The Probing Behavior Component of Disease Transmission in Insect-Transmitted Bacterial Plant Pathogens


{kind=link}
{kind=link}
Abstract
:1. Introduction
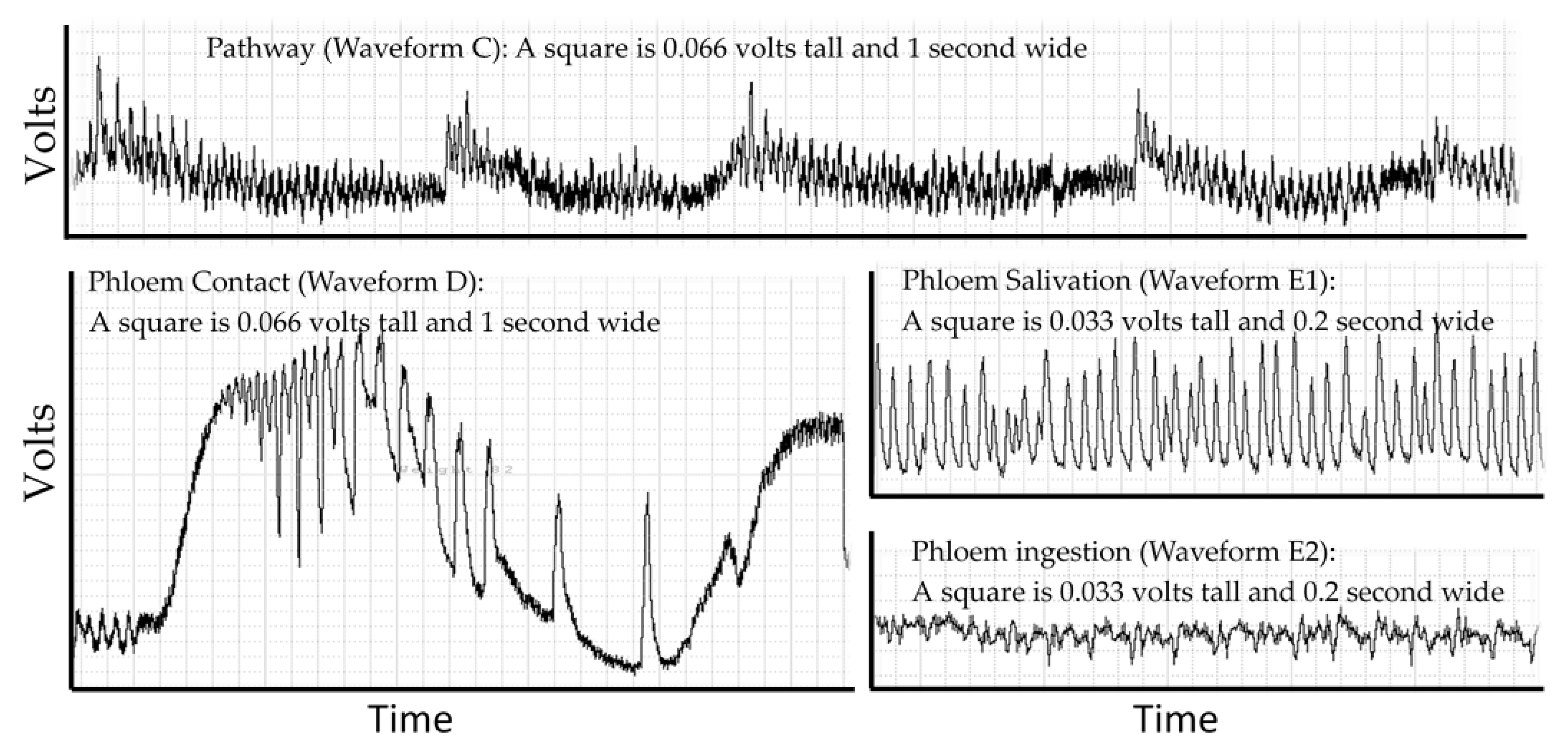
2. Electropenetrography
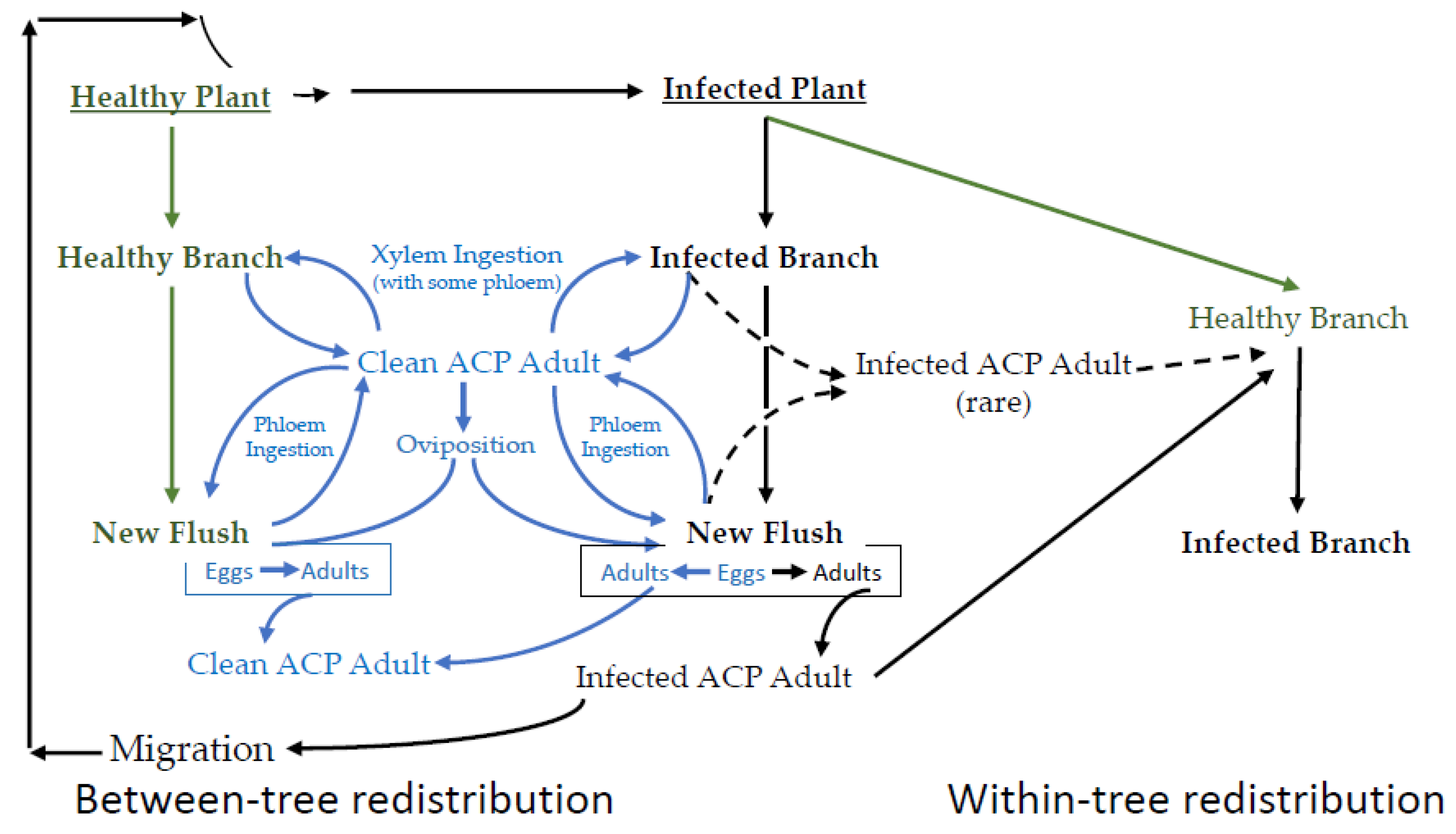
3. Candidatus Liberibacter asiaticus
3.1. CLas Adults
3.2. CLas Eggs
3.3. CLas Nymphs
3.3.1. Changes in Behavior
3.3.2. Changes in Physiology
3.4. Plant Physiology
3.5. Experimental Design Issues
3.6. EPG as a Tool
3.7. Relative Importance of Feeding Site
3.8. Psyllid Nutrition
3.9. Cell Count
4. Other Bacterial Pathosystems
4.1. Xylella fastidiosa
4.2. Candidatus Phytoplasma
5. Environmental Change
6. Conclusions
Acknowledgments
References
- Kannan, V.; Bastas, K.; Devi, R. Scientific and economic impact of plant pathogenic bacteria. In Sustainable Approaches to Controlling Plant Pathogenic Bacteria; Kannan, V., Bastas, K., Rajendran, S., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 369–392. [Google Scholar]
- Halbert, S. The discovery of Huanglongbing in Florida. In Proceedings of the 2nd International Citrus Canker and Huanglongbing Research Workshop, Orlando, FL, USA, 7–11 November 2005; p. H-3. [Google Scholar]
- Court, C.D.; Hodges, A.W.; Rahmani, M.; Spreen, T.H. Economic Contributions of the Florida Citrus Industry in 2015/16; IFAS: Gainesville, FL, USA, 2017; Available online: http://edis.ifas.ufl.edu/fe1021 (accessed on 18 July 2019).
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massonie, G.; Garnier, M.; Bove, J.M. Transmission of Indian citrus decline by Trioza erytreae (Del Guercio), the vector of South African greening. In Proceedings of the Seventh Conference of the International Organization of Citrus Virologists; IOCV: Riverside, CA, USA, 1976; pp. 18–20. Available online: https://escholarship.org/uc/item/4dh8x79m (accessed on 18 July 2019).
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Ann. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Mann, R.S.; Rogers, M.E.; Stelinski, L.L. Insecticide resistance in field populations of Asian citrus psyllid in Florida. Pest Manag. Sci. 2011, 67, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Langdon, K.W.; Ebert, T.A.; Rogers, M.E. Evaluating the effect of imidacloprid administered in artificial diet on feeding behavior of Diaphorina citri (Hemiptera: Liviidae) using electropenetrography. J. Econ. Entomol. 2019, 112, 644–652. [Google Scholar] [CrossRef]
- Serikawa, R.H.; Backus, E.; Rogers, M. Effects of soil-applied imidacloprid on Asian Citrus Psyllid (Hemiptera: Psyllidae) feeding behavior. J. Econ. Entomol. 2012, 105, 1492–1502. [Google Scholar] [CrossRef]
- Serikawa, R.H.; Backus, E.A.; Rogers, M.E. Probing behaviors of adult Asian Citrus Psyllid (Hemiptera Liviidae) are not appreciably affected by soil application of field rate aldicarb to citrus. Fla. Entomol. 2013, 96, 1334–1342. [Google Scholar] [CrossRef]
- Backus, E.A. Sharpshooter feeding behavior in relation to transmission of Xylella fastidiosa: A model for foregut-borne transmission mechanisms. In Vector-Mediated Transmission of Plant Pathogens; Brown, J.K., Ed.; American Phytopathological Society: St. Paul, MN, USA, 2016; pp. 175–193. [Google Scholar]
- Walker, G.P. A Beginner’s guide to electronic monitoring of Homopteran probing behavior. In Principles and Applications of Electronic Monitoring and Other Techniques in the Study of Homopteran Feeding Behavior; Walker, G.P., Backus, E.A., Eds.; Entomological Society of America: Annapolis, MD, USA, 2000; pp. 14–40. [Google Scholar]
- Tjallingii, W.F. Electrical nature of recorded signals during stylet penetration by aphids. Entomol. Exp. Appl. 1985, 38, 177–186. [Google Scholar] [CrossRef]
- Miles, P.W. The saliva of Hemiptera. Adv. Insect Physiol. 1972, 9, 183–255. [Google Scholar]
- Will, T.; Steckbauer, K.; Hardt, M.; van Bel, A.J. Aphid gel saliva: Sheath structure, protein composition and secretory dependence on stylet-tip milieu. PLoS ONE 2012, 7, e46903. [Google Scholar] [CrossRef]
- Morgan, J.K.; Luzio, G.A.; el Ammar, D.; Hunter, W.B.; Hall, D.G.; Shatters, R.G., Jr. Formation of stylet sheaths in aere (in air) from eight species of phytophagous hemipterans from six families (Suborders: Auchenorrhyncha and Sternorrhyncha). PLoS ONE 2013, 8, e62444. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Killiny, N. The secreted salivary proteome of Asian citrus psyllid Diaphorina citri. Physiol. Entomol. 2018, 43, 324–333. [Google Scholar] [CrossRef]
- Dugravot, S.; Backus, E.A.; Reardon, B.J.; Miller, T.A. Correlations of cibarial muscle activities of Homalodisca spp. sharpshooters (Hemiptera: Cicadellidae) with EPG ingestion waveform and excretion. J. Insect Physiol. 2008, 54, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Recatala, V.; Tjallingii, W.F. A new application of the electrical penetration graph (EPG) for acquiring and measuring electrical signals in phloem sieve elements. JOVE J. Vis. Exp. 2015, 101, e52826. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F. Comparison of AC and DC systems for electronic monitoring of stylet penetration activities by homopterans. In Principles and Applications of Electronic Monitoring and Other Techniques in the Study of Homopteran Feeding Behavior; Walker, G.P., Backus, E.A., Eds.; Entomological Society of America: Annapolis, MD, USA, 2000; pp. 41–69. [Google Scholar]
- Backus, E.A.; Cervantes, F.A.; Guedes, R.N.C.; Li, A.Y.; Wayadande, A.C. AC–DC electropenetrography for in-depth studies of feeding and oviposition behaviors. J. Entomol. Soc. Am. 2019, 112, 236–248. [Google Scholar] [CrossRef]
- Blust, M.H.; Hopkins, T.L. Feeding patterns of a specialist and a generalist grasshopper: Electronic monitoring on their host plants. Physiol. Entomol. 1990, 15, 261–267. [Google Scholar] [CrossRef]
- Guo, F.; Zhao, Z. Feeding behaviour of omethoate-resistant spider mites (Acari: Tetrnychidae): A study using electrical penetration graphs. Syst. Appl. Acarol. 2000, 5, 3–7. [Google Scholar] [CrossRef]
- Jacobson, A.L.; Kennedy, G.G. Effect of cyantraniliprole on feeding behavior and virus transmission of Frankliniella fusca and Frankliniella occidentalis (Thysanoptera: Thripidae) on Capsicum annuum. Crop Prot. 2013, 54, 251–258. [Google Scholar] [CrossRef]
- Kindt, F.; Joosten, N.N.; Tjallingii, W.F. Electrical penetration graphs of thrips revised: Combining DC- and AC-EPG signals. J. Insect Physiol. 2006, 52, 1–10. [Google Scholar] [CrossRef]
- Li, A.Y. Use of EMG, EPG, and in vitro tick feeding systems to study mechanisms of blood feeding and effects of acaricides and pharmacological agents on blood feeding in ixodid ticks. In Proceedings of the XXV International Congress of Entomology, Orlando, FL, USA, 25–30 September 2016. [Google Scholar]
- Kashin, P. Electronic recording of the mosquito bite. J. Insect Physiol. 1966, 12, 281–284. [Google Scholar] [CrossRef]
- Zimmermann, M.R.; Mithofer, A.; Will, T.; Felle, H.H.; Furch, A.C. Herbivore-triggered electrophysiological reactions: Candidates for systemic signals in higher plants and the challenge of their identification. Plant Physiol. 2016, 170, 2407–2419. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Trivedi, P. Citrus huanglongbing: A newly relevant disease presents unprecedented challenges. Phytopathology 2013, 103, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Stelinski, L.L.; Pelz-Stelinski, K.S.; Graham, J.H.; Zhang, Y. Tale of the huanglongbing disease pyramid in the context of the citrus microbiome. Phytopathology 2017, 107, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Da Graca, J.V.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G.; Zhao, H. Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Gurung, K.; Wertheim, B.; Falcao Salles, J. The microbiome of pest insects: It is not just bacteria. Entomol. Exp. Appl. 2019, 167, 156–170. [Google Scholar] [CrossRef]
- Simpson, S.J.; Raubenheimer, D.; Chambers, P.G. The mechanisms of nutritional homeostasis. In Regulatory Mechanisms in Insect Feeding; Chapman, R.F., de Boer, G., Eds.; Springer: Boston, MA, USA, 1995; pp. 251–278. [Google Scholar]
- Nouri, S.; Salem, N.; Nigg, J.C.; Falk, B.W. Diverse array of new viral sequences identified in worldwide populations of the Asian Citrus Psyllid (Diaphorina citri) using viral metagenomics. J. Virol. 2016, 90, 2434–2445. [Google Scholar] [CrossRef] [PubMed]
- Kolora, L.D.; Powell, C.M.; Hunter, W.; Bextine, B.; Lauzon, C.R. Internal extracellular bacteria of Diaphorina citri Kuwayama (Hemiptera: Psyllidae), the Asian citrus psyllid. Curr. Microbiol. 2015, 70, 710–715. [Google Scholar] [CrossRef]
- Inoue, H.; Ohnishi, J.; Ito, T.; Tomimura, K.; Miyata, S.; Iwanami, T.; Ashihara, W. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Ann. Appl. Biol. 2009, 155, 29–36. [Google Scholar] [CrossRef]
- Ammar, E.D.; Ramos, J.E.; Hall, D.G.; Dawson, W.O.; Shatters, R.G. Acquisition, replication and inoculation of Candidatus Liberibacter asiaticus following various acquisition periods on Huanglongbing-infected citrus by nymphs and adults of the Asian citrus psyllid. PLoS ONE 2016, 11, e0159594. [Google Scholar] [CrossRef]
- Pelz-Stelinski, K.S.; Brlansky, R.H.; Ebert, T.A.; Rogers, M.E. Transmission parameters for Candidatus Liberibacter asiaticus by Asian Citrus Psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 2010, 103, 1531–1541. [Google Scholar] [CrossRef]
- Mann, R.S.; Pelz-Stelinski, K.; Hermann, S.L.; Tiwari, S.; Stelinski, L.L. Sexual transmission of a plant pathogenic bacterium, Candidatus Liberibacter asiaticus, between conspecific insect vectors during mating. PLoS ONE 2011, 6, e29197. [Google Scholar] [CrossRef]
- Canale, M.C.; Tomaseto, A.F.; Haddad, M.D.; Della Coletta, H.; Lopes, J.R.S. Latency and persistence of ‘Candidatus Liberibacter asiaticus’ in its psyllid vector, Diaphorina citri (Hemiptera: Liviidae). Phytopathol. 2017, 107, 264–272. [Google Scholar] [CrossRef]
- Huang, J.H.; Luo, X.; Huang, B.C.; Yao, X. Studies on citrus psylla Diaphrina citri Kuwayama and its control. Entomol. J. East China 1999, 8, 26–34. [Google Scholar]
- Yang, Y.; Huang, M.; Beattie, G.A.C.; Xia, Y.; Ouyang, G.; Xiong, J. Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: A status report for China. Int. J. Pest Manag. 2006, 52, 343–352. [Google Scholar] [CrossRef]
- De Ibarra, M.J.A. Abundance and Population Dynamics of the Asian Citrus Psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae) as Affected by Flush Shoots in Different Host Plants. Master’s Thesis, Texas A&M University, College Station, TX, USA, 2009. [Google Scholar]
- Xu, C.F.; Xia, Y.; Li, K.B.; Ke, C. Further study of the transmission of citrus Huanglungbin by a psyllid, Diaphorina citri Kuwayama. In Proceedings of the 10th Conference of the International Organization of Citrus Virologists; IOCV: Riverside, CA, USA, 1988; pp. 243–248. [Google Scholar]
- Moir, M.L.; Renton, M.; Hoffmann, B.D.; Leng, M.C.; Lach, L. Development and testing of a standardized method to estimate honeydew production. PLoS ONE 2018, 13, e0201845. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Hall, D.G.; Shatters, R.G., Jr. Stylet morphometrics and citrus leaf vein structure in relation to feeding behavior of the asian citrus psyllid Diaphorina citri, vector of citrus Huanglongbing bacterium. PLoS ONE 2013, 8, e59914. [Google Scholar] [CrossRef]
- Hartung, J.S.; Paul, C.; Achor, D.; Brlansky, R.H. Colonization of Dodder, Cuscuta indecora, by ‘Candidatus Liberibacter asiaticus’ and ‘Ca. L. americanus’. Phytopathology 2010, 100, 756–762. [Google Scholar] [CrossRef]
- George, J.; Ammar, E.; Hall, D.G.; Shatters, R.G.; Lapointe, S.L. Prolonged phloem ingestion by Diaphorina citri nymphs compared to adults is correlated with increased acquisition of citrus greening pathogen. Sci. Rep. 2018, 8, 10352. [Google Scholar] [CrossRef]
- Sandanayaka, M.; Connolly, P.G.; Withers, T.M. Assessment of tomato potato psyllid Bactericera cockerelli (Hemiptera: Triozidae) food plant range by comparing feeding behaviour to survival of early life stages. Austral Entomol. 2017, 58, 387–394. [Google Scholar] [CrossRef]
- Zhao, R.; He, Y.; Lu, Z.; Chen, W.; Zhou, C.; Wang, X.; Li, T. An analysis of the feeding behavior of three stages of Toxoptera citricida by DC electrical penetration graph waveforms. Entomol. Exp. Appl. 2019, 167, 370–376. [Google Scholar] [CrossRef]
- Ghanim, M.; Achor, D.; Ghosh, S.; Kontsedalov, S.; Lebedev, G.; Levy, A. ‘Candidatus Liberibacter asiaticus’ accumulates inside endoplasmic reticulum associated vacuoles in the gut cells of Diaphorina citri. Sci. Rep. 2017, 7, 16945. [Google Scholar] [CrossRef]
- Ghanim, M.; Fattah-Hosseini, S.; Levy, A.; Cilia, M. Morphological abnormalities and cell death in the Asian citrus psyllid (Diaphorina citri) midgut associated with Candidatus Liberibacter asiaticus. Sci. Rep. 2016, 6, 33418. [Google Scholar] [CrossRef]
- Mann, M.; Fattah-Hosseini, S.; Ammar, E.; Stange, R.; Warrick, E.; Sturgeon, K.; Shatters, R.; Heck, M. Diaphorina citri nymphs are resistant to morphological changes induced by “Candidatus Liberibacter asiaticus” in midgut epithelial cells. Infect. Immun. 2018, 86, e00889-17. [Google Scholar] [CrossRef]
- Ramsey, J.S.; Chavez, J.D.; Johnson, R.; Hosseinzadeh, S.; Mahoney, J.E.; Mohr, J.P.; Robison, F.; Zhong, X.; Hall, D.G.; MacCoss, M.; et al. Protein interaction networks at the host-microbe interface in Diaphorina citri, the insect vector of the citrus greening pathogen. R. Soc. Open Sci. 2017, 4, 160545. [Google Scholar] [CrossRef]
- Kruse, A.; Fattah-Hosseini, S.; Saha, S.; Johnson, R.; Warwick, E.; Sturgeon, K.; Mueller, L.; MacCoss, M.J.; Shatters, R.G.; Heck, M.C. Combining’omics and microscopy to visualize interactions between the Asian citrus psyllid vector and the Huanglongbing pathogen Candidatus Liberibacter asiaticus in the insect gut. PLoS ONE 2017, 12, e0179531. [Google Scholar] [CrossRef]
- Gill, T.A.; Chu, C.; Pelz-Stelinski, K.S. Comparative proteomic analysis of hemolymph from uninfected and Candidatus Liberibacter asiaticus-infected Diaphorina citri. Amino Acids 2017, 49, 389–406. [Google Scholar] [CrossRef]
- Jain, M.; Fleites, L.A.; Gabriel, D.W. A small wolbachia protein directly represses phage lytic cycle genes in “Candidatus Liberibacter asiaticus” within Psyllids. Msphere 2017, 2, e00171-17. [Google Scholar] [CrossRef]
- Ammar, E.; Hall, D.G.; Hosseinzadeh, S.; Heck, M. The quest for a non-vector psyllid: Natural variation in acquisition and transmission of the huanglongbing pathogen ‘Candidatus Liberibacter asiaticus’ by Asian citrus psyllid isofemale lines. PLoS ONE 2018, 13, e0195804. [Google Scholar] [CrossRef]
- Killiny, N.; Hijaz, F.; Ebert, T.A.; Rogers, M.E. A plant bacterial pathogen manipulates its insect vector’s energy metabolism. Appl. Environ. Microbiol. 2017, 83, e03005-16. [Google Scholar] [CrossRef]
- Wu, T.Y.; Luo, X.Z.; Xu, C.B.; Wu, F.N.; Qureshi, J.A.; Cen, Y.J. Feeding behavior of Diaphorina citri and its transmission of ‘Candidatus Liberibacter asiaticus’ to citrus. Entomol. Exp. Appl. 2016, 161, 104–111. [Google Scholar] [CrossRef]
- Chu, C.C.; Gill, T.A.; Hoffmann, M.; Pelz-Stelinski, K.S. Inter-population variability of endosymbiont densities in the Asian Citrus Psyllid (Diaphorina citri Kuwayama). Microb. Ecol. 2016, 71, 999–1007. [Google Scholar] [CrossRef]
- Fagen, J.R.; Giongo, A.; Brown, C.T.; Davis-Richardson, A.G.; Gano, K.A.; Triplett, E.W. Characterization of the relative abundance of the Citrus Pathogen Ca. Liberibacter Asiaticus in the microbiome of its insect vector, Diaphorina citri, using high throughput 16S rRNA sequencing. Open Microbiol. J. 2012, 6, 29–33. [Google Scholar] [CrossRef]
- Dan, H.; Ikeda, N.; Fujikami, M.; Nakabachi, A. Behavior of bacteriome symbionts during transovarial transmission and development of the Asian citrus psyllid. PLoS ONE 2017, 12, e0189779. [Google Scholar] [CrossRef]
- Ren, S.L.; Li, Y.H.; Ou, D.; Guo, Y.J.; Qureshi, J.A.; Stansly, P.A.; Qiu, B.L. Localization and dynamics of Wolbachia infection in Asian citrus psyllid Diaphorina citri, the insect vector of the causal pathogens of Huanglongbing. MicrobiologyOpen 2018, 7, e00561. [Google Scholar] [CrossRef]
- Dossi, F.C.A.; da Silva, E.P.; Consoli, F.L. Population dynamics and growth rates of endosymbionts during Diaphorina citri (Hemiptera, Liviidae) ontogeny. Microb. Ecol. 2014, 68, 881–889. [Google Scholar] [CrossRef]
- Nakabachi, A.; Yamashita, A.; Toh, H.; Ishikawa, H.; Dunbar, H.E.; Moran, N.A.; Hattori, M. The 160-kilobase genome of the bacterial endosymbiont Carsonella. Science 2006, 314, 267. [Google Scholar] [CrossRef]
- Nakabachi, A.; Ueoka, R.; Oshima, K.; Teta, R.; Mangoni, A.; Gurgui, M.; Oldham, N.J.; van Echten-Deckert, G.; Okamura, K.; Yamamoto, K.; et al. Defensive bacteriome symbiont with a drastically reduced genome. Curr. Biol. 2013, 23, 1478–1484. [Google Scholar] [CrossRef]
- Ponder, K.L.; Pritchard, J.; Harrington, R.; Bale, J.S. Feeding behaviour of the aphid Rhopalosiphum padi (Hemiptera: Aphididae) on nitrogen and water-stressed barley (Hordeum vulgare) seedlings. Bull. Entomol. Res. 2001, 91, 125–130. [Google Scholar]
- Zavala, J.A.; Nabity, P.D.; DeLucia, E.H. An emerging understanding of mechanisms governing insect herbivory under elevated CO2. Ann. Rev. Entomol. 2013, 58, 79–97. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Futch, D.B.; Shilts, T.; Folimonova, S.Y.; Reyes-De-Corcuera, J.I. GC-MS metabolomic differentiation of selected citrus varieties with different sensitivity to citrus huanglongbing. Plant Physiol. Biochem. 2012, 53, 69–76. [Google Scholar] [CrossRef]
- Hung, W.L.; Wang, Y. A targeted mass spectrometry-based metabolomics approach toward the understanding of host responses to Huanglongbing disease. J. Agric. Food Chem. 2018, 66, 10651–10661. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y. Metabolomic response to Huanglongbing: Role of carboxylic compounds in Citrus sinensis response to Candidatus Liberibacter asiaticus and its vector, Diaphorina citri. Mol. Plant Microbe Interact. 2017, 30, 666–678. [Google Scholar] [CrossRef]
- Cen, Y.; Yang, C.; Holford, P.; Beattie, G.A.C.; Spooner-Hart, R.N.; Liang, G.; Deng, X. Feeding behaviour of the Asiatic citrus psyllid, Diaphorina citri, on healthy and huanglongbing-infected citrus. Entomol. Exp. Appl. 2012, 143, 13–22. [Google Scholar] [CrossRef]
- Shugart, H. Probing Behavior and Host Preference in the Asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae). Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2019. [Google Scholar]
- Setamou, M.; Alabi, O.J.; Simpson, C.R.; Jifon, J.L. Contrasting amino acid profiles among permissive and non-permissive hosts of Candidatus Liberibacter asiaticus, putative causal agent of Huanglongbing. PLoS ONE 2017, 12, e0187921. [Google Scholar] [CrossRef]
- Louzada, E.S.; Vazquez, O.E.; Braswell, W.E.; Yanev, G.; Devanaboina, M.; Kunta, M. Distribution of ‘Candidatus Liberibacter asiaticus’ above and below ground in Texas citrus. Phytopathology 2016, 106, 702–709. [Google Scholar] [CrossRef]
- Etxeberria, E.; Gonzalez, P.; Vincent, C.; Schumann, A. Extended persistence Candidatus Liberibacter asiaticus (CLas) DNA in Huanglongbing-affected citrus tissue after bacterial death. Physiol. Mol. Plant Pathol. 2019, 106, 204–207. [Google Scholar] [CrossRef]
- Bonani, J.P.; Fereres, A.; Garzo, E.; Miranda, M.P.; Appezzato-Da-Gloria, B.; Lopes, J.R.S. Characterization of electrical penetration graphs of the Asian citrus psyllid, Diaphorina citri, in sweet orange seedlings. Entomol. Exp. Appl. 2010, 134, 35–49. [Google Scholar] [CrossRef]
- Youn, Y.; Backus, E.A.; Serikawa, R.H.; Stelinski, L.L. Correlation of an electrical penetration graph waveform with walking by Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Fla. Entomol. 2011, 94, 1084–1087. [Google Scholar] [CrossRef]
- Kim, K.D. Integrated Management of Asian Citrus Psyllid, Diaphorina citri Kuwayama, for Protecting Young Citrus Trees from Huanglongbing; University of Florida: Gainesville, FL, USA, 2013. [Google Scholar]
- Miranda, M.P.; Zanardi, O.Z.; Tomaseto, A.F.; Volpe, H.X.L.; Garcia, R.B.; Prado, E. Processed kaolin affects the probing and settling behavior of Diaphorina citri (Hemiptera: Lividae). Pest Manag. Sci. 2018, 74, 1964–1972. [Google Scholar] [CrossRef]
- Ebert, T.A.; Rogers, M.E. Effect of substrate voltage on EPG recordings of ingestion and probing behavior in Diaphorina citri (Hemiptera: Liviidae). Fla. Entomol. 2016, 99, 528–534. [Google Scholar] [CrossRef]
- Ebert, T.A.; Backus, E.A.; Shugart, H.J.; Rogers, M.E. Behavioral plasticity in probing by Diaphorina citri (Hemiptera, Liviidae): Ingestion from phloem versus xylem is influenced by leaf age and surface. J. Insect Behav. 2018, 31, 119–137. [Google Scholar] [CrossRef]
- Hall, D.G.; Albrecht, U.; Bowman, K.D. Transmission rates of ‘Ca. Liberibacter asiaticus’ by Asian citrus psyllid are enhanced by the presence and developmental stage of citrus flush. J. Econ. Entomol. 2016, 109, 558–563. [Google Scholar] [CrossRef]
- Setamou, M.; Alabi, O.J.; Kunta, M.; Jifon, J.L.; da Graca, J.V. Enhanced acquisition rates of ‘Candidatus Liberibacter asiaticus’ by the Asian Citrus Psyllid (Hemiptera: Liviidae) in the presence of vegetative flush growth in citrus. J. Econ. Entomol. 2016, 109, 1973–1978. [Google Scholar] [CrossRef]
- Almeida, R.P.P. Xylella fastidiosa vector transmission biology. In Vector-Mediated Transmission of Plant Pathogens; Brown, J.K., Ed.; American Phytopathological Society: St. Paul, MN, USA, 2016; pp. 165–173. [Google Scholar]
- Lopes, J.R.S.; Krugner, R. Transmission ecology and epidemioogy of the citrus variegated chlorosis virus strain of Xylella fastidiosa. In Vector-Mediated Transmission of Plant Pathogens; Brown, J.K., Ed.; American Phytopathological Society: St. Paul, MN, USA, 2016; pp. 195–208. [Google Scholar]
- Krugner, R.; Sisterson, M.S.; Backus, E.A.; Burbank, L.P.; Redak, R.A. Sharpshooters: A review of what moves Xylella fastidiosa. Austral Entomol. 2019, 58, 248–267. [Google Scholar] [CrossRef]
- Rapicavoli, J.; Ingel, B.; Blanco-Ulate, B.; Cantu, D.; Roper, C. Xylella fastidiosa: An examination of a re-emerging plant pathogen. Mol. Plant Pathol. 2018, 19, 786–800. [Google Scholar] [CrossRef]
- Backus, E.A.; Morgan, D.J. Spatiotemporal colonization of Xylella fastidiosa in its vector supports the role of egestion in the inoculation mechanism of foregut-borne plant pathogens. Phytopathology 2011, 101, 912–922. [Google Scholar] [CrossRef]
- Backus, E.A.; Shugart, H.J.; Rogers, E.E.; Morgan, J.K.; Shatters, R. Direct evidence of egestion and salivation of Xylella fastidiosa suggests sharpshooters can be “flying syringes”. Phytopathology 2015, 105, 608–620. [Google Scholar] [CrossRef]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Ann. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef]
- Bosco, D.; Marzachi, C. Insect trnsmission of phytoplasmas. In Vector-Mediated Transmission of Plant Pathogens; Brown, J.K., Ed.; American Phytopathological Society: St. Paul, MN, USA, 2016; pp. 319–327. [Google Scholar]
- Beanland, L.; Hoy, C.W.; Miller, S.A.; Nault, L.R. Leafhopper (Homoptera: Cicadellidae) transmission of aster yellows phytoplasma: Does gender matter? Environ. Entomol. 1999, 28, 1101–1106. [Google Scholar] [CrossRef]
- Chuche, J.; Thiery, D. Can differences in feeding behaviour between Scaphoideus titanus males and females be related to phytoplasma transmission effciency? Integr. Prot. Prod. Vitic. IOBC WPRS Bull. 2014, 105, 177–183. [Google Scholar]
- Chuche, J.; Sauvion, N.; Thiery, D. Mixed xylem and phloem sap ingestion in sheath-feeders as normal dietary behavior: Evidence from the leafhopper Scaphoideus titanus. J. Insect Physiol. 2017, 102, 62–72. [Google Scholar] [CrossRef]
- Aucott, M. Are changes in plants due to enhanced CO2 contributing to insect population declines? Environ. Entomol. 2019, 48, 274–275. [Google Scholar] [CrossRef]
- Kremer, J.M.M.; Nooten, S.S.; Cook, J.M.; Ryalls, J.M.W.; Barton, C.V.M.; Johnson, S.N. Elevated atmospheric carbon dioxide concentrations promote ant tending of aphids. J. Anim. Ecol. 2018, 87, 1475–1483. [Google Scholar] [CrossRef]
- Dossi, F.C.A.; da Silva, E.P.; Consoli, F.L. Shifting the balance: Heat stress challenges the symbiotic interactions of the Asian Citrus Psyllid, Diaphorina citri (Hemiptera, Liviidae). Biol. Bull. 2018, 235, 195–203. [Google Scholar] [CrossRef]
- Hussain, M.; Akutse, K.S.; Ravindran, K.; Lin, Y.W.; Bamisile, B.S.; Qasim, M.; Dash, C.K.; Wang, L.D. Effects of different temperature regimes on survival of Diaphorina citri and its endosymbiotic bacterial communities. Environ. Microbiol. 2017, 19, 3439–3449. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Tada, A.; Musolin, D.L.; Hari, N.; Hosokawa, T.; Fujisaki, K.; Fukatsu, T. Collapse of insect gut symbiosis under simulated climate change. MBio 2016, 7, e01578-16. [Google Scholar] [CrossRef]
- El-Shesheny, I.; Hijaz, F.; El-Hawary, I.; Mesbah, I.; Killiny, N. Impact of different temperatures on survival and energy metabolism in the Asian citrus psyllid, Diaphorina citri Kuwayama. Comp. Biochem. Physiol. A 2016, 192, 28–37. [Google Scholar] [CrossRef]
- Dader, B.; Fereres, A.; Moreno, A.; Trebicki, P. Elevated CO2 impacts bell pepper growth with consequences to Myzus persicae life history, feeding behaviour and virus transmission ability. Sci. Rep. 2016, 6, 19120. [Google Scholar] [CrossRef]


© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebert, T.A. The Probing Behavior Component of Disease Transmission in Insect-Transmitted Bacterial Plant Pathogens. Insects 2019, 10, 212. https://doi.org/10.3390/insects10070212
Ebert TA. The Probing Behavior Component of Disease Transmission in Insect-Transmitted Bacterial Plant Pathogens. Insects. 2019; 10(7):212. https://doi.org/10.3390/insects10070212
Chicago/Turabian StyleEbert, Timothy A. 2019. "The Probing Behavior Component of Disease Transmission in Insect-Transmitted Bacterial Plant Pathogens" Insects 10, no. 7: 212. https://doi.org/10.3390/insects10070212
APA StyleEbert, T. A. (2019). The Probing Behavior Component of Disease Transmission in Insect-Transmitted Bacterial Plant Pathogens. Insects, 10(7), 212. https://doi.org/10.3390/insects10070212