The Dominance Hierarchy of Wood-Eating Termites from China


Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Method
2.2. Experiment 1: Equal Number of Termites
2.3. Experiment 2: Equal Biomass of Termites
2.4. Experiment 3: Chemical Detection of Species
2.5. Field Survey
2.6. Analysis
3. Results
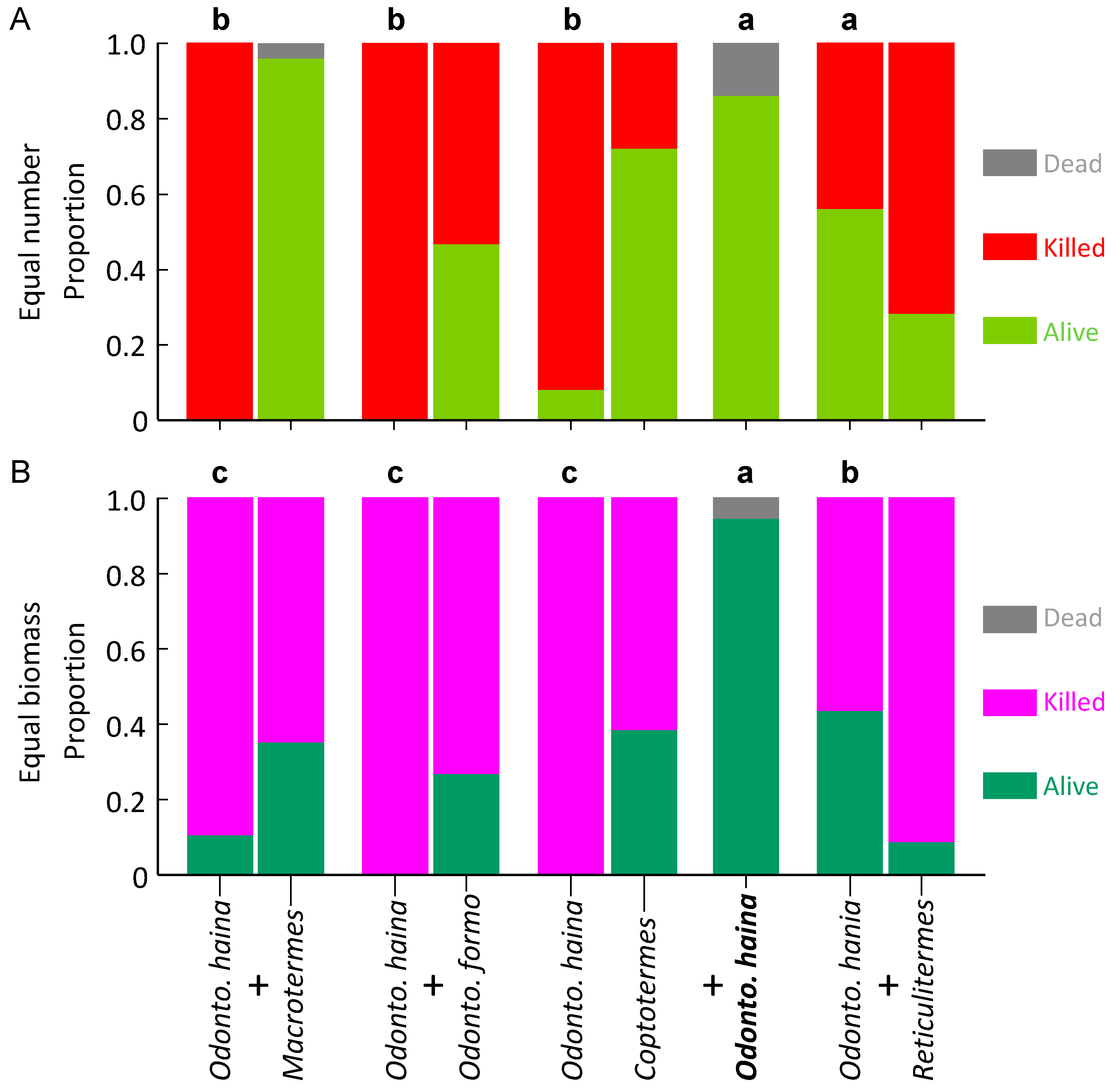
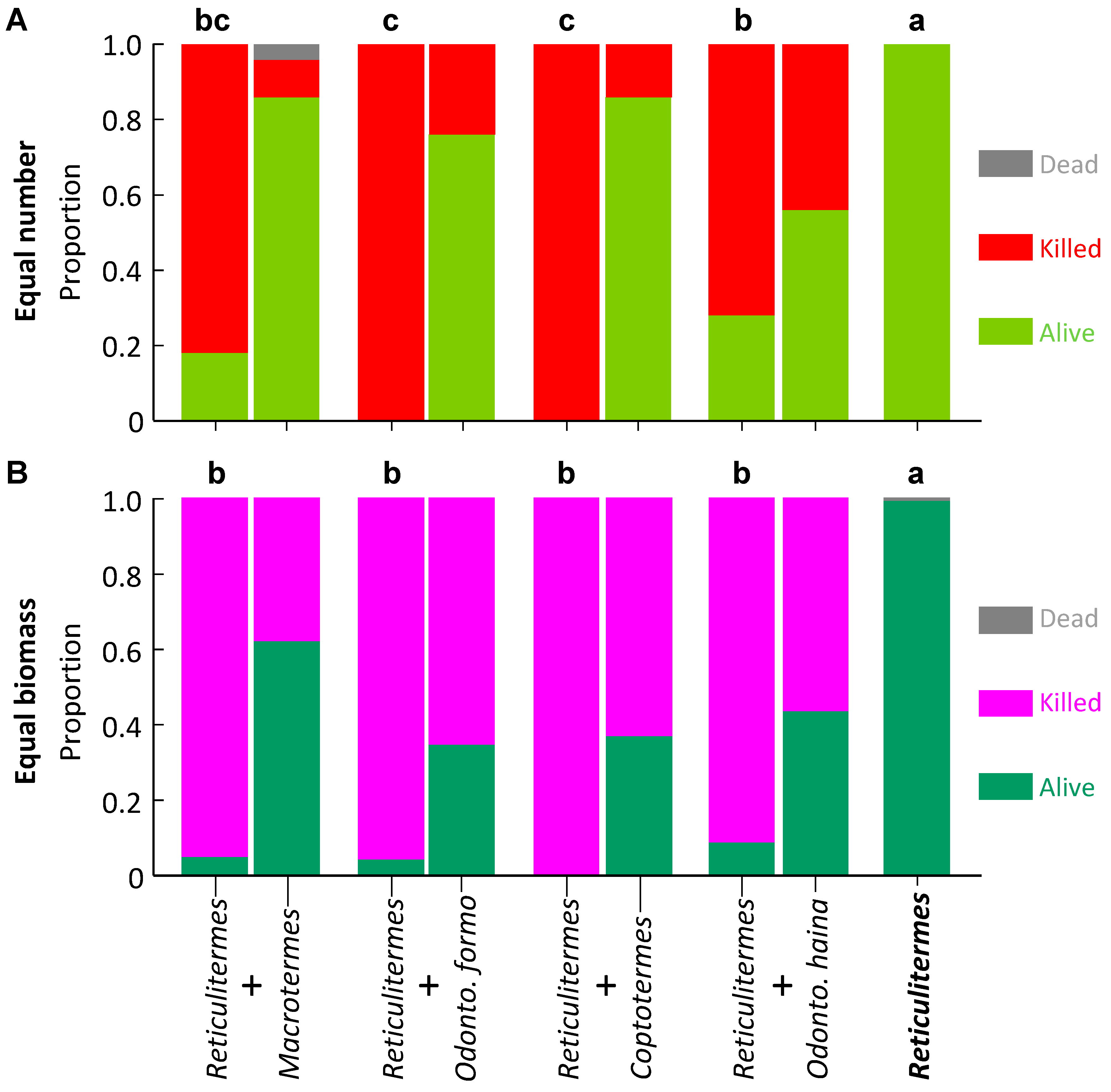
3.1. Experiment 1: Equal Number of Termites
3.2. Experiment 2: Equal Biomass of Termites
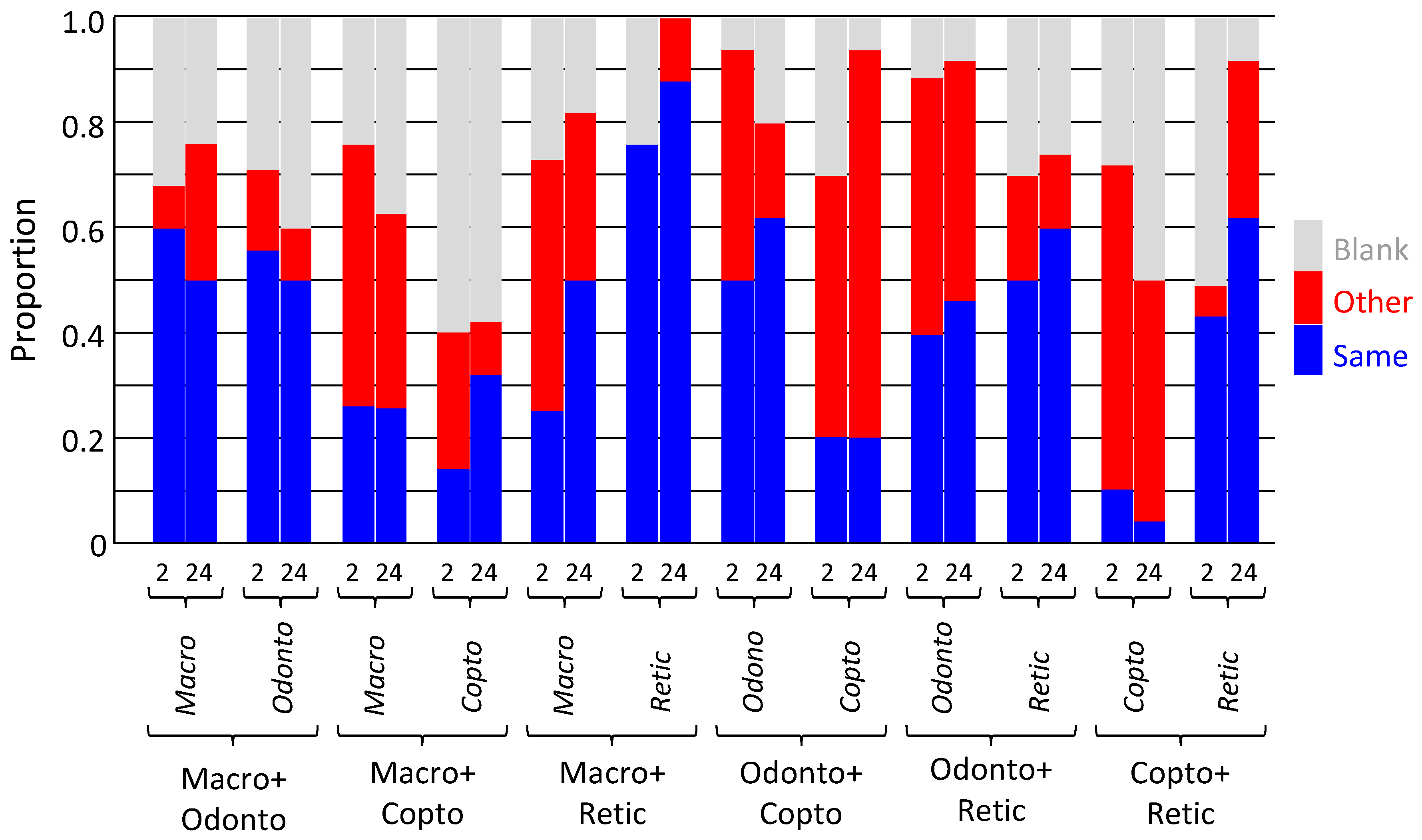
3.3. Experiment 3: Chemical Detection of Species
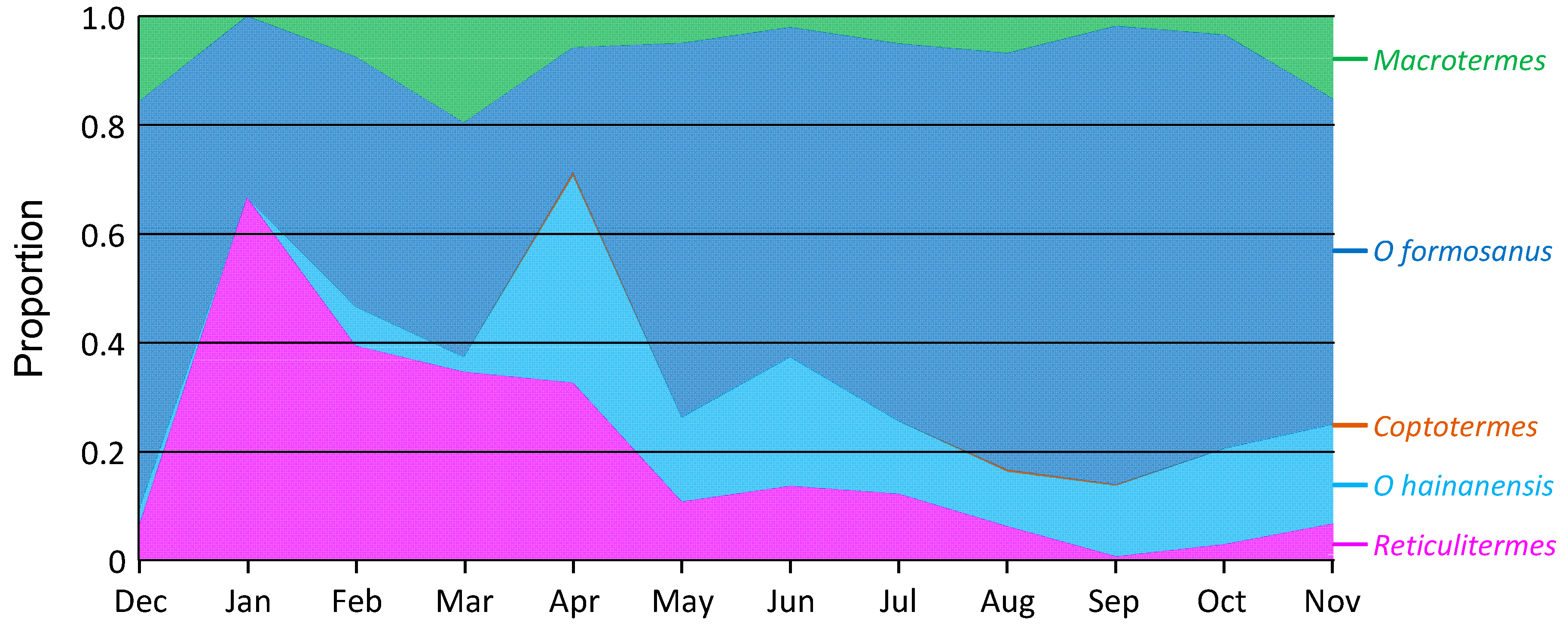
3.4. Field Survey
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C.D. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. Murray, London, UK, 1859; p. 140. Available online: http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=1 (accessed on 13 July 2019).
- Gause, G.F. The Struggle for Existence. The Williams & Wilkins Company: Baltimore, MD, USA, 1934. Available online: https://doi.org/10.5962/bhl.title.4489 (accessed on 13 July 2019).
- Elton, C. Competition and the structure of ecological communities. J. Anim. Ecol. 1946, 15, 54–68. [Google Scholar] [CrossRef]
- Park, T.; Gregg, E.V.; Lutherman, C.Z. Studies in population pysiology. X. Interspecific competition in populations of granary beetles. Physiol. Zool. 1941, 14, 395–430. [Google Scholar] [CrossRef]
- Crombie, A.C. On competition between different species of graminivorous insects. Proc. R. Soc. B 1945, 132, 362–395. [Google Scholar] [CrossRef] [Green Version]
- Crombie, A.C. Further experiments on insect competition. Proc. R. Soc. B 1946, 133, 76–109. [Google Scholar] [CrossRef]
- Park, T. Interspecies competition in populations of Trilobium confusum Duval and Trilobium castaneum Herbst. Ecol. Monogr. 1948, 18, 265–307. [Google Scholar] [CrossRef]
- Schoener, T.W. Field experiments on interspecific competition. Am. Nat. 1983, 122, 240–285. [Google Scholar] [CrossRef]
- Carothers, J.H.; Jaksić, F.M. Time as a niche difference: The role of interference competition. Oikos 1984, 42, 403–406. [Google Scholar] [CrossRef]
- Davidson, D.W. An experimental study of diffuse competition in harvester ants. Am. Nat. 1985, 125, 500–506. [Google Scholar] [CrossRef]
- Kronfeld-Schor, N.; Dayan, T. Partitioning of time as an ecological resource. Ann. Rev. Ecol. Evol. Syst. 2003, 34, 153–181. [Google Scholar] [CrossRef]
- Evans, T.A.; Inta, R.; Lai, J.C.S.; Prueger, S.; Foo, N.W.; Fu, E.W.; Lenz, M. Termites eavesdrop to avoid competitors. Proc. R. Soc. B 2009, 276, 4035–4041. [Google Scholar] [CrossRef] [Green Version]
- Eggleton, P.; Tayasu, I. Feeding groups, lifetypes and the global ecology of termites. Ecol. Res. 2001, 16, 941–960. [Google Scholar] [CrossRef]
- Ohkuma, M.; Brune, A. Diversity, structure, and evolution of the termite gut microbial community. In Biology of Termites: A Modern Synthesis; Bignell, D., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, Netherlands, 2011; pp. 413–438. ISBN 978-90-481-3976-7. [Google Scholar]
- Brune, A.; Dietrich, C. The gut microbiota of termites: Digesting the diversity in the light of ecology and evolution. Ann. Rev. Microbiol. 2015, 69, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Bultman, J.D.; Southwell, C.R. Natural resistance of tropical American woods to terrestrial wood-destroying organisms. Biotropica 1976, 8, 71–95. [Google Scholar] [CrossRef]
- Wood, T.G.; Sands, W.A. The role of termites in ecosystems. In Production Ecology of Ants and Termites; Brian, M.V., Ed.; Cambridge University Press: Cambridge, UK, 1978; pp. 245–292. ISBN 0 521 215196. [Google Scholar]
- Scheffer, T.C.; Morrell, J.J. Natural Durability of Wood: A Worldwide Checklist of Species. Oregon State University, Forest Research Laboratory, Technical Report 22. 1998. Available online: https://ir.library.oregonstate.edu/concern/technical_reports/dz010r37p (accessed on 13 July 2019).
- Nel, J.J.C. Aggressive behaviour of the harvester termites Hodotermes mossambicus (Hagen) and Trinervitermes trinervoides (Sjöstedt). Insectes Soc. 1968, 15, 145–156. [Google Scholar] [CrossRef]
- Thorne, B.L.; Haverty, M.I. Review of intracolony, intraspecific and interspecific agonism in termites. Sociobiology 1991, 19, 115–145. [Google Scholar]
- Shelton, T.G.; Grace, J.K. Review of agonistic behaviors in the Isoptera. Sociobiology 1996, 28, 155–176. [Google Scholar]
- Adams, E.S. Nest-mate recognition based on heritable odors in the termite Microcerotermes arboreus. Proc. Natl. Acad. Sci. USA 1991, 88, 2031–2034. [Google Scholar] [CrossRef]
- Haverty, M.I.; Thorne, B.L. Agonistic behavior correlated with hydrocarbon phenotypes in dampwood termites, Zootermopsis (Isoptera: Termopsidae). J. Insect Behav. 1989, 2, 523–543. [Google Scholar] [CrossRef]
- Bagneres, A.-G.; Killian, A.; Clement, J.-L.; Lange, C. Interspecific recognition among termites of the genus Reticulitermes: Evidence for a role for the cuticular hydrocarbons. J. Chem. Ecol. 1991, 17, 2397–2420. [Google Scholar] [CrossRef]
- Bagnères, A.-G.; Hanus, R. Communication and social regulation in termites. In Social Recognition in Invertebrates; Aquiloni, L., Tricarico, E., Eds.; Springer: Cham, Switzerland, 2015; pp. 193–248. ISBN 978-3-319-17598-0. [Google Scholar]
- Gontijo, T.A.; Domingos, D.J. Guild distribution of some termites from cerrado vegetation in south-east Brazil. J. Trop. Ecol. 1991, 7, 523–529. [Google Scholar] [CrossRef]
- Korb, J.; Linsenmair, K.E. The causes of spatial patterning of mounds of a fungus-cultivating termite: Results from nearest-neighbour analysis and ecological studies. Oecologia 2001, 127, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.B.; Levick, S.R.; Asner, G.P.; Robertson, M.P.; van Rensburg, B.J.; Parr, C.L. Spatial variability and abiotic determinants of termite mounds throughout a savanna catchment. Ecography 2014, 37, 852–862. [Google Scholar] [CrossRef]
- Huang, F.S.; Zhu, S.M.; Ping, Z.M.; He, X.S.; Li, G.X.; Gao, D.R. Isoptera. In Fauna Sinica, Insecta; Science Press: Beijing, China, 2000; Volume 17. (In Chinese) [Google Scholar]
- Cowie, R.H.; Logan, J.W.M.; Wood, T.G. Termite (Isoptera) damage and control in tropical forestry with special reference to Africa and Indo-Malaysia: A review. Bull. Ent. Res. 1989, 79, 173–184. [Google Scholar] [CrossRef]
- Constantino, R. The pest termites of South America: Taxonomy, distribution and status. J. Appl. Ent. 2002, 126, 355–365. [Google Scholar] [CrossRef]
- Rouland-Lefèvre, C. Termites as pests of agriculture. In Biology of Termites: A Modern Synthesis; Bignell, D., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 499–517. ISBN 978-90-481-3976-7. [Google Scholar]
- Polizzi, J.M.; Forschler, B.T. Factors that affect aggression among the worker caste of Reticulitermes spp. subterranean termites (Isoptera: Rhinotermitidae). J. Insect Behav. 1999, 12, 133–146. [Google Scholar] [CrossRef]
- Wong, N.; Lee, C.-Y. Intra- and interspecific agonistic behavior of the subterranean termite Microcerotermes crassus (Isoptera: Termitidae). J. Econ. Entomol. 2010, 103, 1754–1760. [Google Scholar] [CrossRef]
- Rojo, M.J.A.; Acda, M.N. Interspecific agonistic behavior of Macrotermes gilvus (Isoptera: Termitidae): Implication on termite baiting in the Philippines. J. Insect Behav 2016, 273–282. [Google Scholar] [CrossRef]
- Simkovic, V.; Thompson, G.J.; McNeil, J.N. Testing for aggression and nestmate recognition in the Eastern subterranean termite (Reticulitermes flavipes). Insectes Sociaux 2018, 65, 281–288. [Google Scholar] [CrossRef]
- Thorne, B.L. Termite-termite interactions: Workers as an agonistic caste. Psyche 1982, 89, 133–150. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Osbrink, W.L.A. Bioassay design and length of time in the laboratory affect intercolonial interactions of the Formosan subterranean termite (Isoptera, Rhinotermitidae). Insectes Soc. 2009, 56, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, R.; Vepsäläinen, K. A competition hierarchy among boreal ants: Impact on resource partitioning and community structure. Oikos 1988, 51, 135–155. [Google Scholar] [CrossRef]
- Andersen, A.N.; Patel, A.D. Meat ants as dominant members of Australian ant communities: An experimental test of their influence on the foraging success and forager abundance of other species. Oecologia 1994, 98, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, X.; Retana, J.; Manzaneda, A. The role of competition by dominants and temperature in the foraging of subordinate species in Mediterranean ant communities. Oecologia 1998, 117, 404–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wcislo, W.T.; Arneson, L.; Roesch, K.; Gonzalez, V.; Smith, A.; Fernández, H. The evolution of nocturnal behaviour in sweat bees, Megalopta genalis and M. ecuadoria (Hymenoptera: Halictidae): An escape from competitors and enemies? Biol. J. Linn. Soc. 2004, 83, 377–387. [Google Scholar] [CrossRef]
- Wcislo, W.T.; Tierney, S.M. Behavioural environments and niche construction: The evolution of dim-light foraging in bees. Biol. Rev. 2009, 84, 19–37. [Google Scholar] [CrossRef]
- Shavit, O.; Dafni, A.; Ne’eman, G. Competition between honeybees (Apis mellifera) and native solitary bees in the Mediterranean region of Israel—Implications for conservation. Israel J. Plant Sci. 2009, 57, 171–183. [Google Scholar] [CrossRef]
- Smith, A.R.; Kitchen, S.M.; Toney, R.M.; Ziegler, C. Is nocturnal foraging in a tropical bee an escape from interference competition? J. Insect Sci. 2017, 17, 62. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.A.; Lenz, M.; Gleeson, P.V. Testing assumptions of mark–recapture protocols for estimating population size using Australian mound-building, subterranean termites. Ecol. Entomol. 1998, 23, 139–159. [Google Scholar] [CrossRef]
- Evans, T.A.; Lenz, M.; Gleeson, P.V. Estimating population size and forager movement in a tropical subterranean termite (Isoptera: Rhinotermitidae). Environ. Entomol. 1999, 28, 823–830. [Google Scholar] [CrossRef]
- Noirot, C. Glands and secretion. In Biology of Termites, vol. 1; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1969; pp. 89–123. ISBN 978-0-12-4263017/978-0-12-395529-6. [Google Scholar]
- Prestwich, G.D. Chemical systematics of termite exocrine secretions. Ann. Rev. Ecol. Syst. 1983, 14, 287–311. [Google Scholar] [CrossRef]
- Costa-Leonardo, A.M.; Haifig, I. Pheromones and exocrine glands in Isoptera. Vitamins Hormones 2010, 83, 521–549. [Google Scholar] [CrossRef]
- Bordereau, C.; Pasteels, J.M. Pheromones and chemical ecology of dispersal and foraging in termites. In Biology of Termites: A Modern Synthesis; Bignell, D., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 279–320. ISBN 978-90-481-3976-7. [Google Scholar]
- Palma-Onetto, V.; Pflegerová, J.; Plarre, R.; Synek, J.; Cvačka, J.; Sillam-Dussès, D.; Šobotník, J. The labral gland in termites: Evolution and function. Biol. J. Linn. Soc. 2019, 126, 587–597. [Google Scholar] [CrossRef]
- Jirošová, A.; Sillam-Dussès, D.; Kyjaková, P.; Kalinová, B.; Dolejšová, K.; Jančařík, A.; Majer, P.; Cristaldo, P.F.; Hanus, R. Smells like home: Chemically mediated co-habitation of two termite species in a single nest. J. Chem. Ecol. 2016, 42, 1070–1081. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.A.; Inta, R.; Lai, J.C.S.; Lenz, M. Foraging vibration signals attract foragers and identify food size in the drywood termite, Cryptotermes secundus. Insectes Soc. 2007, 374–382. [Google Scholar] [CrossRef]
- Oberst, S.; Bann, G.; Lai, J.C.S.; Evans, T.A. Cryptic termites avoid predatory ants by eavesdropping on vibrational cues from their footsteps. Ecol. Lett. 2017, 20, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Ribas, C.R.; Schoereder, J.H. Are all ant mosaics caused by competition? Oecologia 2002, 131, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, X.; Arnan, X.; Retana, J. Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology? Myrmecol. News 2013, 131–147. [Google Scholar]
- Stuble, K.L.; Jurić, I.; Cerdá, X.; Sanders, N.J. Dominance hierarchies are a dominant paradigm in ant ecology (Hymenoptera: Formicidae), but should they be? And what is a dominance hierarchy anyways? Myrmecol. News 2017, 24, 71–81. [Google Scholar]
- Lenz, M.; Kard, B.; Mauldin, J.K.; Evans, T.A.; Etheridge, J.L.; Abbey, H.M. Size of food resource determines brood placement in Reticulitermes flavipes. Sociobiology 2001, 37, 361–362. [Google Scholar]
- Evans, T.A.; Gleeson, P.V. The effect of bait size, composition, presentation and inspection frequency on bait consumption in termites (Isoptera: Rhinotermitidae). Bull. Ent. Res. 2006, 96, 85–90. [Google Scholar] [CrossRef]
- Iqbal, N.; Wijedasa, L.S.; Evans, T.A. Bait station preferences in two Macrotermes species. J. Pest Sci. 2017, 90, 217–225. [Google Scholar] [CrossRef]
- Korb, J.; Schmidinger, S. Help or disperse? Cooperation in termites influenced by food conditions. Behav. Ecol. Sociobiol. 2004, 56, 89–95. [Google Scholar] [CrossRef]
- Evans, T.A.; Inta, R.; Lai, J.C.S. Foraging choice and replacement reproductives facilitate invasiveness in drywood termites. Biol. Invasions 2011, 13, 1579–1587. [Google Scholar] [CrossRef]
- Souza, T.S.; Gazal, V.S.; Fernandes, V.J.; Oliveira, A.C.C.; Aguiar-Menezes, E.L. Influence of food resource size on the foraging behavior of Nasutitermes corniger (Motschulsky). Sociobiology 2018, 65, 291–298. [Google Scholar] [CrossRef]
- Eggleton, P.; Bignell, D.E.; Sands, W.A.; Waite, B.; Wood, T.G.; Lawton, J.H. The species richness of termites (Isoptera) under differing levels of forest disturbance in the Mbalmayo Forest Reserve, southern Cameroon. J. Trop. Ecol. 1995, 11, 85–98. [Google Scholar] [CrossRef]
- Eggleton, P.; Bignell, D.E.; Hauser, S.; Dibog, L.; Norgrove, L.; Madong, B. Termite diversity across an anthropogenic disturbance gradient in the humid forest zone of West Africa. Agric. Ecosyst. Environ. 2002, 90, 189–202. [Google Scholar] [CrossRef]
- Jones, D.T.; Susilo, F.X.; Bignell, D.E.; Hardiwinoto, S.; Gillison, A.N.; Eggleton, P. Termite assemblage collapse along a land-use intensification gradient in lowland central Sumatra, Indonesia. J. Appl. Ecol. 2003, 40, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Bandeira, A.G.; Vasconcellos, A.; Silva, M.p.; Constantino, R. Effects of habitat disturbance on the termite fauna in a highland humid forest in the caatinga domain, Brazil. Sociobiology 2003, 42, 1–11. [Google Scholar]
- Carrijo, T.F.; Brandão, D.; de Oliveira, D.E.; Costa, D.A.; Santos, T. Effects of pasture implantation on the termite (Isoptera) fauna in the Central Brazilian Savanna (Cerrado). J. Insect Conserv. 2009, 13, 575–581. [Google Scholar] [CrossRef]
- Gathorne-Hardy, F.J.; Syaukani; Eggleton, P. The effects of altitude and rainfall on the composition of the termites (Isoptera) of the Leuser Ecosystem (Sumatra, Indonesia). J. Trop. Ecol. 2001, 17, 379–393. [Google Scholar] [CrossRef]
- Gathorne-Hardy, F.J.; Syaukani; Inward, D.J.G. Recovery of termite (Isoptera) assemblage structure from shifting cultivation in Barito Ulu, Kalimantan, Indonesia. J. Trop. Ecol. 2006, 22, 605–608. [Google Scholar] [CrossRef]
- Bourguignon, T.; Dahlsjö, C.A.L.; Jacquemin, J.; Gang, L.; Wijedasa, L.S.; Evans, T.A. Ant and termite communities in isolated and continuous forest fragments in Singapore. Insectes Soc. 2017, 64, 505–514. [Google Scholar] [CrossRef]
- Hausberger, B.; Korb, J. The impact of anthropogenic disturbance on assembly patterns of termite communities. Biotropica 2016, 48, 356–364. [Google Scholar] [CrossRef]
- Kalshoven, L.G.E. Observations on Coptotermes havilandi Holmgr. (javanicus Kemn.) (Isoptera). Beaufortia 1962, 101, 121–137. [Google Scholar]
- Harris, W.V. On the genus Coptotermes in Africa (Isoptera: Rhinotermitidae). Proc. R. Entomol. Soc. Lond. B 1966, 35, 161–171. [Google Scholar] [CrossRef]
- Jasmi, A.H.; Ahmad, A.H. Termite incidence on an araucaria plantation forest in Teluk Bahang, Penang. Insects 2011, 2, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.V. The role of termites in tropical forestry. Insectes Soc. 1966, 13, 255–266. [Google Scholar] [CrossRef]
- Acda, M.N. Economically important termites (Isoptera) of the Philippines their control. Sociobiology 2004, 43, 159–169. [Google Scholar]
- Lee, C.-Y.; Vongkaluang, C.; Lenz, M. Challenges to subterranean termite management of multi-genera faunas in Southeast Asia and Australia. Sociobiology 2007, 50, 213–221. [Google Scholar]
- Zhong, J.; Liu, L. Experience with Coptotermes formosanus in China (Isoptera: Rhinotermitidae). Sociobiology 2003, 41, 17–26. [Google Scholar]
- Lee, T.R.C.; Evans, T.A.; Cameron, S.L.; Hochuli, D.F.; Ho, S.Y.W.; Lo, N. Ecological diversification of the Australian Coptotermes termites and the evolution of mound building. J. Biogeogr. 2017, 44, 1405–1417. [Google Scholar] [CrossRef]
- Bourguignon, T.; Lo, N.; Šobotník, J.; Sillam-Dussès, D.; Roisin, Y.; Evans, T.A. Oceanic dispersal, vicariance and human introduction shaped the modern distribution of the termites Reticulitermes, Heterotermes and Coptotermes. Proc. R. Soc. B 2016, 283, 20160179. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, T.; Lo, N.; Šobotník, J.; Ho, S.Y.W.; Iqbal, N.; Coissac, E.; Lee, M.; Jendryka, M.M.; Sillam-Dussès, D.; Křížková, B.; et al. Mitochondrial phylogenomics resolves the global spread of higher termites, ecosystem engineers of the tropics. Mol. Biol. Evol. 2017, 34, 589–597. [Google Scholar] [CrossRef]
- Case, T.J.; Gilpin, M.E. Interference Competition and Niche Theory. Proc. Natl. Acad. Sci. USA 1974, 71, 3073–3077. [Google Scholar] [CrossRef] [Green Version]
- Schyra, J.; Korb, J. Termite communities along a disturbance gradient in a west African savanna. Insects 2019, 10, 17. [Google Scholar] [CrossRef]
- Woon, J.S.; Boyle, M.J.W.; Ewers, R.M.; Chung, A.; Eggleton, P. Termite environmental tolerances are more linked to desiccation than temperature in modified tropical forests. Insectes Sociaux 2019, 66, 57–64. [Google Scholar] [CrossRef]
- Evans, T.A.; Forschler, B.T.; Grace, J.K. Biology of invasive termites: A worldwide review. Ann. Rev. Entomol. 2013, 58, 455–474. [Google Scholar] [CrossRef]
- Evans, T.A.; Forschler, B.T.; Trettin, C.C. Not just urban: The Formosan subterranean termite, Coptotermes formosanus, is invading forests in the Southeastern USA. Biol. Invasions 2019, 21, 1283–1294. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Worker | Soldier | Number of Individuals for Equal Biomass | |
|---|---|---|---|---|
| Macrotermes barneyi | large | 10.41 ± 0.54 | 22.43 ± 0.52 | 3 |
| small | 5.45 ± 0.32 | 12.01 ± 1.67 | 2 | |
| Odontotermes formosanus | 4.99 ± 0.19 | 4.57 ± 0.13 | 8.8 ± 0.9 | |
| Coptotermes formosanus | 4.05 ± 0.06 | 4.43 ± 0.06 | 10.9 ± 1.3 | |
| Odontotermes hainanensis | 2.13 ± 0.06 | 2.11 ± 0.05 | 23.0 ± 1.0 | |
| Reticulitermes flaviceps | 2.08 ± 0.05 | 2.52 ± 0.05 | 21.1 ± 2.2 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, T.A.; Kasseney, B.D. The Dominance Hierarchy of Wood-Eating Termites from China. Insects 2019, 10, 210. https://doi.org/10.3390/insects10070210
Evans TA, Kasseney BD. The Dominance Hierarchy of Wood-Eating Termites from China. Insects. 2019; 10(7):210. https://doi.org/10.3390/insects10070210
Chicago/Turabian StyleEvans, Theodore A., and Boris Dodji Kasseney. 2019. "The Dominance Hierarchy of Wood-Eating Termites from China" Insects 10, no. 7: 210. https://doi.org/10.3390/insects10070210
APA StyleEvans, T. A., & Kasseney, B. D. (2019). The Dominance Hierarchy of Wood-Eating Termites from China. Insects, 10(7), 210. https://doi.org/10.3390/insects10070210