Impact of Exclusion Netting Row Covers on ‘Honeycrisp’ Apple Trees Grown under Northeastern North American Conditions: Effects on Photosynthesis and Fruit Quality

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Exclusion System
2.2. Photosynthetic Activity
2.3. Fruit Quality
2.4. Fruit Cuticle Integrity
2.5. Fruit Load and Yield
2.6. Analyses
3. Results
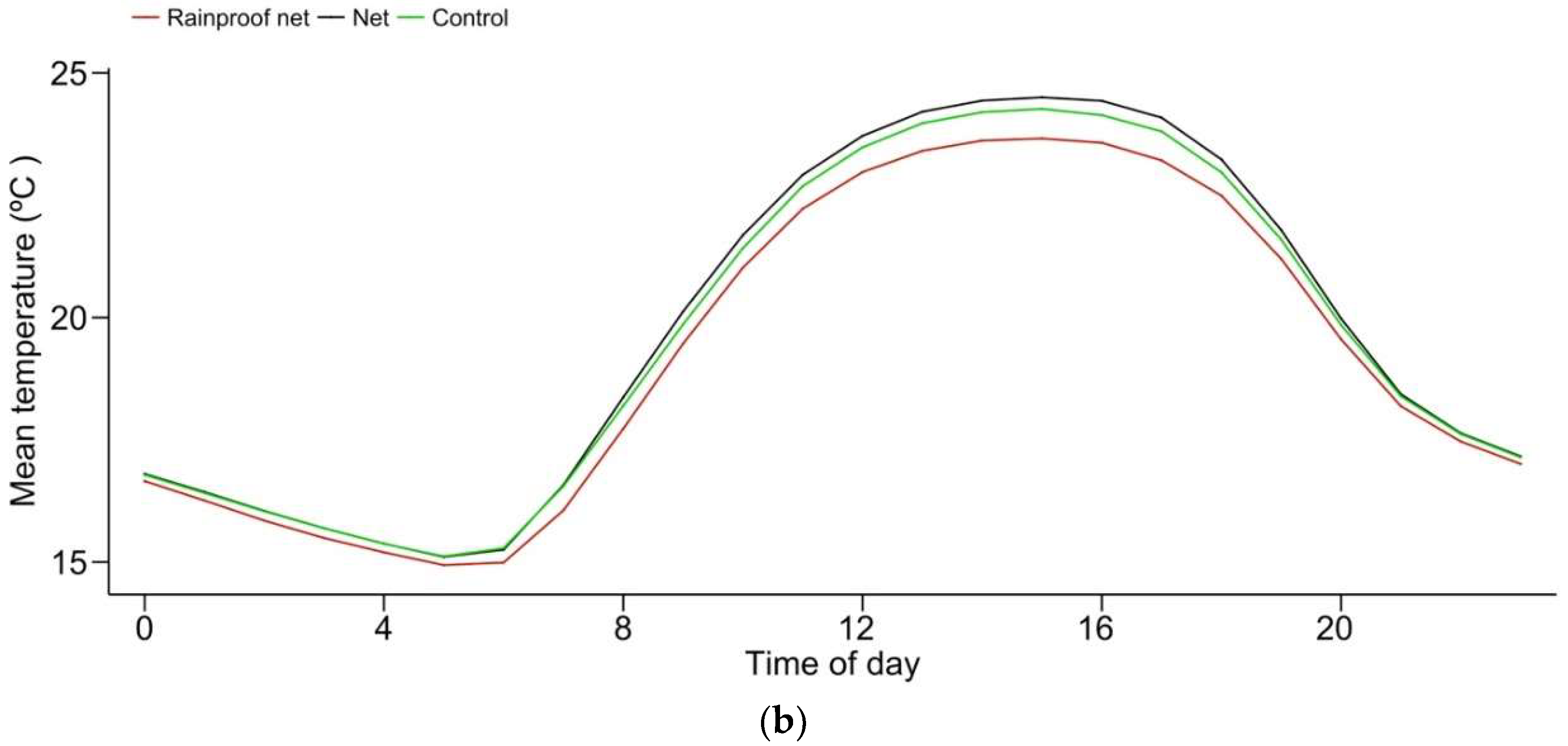
3.1. Abiotic Conditions
3.2. Photosynthetic Activity
3.3. Fruit Quality
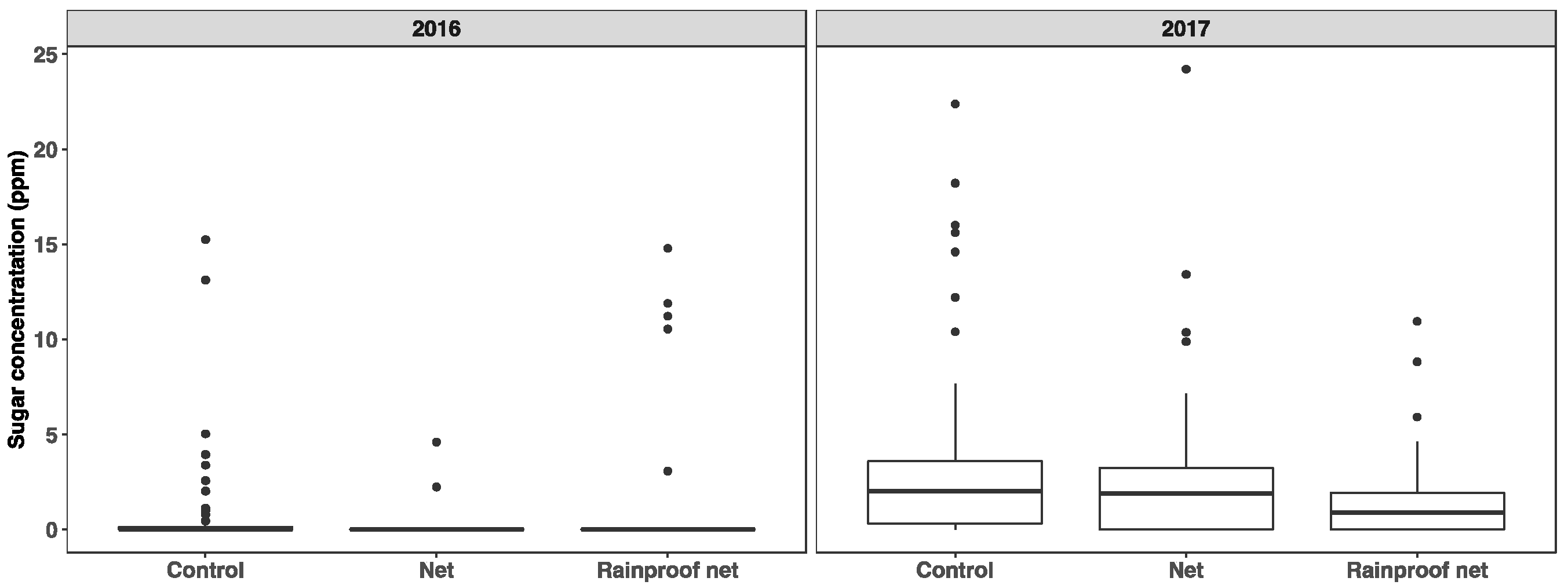
3.4. Fruit Cuticle Integrity
3.5. Fruit Load and Yield
4. Discussion
4.1. Abiotic Conditions
4.2. Photosynthetic Activity
4.3. Fruit Quality
4.4. Fruit Cuticle Integrity
4.5. Fruit Load and Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chouinard, G.; Veilleux, J.; Pelletier, F.; Larose, M.; Philion, V.; Cormier, D. Impact of exclusion netting row covers on arthropod presence and crop damage to ‘Honeycrisp’ apple trees in North America: A five-year study. Crop Prot. 2017, 98, 248–254. [Google Scholar] [CrossRef]
- Chouinard, G.; Firlej, A.; Cormier, D. Going beyond sprays and killing agents: Exclusion, sterilization and disruption for insect pest control in pome and stone fruit orchards. Sci. Hortic. 2016, 208, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Dorigoni, A. Innovative fruit tree architecture as a nexus to improve sustainability in orchards. Acta Hortic. 2016, 1137, 1–10. [Google Scholar] [CrossRef]
- Grasswitz, T.R. Integrated Pest Management (IPM) for Small-Scale Farms in Developed Economies: Challenges and Opportunities. Insects 2019, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Mupambi, G.; Anthony, B.M.; Layne, D.R.; Musacchi, S.; Serra, S.; Schmidt, T.; Kalcsits, L.A. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. 2018, 236, 60–72. [Google Scholar] [CrossRef]
- Vincent, R.; Roy, M. Entomological limits to biological control programs in Quebec apple orchards. Acta Phytopathol. Entomol. Hung. 1992, 27, 649–657. [Google Scholar]
- Candian, V.; Pansa, M.G.; Briano, R.; Peano, C.; Tedeschi, R.; Tavela, L. Exclusion nets: A promising tool to prevent Halyomorpha Halys from damaging nectarines and apples in NW Italy. Bull. Insectol. 2018, 71, 21–30. [Google Scholar]
- Stampar, F.; Veberic, R.; Zadravec, P.; Hudina, M.; Usenik, V.; Solar, A.; Osterc, G. Yield and fruit quality of apples cv. ‘Jonagold’ under hail protection nets/Ertrag und fruchtqualität der apfelsorte ‘Jonagold’ unter hagelschutznetzen. Die Gartenbauwissenschaft 2002, 67, 205–210. [Google Scholar]
- Kjaer, K.H.; Petersen, K.K.; Bertelsen, M. Protective rain shields alter leaf microclimate and photosynthesis in organic apple production. Acta Hortic. 2016, 1134, 317–326. [Google Scholar] [CrossRef]
- Martin, L.B.B.; Rose, J.K.C. There’s more than one way to skin a fruit: Formation and functions of fruit cuticles. J. Exp. Bot. 2014, 65, 4639–4651. [Google Scholar] [CrossRef]
- Børve, J.; Sekse, L.; Stensvand, A. Cuticular fractures promote postharvest fruit rot in sweet cherries. Plant Dis. 2000, 84, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Peschel, S.; Knoche, M. Characterization of microcracks in the cuticle of developing sweet cherry fruit. J. Am. Soc. Hortic. Sci. 2005, 130, 487–495. [Google Scholar] [CrossRef]
- Knoche, M.; Grimm, E. Surface Moisture Induces Microcracks in the Cuticle of ‘Golden Delicious’ Apple. HortScience 2008, 43, 1929–1931. [Google Scholar] [CrossRef]
- Agnello, A.; Chouinard, G.; Firlej, A.; Turechek, W.; Vanoosthuyse, F.; Vincent, C. Tree Fruit Field Guide to Insect, Mite and Disease Pests and Natural Enemies of Eastern North America; Natural Resource, Agriculture, and Engineering Service: Ithaca, NY, USA, 2006. [Google Scholar]
- Gitelson, A.A.; Buschmann, C.; Lichtenthaler, H.K. Chlorophyll Fluorescence Ratio F735/F700 as an accurate measure of the chlorophyll content in plants. Remote Sens. Environ. 1999, 69, 296–302. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Gitelson, A.A.; Schepers, J.S.; Walthall, C.L. Application of Spectral Remote Sensing for Agronomic Decisions. Agron. J. 2008, 100 (Suppl. S3), 117–131. [Google Scholar] [CrossRef]
- Producteurs de pommes du Québec. Charte Honeycrisp. Available online: http://producteursdepommesduquebec.ca/wp-content/uploads/2013/12/charte-honeycrisp_final2012.pdf (accessed on 13 May 2019).
- Nickerson, D. Horticultural colour chart names with Munsell key. J. Opt. Soc. Am. 1957, 47, 619–621. [Google Scholar] [CrossRef]
- Nicolaï, B.M.; Defraeye, T.; De Ketelaere, B.; Herremans, E.; Hertog, M.L.A.T.M.; Saeys, W.; Torricelli, A.; Vandendriessche, T.; Verboven, P. Nondestructive Measurement of Fruit and Vegetable Quality. Annu. Rev. Food Sci. Technol. 2014, 5, 285–312. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Sun, D.W. Colour measurements by computer vision for food quality control—A review. Trends Food Sci. Technol. 2013, 29, 5–20. [Google Scholar] [CrossRef]
- Musacchi, S.; Serra, S. Apple Fruit Quality: Overview on Pre-Harvest Factors. Sci. Hortic. 2018, 234, 409–430. [Google Scholar] [CrossRef]
- Blanpied, G.D.; Silsby, K. Predicting Harvest Date Windows for Apples; Cornell Cooperative Extension: Ithaca, NY, USA, 1992; pp. 11–12. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Nakawaga, S.; Schielzeth, H.A. General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Horthorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J. Stat. Soft. 2010, 34. Available online: https://www.jstatsoft.org/index.php/jss/article/view/v034i02/v34i02.pdf (accessed on 12 June 2019). [CrossRef]
- Zeileis, A.; Hothorn, T. Diagnostic Checking in Regression Relationships. R News 2002, 2, 7–10. Available online: https://cran.r-project.org/doc/Rnews/Rnews_2002-3.pdf (accessed on 12 June 2019).
- Ogle, D.H.; Wheeler, P.; Dinno, A. Simple Fisheries Stock Assessment Methods. R package version 0.8.24. 2019. Available online: https://cran.r-project.org/web/packages/FSA/FSA.pdf (accessed on 12 June 2019).
- Christensen, R.H.B. Regression Models for Ordinal Data, 2015 ed.; R Foundation for Statistical Computing: Vienna, Austria, 2015; pp. 6–28. [Google Scholar]
- Mahmood, A.; Hu, Y.; Tanny, J.; Asante, E.A. Effects of shading and insect-proof screens on crop microclimate and production: A review of recent advances. Sci. Hortic. 2018, 241, 241–251. [Google Scholar] [CrossRef]
- Kalcsits, L.; Musacchi, S.; Layne, D.R.; Schmidt, T.; Mupambi, G.; Serra, S.; Mendoza, M.; Asteggiano, L.; Jarolmasjed, S.; Sankaran, S.; et al. Above and below-ground environmental changes associated with the use of photoselective protective netting to reduce sunburn in apple. Agric. For. Meteorol. 2017, 237, 9–17. [Google Scholar] [CrossRef]
- Sévérac, G.; Siegwart, M. Protection Alt’Carpo, nouvelles études sur trois ans. Phytoma 2013, 668, 33–37. [Google Scholar]
- Lutte aux ennemis du pommier à l’aide de filets d’exclusion pour la production de pommes à haute valeur économique: Quatre années d’études multidisciplinaires. Available online: https://www.mapaq.gouv.qc.ca/SiteCollectionDocuments/Recherche_Innovation/Arbresfruitiers/IA113067.pdf (accessed on 6 June 2019).
- Blanke, M. The structure of coloured hail nets affects light transmission, light spectrum, phytochrome and apple fruit colouration. Acta Hortic. 2009, 817, 177–184. [Google Scholar] [CrossRef]
- Basile, B.; Giaccone, M.; Cirillo, C.; Ritieni, A.; Graziani, G.; Shahak, Y.; Forlani, M. Photo-selective hail nets affect fruit size and quality in Hayward kiwifruit. Sci. Hortic. 2012, 141, 91–97. [Google Scholar] [CrossRef]
- Rajapakse, N.C.; Shahak, Y. Light quality manipulation by horticulture industry. In Annual Plant Reviews, Light and Plant Development; Whitelam, G., Halliday, K., Eds.; Blackwell Publishing: Oxford, UK, 2007; Volume 30, pp. 290–312. [Google Scholar]
- Shahak, Y.; Gussakovsky, E.E.; Cohen, Y.; Lurie, S.; Stern, R.; Kfir, S.; Naor, A.; Atz- mon, I.; Doron, I.; Greenblat-Avron, Y. ColorNets: A new approach for light manipulation in fruit trees. Acta Hortic. 2004, 636, 609–616. [Google Scholar] [CrossRef]
- Barriault, E.; (Ministère de L’agriculture, des Pêcheries et de L’alimentation, Saint-JEAN-Sur-Richelieu, QC, Canada). Personal communication, 2019.
- Mupambi, G.; Musacchi, S.; Serra, S.; Kalcsits, L.A.; Layne, D.R.; Schmidt, T. Protective Netting Improves Leaf-level Photosynthetic Light Use Efficiency in ‘Honeycrisp’ Apple Under Heat Stress. HortScience 2018, 53, 1416–1422. [Google Scholar] [CrossRef]
- Gouws, A.; Steyn, W.J. The effect of temperature, region and season on red colour development in apple peel under constant irradiance. Sci. Hortic. 2014, 173, 79–85. [Google Scholar] [CrossRef]
- Solomakhin, A.; Blanke, M.M. Can coloured hailnets improve taste (sugar, sugar: Acid ratio), consumer appeal (colouration) and nutritional value (anthocyanin, vitamin C) of apple fruit? LWT Food Sci. Technol. 2010, 43, 1277–1284. [Google Scholar] [CrossRef]
- Do Amarante, C.V.T.; Steffens, C.A.; Argenta, L.C. Yield and fruit quality of ‘Gala’ and ‘Fuji’ apple trees protected by white anti-hail net. Sci. Hortic. 2011, 129, 79–85. [Google Scholar] [CrossRef]
- Gindaba, J.; Wand, S.J.E. Comparative Effects of Evaporative Cooling, Kaolin Particle Film, and Shade Net on Sunburn and Fruit Quality in Apples. HortScience 2005, 40, 592–596. [Google Scholar] [CrossRef]
- Mukherjee, A.; Knoch, S.; Chouinard, G.; Tavares, J.R.; Dumont, M.-J. Use of bio-based polymers in agricultural exclusion nets: A perspective. Biosyst. Eng. 2019, 180, 121–145. [Google Scholar] [CrossRef]
- Manja, K.; Aoun, M. The use of nets for tree fruit crops and their impact on the production: A review. Sci. Hortic. 2019, 246, 110–122. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Value or Interval |
|---|---|
| Shutter Speed | 1/15 |
| Sensitivity | 4.0 |
| Shift | 10 |
| Span | 1.0 |
| White Balance Coefficient 1 | R 0.931; G 1.000; B 1.268 |
| Hue | 15–200 |
| Saturation | 0–255 |
| Brightness | 40–255 |
| Year (n) | Control | Net | Rainproof Net |
|---|---|---|---|
| Chlorophyll concentration (mg/m2): | |||
| 2012 (36; 36) | 682.42 ± 23.26 a | 628.33 ± 23.71 b | |
| 2013 (36; 36) | 671.28 ± 21.23 | 711.22 ± 23.07 | |
| 2015 (72; 72; 66) | 795.85 ± 15.46 | 810.79 ± 15.69 | 822.83 ± 18.21 |
| 2016 (48; 48; 48) | 667.58 ± 18.90 | 684.77 ± 20.37 | 719.54 ± 18.68 |
| 2017 (24; 24; 24) | 752.17 ± 28.24 | 740.50 ± 30.56 | 791.54 ± 21.71 |
| Fv/Fm ratio: | |||
| 2012 (36; 36) | 0.78 ± 0.01 | 0.77 ± 0.02 | |
| 2013 (36; 36) | 0.75 ± 0.06 a | 0.76 ± 0.09 b | |
| 2015 (72; 72; 66) | 0.78 ± 0.01 | 0.78 ± 0.00 | 0.79 ± 0.01 |
| 2016 (48; 48; 48) | 0.78 ± 0.01 ab | 0.76 ± 0.01 a | 0.80 ± 0.00 b |
| 2017 (24; 24; 24) | 0.74 ± 0.02 | 0.71 ± 0.02 | 0.77 ± 0.01 |
| Year (n) | Control | Net | Rainproof Net |
|---|---|---|---|
| Red colour index (2013–2014) or % red surface: | |||
| 2013 (120; 120) | 2.26 ± 0.07 a | 2.06 ± 0.07 b | |
| 2014 (80; 73) | 1.75 ± 0.08 | 1.64 ± 0.09 | |
| 2015 (120; 120; 120) | 47.74 ± 2.13 | 46.60 ± 2.60 | 43.11 ± 1.92 |
| 2016 (103; 120; 120) | 37.89 ± 2.63 | 41.86 ± 2.18 | 32.05 ± 2.00 |
| 2017 (120; 120; 120) | 47.95 ± 2.18 | 50.01 ± 2.16 | 32.92 ± 4.15 |
| Soluble solids concentration (°Brix): | |||
| 2013 (60; 60) | 12.79 ± 0.14 | 12.14 ± 0.10 | |
| 2014 (79; 73) | 12.01 ± 0.08 | 11.93 ± 0.08 | |
| 2015 (60; 60; 60) | 11.84 ± 0.15 | 11.94 ± 0.16 | 11.91 ± 0.14 |
| 2016 (60; 60; 61) | 12.36 ± 0.18 | 12.31 ± 0.12 | 12.31 ± 0.13 |
| 2017 (60; 60; 60) | 11.72 ± 0.13 | 12.00 ± 0.14 | 12.00 ± 0.09 |
| Number of seeds (per fruit): | |||
| 2013 (120; 120) | 3.78 ± 0.21 | 3.53 ± 0.19 | |
| 2014 (79; 73) | 5.82 ± 0.23 a | 4.89 ± 0.29 b | |
| 2015 (120; 120; 120) | 5.40 ± 0.21 a | 4.35 ± 0.21 ab | 3.83 ± 0.20 b |
| 2016 (103; 120; 119) | 6.94 ± 0.22 | 6.14 ± 0.21 | 6.07 ± 0.22 |
| 2017 (120; 120; 120) | 7.18 ± 0.21 a | 5.93 ± 0.23 b | 4.91 ± 0.22 c |
| Firmness (kg): | |||
| 2013 (60;60) | 6,24 ± 0.07 | 6.20 ± 0.08 | |
| 2014 (79; 73) | 8.01 ± 0.06 | 8.32 ± 0.07 | |
| 2015 (120; 120; 120) | 6.37 ± 0.06 | 6.28 ± 0.06 | 6.31 ± 0.06 |
| 2016 (60; 60; 61) | 7.05 ± 0.10 | 6.75 ± 0.06 | 6.57 ± 0.06 |
| 2017 (60; 60; 60) | 7.07 ± 0.09 | 7.23 ± 0.08 | 7.36 ± 0.08 |
| Maturity (% starch staining): | |||
| 2013 (60; 60) | 20.57 ± 0.99 a | 24.00 ± 0.99 b | |
| 2014 (79; 73) | 48.43 ± 1.84 a | 72.86 ± 3.25 b | |
| 2015 (60; 60; 60) | 13.09 ± 1.59 a | 14.65 ± 1.80 a | 28.10 ± 2.40 b |
| 2016 (60; 60; 60) | 25.29 ± 2.40 a | 27.57 ± 1.69 a | 33.86 ± 2.40 b |
| 2017 (60; 60; 60) | 12.43 ± 1.69 a | 15.43 ± 1.84 a | 41.15 ± 2.82 b |
| Year (n) | Control | Net | Rainproof Net |
|---|---|---|---|
| Set fruits (average number per cluster): | |||
| 2012 (720; 720) | 1.35 ± 0.03 a | 1.19 ± 0.03 b | |
| 2013 (720; 720) | 0.97 ± 0.04 | 0.98 ± 0.04 | |
| 2014 (718; 412) | 0.85 ± 0.03 | 0.77 ± 0.04 | |
| 2015 (480; 480; 480) | 2.40 ± 0.05 a | 2.22 ± 0.06 b | 2.02 ± 0.05 b |
| 2016 (480; 480; 480) | 1.65 ± 0.04 | 1.64 ± 0.04 | 1.68 ± 0.04 |
| 2017 (456; 440; 426) | 2.05 ± 0.04 a | 2.10 ± 0.04 b | 1.91 ± 0.04 c |
| Thinned fruits (number removed per tree): | |||
| 2015 (43; 46; 44) | 60.42 ± 6.40 | 43.30 ± 4.70 | 26.07 ± 2.63 |
| 2016 (24; 24; 24) | 44.71 ± 6.24 | 66.08 ± 6.57 | 53.58 ± 6.20 |
| 2017 (24; 24; 24) | 85.00 ± 12.0 | 46.50 ± 9.63 | 36.33 ± 6.98 |
| Fruit diameter at harvest (mm): | |||
| 2014 (80; 73) | 76.22 ± 0.54 a | 79.18 ± 0.67 b | |
| 2015 (120; 119; 120) | 86.33 ± 0.66 a | 89.57 ± 0.60 b | 91.62 ± 0.66 c |
| 2016 (102; 120; 120) | 78.39 ± 0.63 a | 78.94 ± 0.52 a | 84.59 ± 0.50 b |
| 2017 (120; 120; 120) | 81.38 ± 0.52 a | 82.18 ± 0.52 a | 85.94 ± 0.58 b |
| Fruit weight at harvest (g): | |||
| 2015 (120; 120; 120) | 244.73 ± 5.40 a | 269.33 ± 5.13 b | 291.11 ± 5.76 c |
| 2016 (103; 119; 120) | 186.74 ± 4.25 a | 189.74 ± 3.50 a | 231.23 ± 4.13 b |
| 2017 (120; 120; 120) | 213.36 ± 4.04a | 218.39 ± 4.32 a | 249.99 ± 4.95 b |
| Estimated (2015–2016) or total yield (kg/tree): | |||
| 2015 (23; 24; 23) | 17.09 ± 0.97 | 14.93 ± 0.98 | 13.98 ± 0.88 |
| 2016 (24; 24; 24) | 12.45 ± 1.07 | 14.15 ± 0.67 | 16.10 ± 0.84 |
| 2017 (24; 24; 23) | 11.87 ± 0.89 | 10.61 ± 0.84 | 9.63 ± 1.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chouinard, G.; Veilleux, J.; Pelletier, F.; Larose, M.; Philion, V.; Joubert, V.; Cormier, D. Impact of Exclusion Netting Row Covers on ‘Honeycrisp’ Apple Trees Grown under Northeastern North American Conditions: Effects on Photosynthesis and Fruit Quality. Insects 2019, 10, 214. https://doi.org/10.3390/insects10070214
Chouinard G, Veilleux J, Pelletier F, Larose M, Philion V, Joubert V, Cormier D. Impact of Exclusion Netting Row Covers on ‘Honeycrisp’ Apple Trees Grown under Northeastern North American Conditions: Effects on Photosynthesis and Fruit Quality. Insects. 2019; 10(7):214. https://doi.org/10.3390/insects10070214
Chicago/Turabian StyleChouinard, Gérald, Jonathan Veilleux, Francine Pelletier, Mikael Larose, Vincent Philion, Valentin Joubert, and Daniel Cormier. 2019. "Impact of Exclusion Netting Row Covers on ‘Honeycrisp’ Apple Trees Grown under Northeastern North American Conditions: Effects on Photosynthesis and Fruit Quality" Insects 10, no. 7: 214. https://doi.org/10.3390/insects10070214
APA StyleChouinard, G., Veilleux, J., Pelletier, F., Larose, M., Philion, V., Joubert, V., & Cormier, D. (2019). Impact of Exclusion Netting Row Covers on ‘Honeycrisp’ Apple Trees Grown under Northeastern North American Conditions: Effects on Photosynthesis and Fruit Quality. Insects, 10(7), 214. https://doi.org/10.3390/insects10070214