CrossRef]
- Young, Y.; Buckiewicz, N.; Long, T.A.F. Nutritional geometry and fitness consequences in Drosophila suzukii, the spotted-wing Drosophila. Ecol. Evol. 2018, 8, 2842–2851. [Google Scholar] [CrossRef]
- Plantamp, C.; Estragnat, V.; Fellous, S.; Desouhant, E.; Gibert, P. Where and what to feed? Differential effects on fecundity and longevity in the invasive Drosophila suzukii. Basic Appl. Ecol. 2017, 19, 56–66. [Google Scholar] [CrossRef]
- Shearer, P.W.; West, J.D.; Walton, V.M.; Brown, P.H.; Svetec, N.; Chiu, J.C. Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecol. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, A.K.; Rice, K.B.; Leskey, T.C.; Loeb, G.M. Overwintering behavior of Drosophila suzukii, and potential springtime diets for egg maturation. Environ. Entomol. 2018, 47, 1266–1271. [Google Scholar] [CrossRef] [PubMed]
- Panel, A.D.C.; Zeeman, L.; van der Sluis, B.J.; van Elk, P.; Pannebakker, B.A.; Wertheim, B.; Helsen, H.H.M. Overwintered Drosophila suzukii are the main source for infestations of the first fruit crops of the season. Insects 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Grassi, A.; Gottardello, A.; Dalton, D.T.; Tait, G.; Rendon, D.; Ioriatti, C.; Gibeaut, D.; Rossi-Stacconi, M.V.; Walton, V.M. Seasonal reproductive biology of Drosophila suzukii (Diptera: Drosophilidae) in temperate climates. Environ. Entomol. 2018, 47, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Royal Observatory of Belgium. Ephemerides. 2017. Available online: https://www.astro.oma.be/en/ (accessed on 8 May 2019).
- Krause Pham, C.; Ray, A. Conservation of olfactory avoidance in Drosophila species and identification of repellents for Drosophila suzukii. Sci. Rep. 2015, 5, 11527. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Zhang, A.; Angeli, S.; Abubeker, S.; Michel, C.; Feng, Y.; Rodriguez-Saona, C. Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 2015, 44, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Revadi, S.; Vitagliano, S.; Rossi-Stacconi, M.V.; Ramasamy, S.; Mansourian, S.; Carlin, S.; Vrhovsek, U.; Becher, P.G.; Mazzoni, V.; Rota-Stabelli, O.; et al. Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Solieri, L.; Giudici, P. Vinegars of the world. In Vinegars of the World; Solieri, L., Giudici, P., Eds.; Springer: Milan, Italy, 2009; pp. 1–14. [Google Scholar]
- Chandler, J.A.; James, P.M.; Jospin, G.; Lang, J.M. The bacterial communities of Drosophila suzukii collected from undamaged cherries. PeerJ 2014, 2, e474. [Google Scholar] [CrossRef]
- Mazzetto, F.; Gonella, E.; Crotti, E.; Vacchini, V.; Syrpas, M.; Pontini, M.; Mangelinckx, S.; Daffonchio, D.; Alma, A. Olfactory attraction of Drosophila suzukii by symbiotic acetic acid bacteria. J. Pest Sci. 2016, 89, 783–792. [Google Scholar] [CrossRef]
- Hamby, K.A.; Becher, P.G. Current knowledge of interactions between Drosophila suzukii and microbes, and their potential utility for pest management. J. Pest Sci. 2016, 89, 621–630. [Google Scholar] [CrossRef]
- Ioriatti, C.; Guzzon, R.; Anfora, G.; Ghidoni, F.; Mazzoni, V.; Villegas, T.R.; Dalton, D.T.; Walton, V.M. Drosophila suzukii (Diptera: Drosophilidae) contributes to the development of sour rot in grape. J. Econ. Entomol. 2018, 111, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Sañudo, I.; Simonato, M.; Squartini, A.; Mori, N.; Marri, L.; Mazzon, L. Metagenomic analysis reveals changes of the Drosophila suzukii microbiota in the newly colonized regions. Insect Sci. 2018, 25, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Vacchini, V.; Gonella, E.; Crotti, E.; Prosdocimi, E.M.; Mazzetto, F.; Chouaia, B.; Callegari, M.; Mapelli, F.; Mandrioli, M.; Alma, A.; et al. Bacterial diversity shift determined by different diets in the gut of the spotted wing fly Drosophila suzukii is primarily reflected on acetic acid bacteria. Environ. Microbiol. Rep. 2017, 9, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Fountain, M.T.; Bennett, J.; Cobo-Medina, M.; Conde Ruiz, R.; Deakin, G.; Delgado, A.; Harrison, R.; Harrison, N. Alimentary microbes of winter-form Drosophila suzukii. Insect Mol. Biol. 2018, 27, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Bing, X.; Gerlach, J.; Loeb, G.; Buchon, N. Nutrient-dependent impact of microbes on Drosophila suzukii development. MBio 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Scheidler, N.H.; Liu, C.; Hamby, K.A.; Zalom, F.G.; Syed, Z. Volatile codes: Correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [PubMed]
- Bellutti, N.; Gallmetzer, A.; Innerebner, G.; Schmidt, S.; Zelger, R.; Koschier, E.H. Dietary yeast affects preference and performance in Drosophila suzukii. J. Pest Sci. 2018, 91, 651–660. [Google Scholar] [CrossRef]
- Pavlova, A.K.; Dahlmann, M.; Hauck, M.; Reineke, A. Laboratory bioassays with three different substrates to test the efficacy of insecticides against various stages of Drosophila suzukii (Diptera: Drosophilidae). J. Insect Sci. 2017, 17, 8. [Google Scholar] [CrossRef]
- Briem, F.; Zeisler, C.; Guenay, Y.; Staudacher, K.; Vogt, H.; Traugott, M. Identifying plant DNA in the sponging—Feeding insect pest Drosophila suzukii. J. Pest Sci. 2018, 91, 985–994. [Google Scholar] [CrossRef]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef]
- Dekker, T.; Revadi, S.; Mansourian, S.; Ramasamy, S.; Lebreton, S.; Becher, P.G.; Angeli, S.; Rota-Stabelli, O.; Anfora, G. Loss of Drosophila pheromone reverses its role in sexual communication in Drosophila suzukii. Proc. R. Soc. B. 2015, 282, 20143018. [Google Scholar] [CrossRef] [PubMed]
- Snellings, Y.; Herrera, B.; Wildemann, B.; Beelen, M.; Zwarts, L.; Wenseleers, T.; Callaerts, P. The role of cuticular hydrocarbons in mate recognition in Drosophila suzukii. Sci. Rep. 2018, 8, 4996. [Google Scholar] [CrossRef] [PubMed]
- Revadi, S.; Lebreton, S.; Witzgall, P.; Anfora, G.; Dekker, T.; Becher, P.G. Sexual behavior of Drosophila suzukii. Insects 2015, 6, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Connolly, K.J. Locomotor activity in Drosophila as a function of food deprivation. Nature 1966, 209, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Sayin, S.; De Backer, J.-F.; Wosniack, M.E.; Lewis, L.P.; Siju, K.P.; Frisch, L.-M.; Schlegel, P.; Edmondson-Stait, A.; Sharifi, N.; Fisher, C.B.; et al. A neural circuit arbitrates between perseverance and withdrawal in hungry Drosophila. bioRxiv 2018. [Google Scholar] [CrossRef]
- Turelli, M.; Hoffmann, A.A. Effects of starvation and experience on the response of Drosophila to alternative resources. Oecologia 1988, 77, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Dickson, B.J. Sex peptide receptor and neuronal TOR/S6K signaling modulate nutrient balancing in Drosophila. Curr. Biol. 2010, 20, 1000–1005. [Google Scholar] [CrossRef]
- Swoboda-Bhattarai, K.A.; McPhie, D.R.; Burrack, H.J. Reproductive status of Drosophila suzukii (Diptera: Drosophilidae) females influences attraction to fermentation-based baits and ripe fruits. J. Econ. Entomol. 2017, 110, 1648–1652. [Google Scholar] [CrossRef]
- Wyman, R. The temporal stability of the Drosophila oocyte. J. Embryol. Exp. Morph. 1979, 50, 137–144. [Google Scholar]
- Boulétreau-Merle, J.; Fouillet, P. How to overwinter and be a founder: Egg-retention phenotypes and mating status in Drosophila melanogaster. Evol. Ecol. 2002, 16, 309–332. [Google Scholar] [CrossRef]
- Akhund-Zade, J.; Bergland, A.O.; Crowe, S.O.; Unckless, R.L. The genetic basis of natural variation in Drosophila (Diptera: Drosophilidae) virgin egg retention. J. Insect Sci. 2017, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, A.K.; Hesler, S.P.; Cha, D.H.; Loeb, G.M. Behavioral response of spotted-wing Drosophila, Drosophila suzukii Matsumura, to aversive odors and a potential oviposition deterrent in the field. Pest Manag. Sci. 2016, 72, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, A.K.; Cha, D.H.; Linn, C.E.; Wolfin, M.S.; Loeb, G.M. Robust manipulations of pest insect behavior using repellents and practical application for integrated pest management. Environ. Entomol. 2017, 46, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Zhao, C.; Zheng, W.; Wang, W.; Zhao, X.; Jiao, L. Analyzing strawberry spoilage via its volatile compounds using longpath fourier transform infrared spectroscopy. Sci. Rep. 2013, 3, 2585. [Google Scholar] [CrossRef] [PubMed]
- Vandendriessche, T.; Keulemans, J.; Geeraerd, A.; Nicolai, B.M.; Hertog, M.L. Evaluation of fast volatile analysis for detection of Botrytis cinerea infections in strawberry. Food Microbiol. 2012, 32, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Tochen, S.; Walton, V.M.; Lee, J.C. Impact of floral feeding on adult Drosophila suzukii survival and nutrient status. J. Pest Sci. 2016, 89, 793–802. [Google Scholar] [CrossRef]
- Bal, H.K.; Adams, C.; Grieshop, M. Evaluation of off-season potential breeding sources for spotted wing Drosophila (Drosophila suzukii Matsumura) in Michigan. J. Econ. Entomol. 2017, 110, 2466–2470. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Bruton, R.; Park, A.; Zhang, A. Identification of attractive blend for spotted wing drosophila, Drosophila suzukii, from apple juice. J. Pest Sci. 2018, 91, 1251–1267. [Google Scholar] [CrossRef]
- Cha, D.H.; Hesler, S.P.; Cowles, R.S.; Vogt, H.; Loeb, G.M.; Landolt, P.J. Comparison of a synthetic chemical lure and standard fermented baits for trapping Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2013, 42, 1052–1060. [Google Scholar] [CrossRef]
- Lux, S.A.; Wnuk, A.; Vogt, H.; Belien, T.; Spornberger, A.; Studnicki, M. Validation of individual-based Markov-like stochastic process model of insect behavior and a “virtual farm” concept for enhancement of site-specific IPM. Front. Physiol. 2016, 7, 1–20. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clymans, R.; Van Kerckvoorde, V.; Bangels, E.; Akkermans, W.; Alhmedi, A.; De Clercq, P.; Beliën, T.; Bylemans, D. Olfactory Preference of Drosophila suzukii Shifts between Fruit and Fermentation Cues over the Season: Effects of Physiological Status. Insects 2019, 10, 200. https://doi.org/10.3390/insects10070200
Clymans R, Van Kerckvoorde V, Bangels E, Akkermans W, Alhmedi A, De Clercq P, Beliën T, Bylemans D. Olfactory Preference of Drosophila suzukii Shifts between Fruit and Fermentation Cues over the Season: Effects of Physiological Status. Insects. 2019; 10(7):200. https://doi.org/10.3390/insects10070200
Chicago/Turabian StyleClymans, Rik, Vincent Van Kerckvoorde, Eva Bangels, Wannes Akkermans, Ammar Alhmedi, Patrick De Clercq, Tim Beliën, and Dany Bylemans. 2019. "Olfactory Preference of Drosophila suzukii Shifts between Fruit and Fermentation Cues over the Season: Effects of Physiological Status" Insects 10, no. 7: 200. https://doi.org/10.3390/insects10070200
APA StyleClymans, R., Van Kerckvoorde, V., Bangels, E., Akkermans, W., Alhmedi, A., De Clercq, P., Beliën, T., & Bylemans, D. (2019). Olfactory Preference of Drosophila suzukii Shifts between Fruit and Fermentation Cues over the Season: Effects of Physiological Status. Insects, 10(7), 200. https://doi.org/10.3390/insects10070200