Impacts of Deciduous Leaf Litter and Snow Presence on Nymphal Ixodes scapularis (Acari: Ixodidae) Overwintering Survival in Coastal New England, USA



 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Treatment Assignments
2.3. Statistical Procedures
2.3.1. Two-Way Analysis of Variance
2.3.2. Multiple Linear Regression
2.3.3. One-Way Repeated Measures ANOVA
3. Results
3.1. Two-Way Analysis of Variance
3.2. Multiple Linear Regression
3.3. One-Way Repeated Measures ANOVA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J.; et al. Vital Signs: Trends in Reported Vectorborne Disease Cases United States and Territories, 2004–2016. Mmwr Morb. Mortal Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Telford, S.R.; Armstrong, P.M.; Katavolos, P.; Foppa, I.; Wilson, M.L.; Spielman, A. A new tick-borne encephalitis-like virus infecting New England deer ticks. Ixodes dammini. Emerg. Infect. Dis. 1997, 3, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Ebel, G.D. Update on Powassan virus: emergence of a North American tick-borne Flavivirus. In Annual Review of Entomology; Berenbaum, M., Cardé, R.T., Robinson, G.E., Eds.; Annual Reviews: Palto Alto, CA, USA, 2010; Volume 55, pp. 95–110. [Google Scholar]
- Krause, P.J.; Narasimhan, S.; Wormser, G.P.; Rollend, L.; Fikrig, E.; Lepore, T.; Barbour, A.; Fish, D. Human Borrelia miyamotoi infection in the United States. N. Eng. J. Med. 2013, 368, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L. The blacklegged tick, Ixodes scapularis: An increasing public health concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.A. Summary of notifiable infectious diseases and conditions—United States, 2014. Mmwr. Morb. Mortal Wkly. Rep 2016, 63, 1–152. [Google Scholar] [CrossRef] [PubMed]
- Lyme, D. Available online: https://www.cdc.gov/lyme/index.html (accessed on 23 May 2019).
- Kilpatrick, H.J.; LaBonte, A.M.; Stafford, K.C., III. The relationship between deer density, tick abundance, and human cases of Lyme disease in a residential community. J. Med. Entomol. 2014, 51, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Linske, M.A.; Williams, S.C.; Stafford, K.C., III; Ortega, I.M. Ixodes scapularis (Acari: Ixodidae) reservoir host diversity and abundance impacts on dilution of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) in residential and woodland habitats in Connecticut, United States. J. Med. Entomol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Miller, M.C.; Hazler, K.R. Causes and consequences of tick (Ixodes scapularis) burdens on white-footed mice (Peromyscus leucopus). J. Mammal. 1996, 77. [Google Scholar] [CrossRef]
- Schmidt, K.A.; Ostfeld, R.S.; Schauber, E.M. Infestation of Peromyscus leucopus and Tamius striatus by Ixodes scapularis (Acari: Ixodidae) in relation to the abundance of hosts and parasites. J. Med. Entomol. 1999, 36, 749–757. [Google Scholar] [CrossRef]
- Wilson, M.L.; Levine, J.F.; Spielman, A. Effect of deer reduction on abundance of the deer tick (Ixodes dammini). Yale J. Biol. Med. 1984, 57, 697–705. [Google Scholar]
- Wilson, M.L.; Adler, G.H.; Spielman, A. Correlation between abundance of deer and that of the deer tick, Ixodes dammini (Acari: Ixodidae). Ann. Entomol. Soc. Am. 1985, 78, 172–176. [Google Scholar] [CrossRef]
- Wilson, M.L.; Telford, S.R., III; Piesman, J.; Spielman, A. Reduced abundance of immature Ixodes dammini (Acari: Ixodidae) following elimination of deer. J. Med. Entomol. 1988, 25, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C., III; DeNicola, A.J.; Kilpatrick, H.J. Reduced abundance of Ixodes scapularis (Acari: Ixodidae) and the tick parasitoid Ixodiphagus hookeri (Hymenoptera: Encyrtidae) with reduction of white-tailed deer. J. Med. Entomol. 2003, 40, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Rand, P.W.; Lubelczyk, C.; Holman, M.S.; Combe, E.H.; Smith, R.P., III. Abundance of Ixodes scapularis (Acari: Ixodidae) after the complete removal of deer from an isolated island, endemic for Lyme disease. J. Med. Entomol. 2004, 41, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Daniels, T.J.; Fish, D.; Schwartz, I. Reduced abundance of Ixodes scapularis (Acari: Ixodidae) and Lyme disease risk by deer exclusion. J. Med. Entomol. 1993, 30, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Daniels, T.J.; Fish, D. Effect of deer exclusion on the abundance of immmature Ixodes scapularis (Acari: Ixodidae) parasitizing small and medium-sized mammals. J. Med. Entomol. 1995, 32, 5–11. [Google Scholar] [CrossRef] [PubMed]
- LoGiudice, K.; Duerr, S.T.K.; Newhouse, M.J.; Schmidt, K.A.; Killilea, M.E.; Ostfeld, R.S. Impact of host community composition on Lyme disease risk. Ecology 2008, 89. [Google Scholar] [CrossRef]
- Williams, S.C.; Stafford, K.C.; Molaei, G.; Linske, M.A. Integrated control of nymphal Ixodes scapularis: Effectiveness of white-tailed deer reduction, the entomopathogenic fungus Metarhizium anisopliae, and fipronil-based rodent bait boxes. Vector Borne Zoonotic Dis. 2018, 18, 55–64. [Google Scholar] [CrossRef]
- Brunner, J.L.; Ostfeld, R.S. Multiple causes of variable tick burdens on small-mammal hosts. Ecology 2008, 89, 2259–2272. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Schauber, E.M.; Canham, C.D.; Keesing, F.; Jones, C.G.; Wolff, J.O. Effects of acorn production and mouse abundance on abundance and Borrelia burgdorferi infection prevalence of nymphal Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001, 1, 55–63. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Canham, C.D.; Oggenfuss, K.; Winchcombe, R.J.; Keesing, F. Climate, deer, rodents, and acorns as determinants of variation in Lyme-disease risk. Plos Biol. 2006, 4, e145. [Google Scholar] [CrossRef] [PubMed]
- Vandyk, J.K.; Bartholomew, D.M.; Rowley, W.A.; Platt, K.B. Survival of Ixodes scapularis (Acari: Ixodidae) exposed to cold. J. Med. Entomol. 1996, 33, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Hayes, L.E.; Scott, J.A.; Stafford, K.C., III. Influences of weather on Ixodes scapularis nymphal densities at long-term study sites in Connecticut. Ticks Tick-Borne Dis. 2015, 6, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.L.; Killilea, M.; Ostfeld, R.S. Overwintering survival of nymphal Ixodes scapularis (Acari: Ixodidae) under natural conditions. J. Med. Entomol. 2014, 49, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C., III. Survival of immature Ixodes scapularis (Acari: Ixodidae) at different relative humidities. J. Med. Entomol. 1994, 31, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Burks, C.S.; Stewart, R.L.; Needham, G.R.; Lee, R.E. Cold hardiness in the ixodid ticks (Ixodidae). In Acarology IX: Volume 1, Proceedings; Mitchell, R., Horn, D.J., Needham, G.R., Welborne, W.C., Eds.; Ohio Biological Survey: Columbus, OH, USA, 1996; pp. 85–87. [Google Scholar]
- Burgdorfer, W. Vector/host relationship of the Lyme disease spirochete, Borrelia burgdorferi. Rhem. Dis. Clin. N Am. 1989, 15, 775–787. [Google Scholar]
- Oliver, J.H. Lyme borreliosis in the southern United States: A review. J. Parasitol. 1996, 82, 926–935. [Google Scholar] [CrossRef]
- NRCS Connecticut. Soils. Natural Resources Conservation Service. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/main/ct/soils/ (accessed on 23 May 2019).
- Service, S.C. Soil Survey, Cumberland County, Maine; United States Dept. of Agriculture: Washington, DC, USA, 1974.
- Kleinbaum, D.G.; Kupper, L.L.; Nizam, A.; Rosenberg, E.S. Applied Regression Analysis and Other Multivariable Methods, 5th ed.; Cengage Learning: Boston, MA, USA, 2013; pp. 438–480. [Google Scholar]
- Planet, C.A.C.; Wake, C. Indicators of Climate Change in the Northeast 2005. Available online: http://www.cleanair-coolplanet.org/information/pdf/indicators.pdf (accessed on 11 June 2019).
- Thibeault, J.M.; Seth, A. Changing climate extremes in the Northeast United States: Observations and projections from CMIP5. Clim. Chang. 2014, 127, 273–287. [Google Scholar] [CrossRef]
- Ahrens, C.D. Meteorology Today: An Introduction to Weather, Climate, and the Environment; Cengage Learning: Belmont, CA, USA, 2012; p. 633. [Google Scholar]
- Berger, K.A.; Ginsberg, H.S.; Gonzalez, L.; Mather, T.N. Relative humidity and activity patterns of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2014, 51, 769–776. [Google Scholar] [CrossRef]
- Neelakanta, G.; Sultana, H.; Fish, D.; Anderson, J.F.; Fikrig, E. Anaplasma phagocytophilum induces Ixodes scapularis ticks to express an antifreeze glycoprotein gene that enhances their survival in the cold. J. Clin. Investig. 2010, 120, 3179–3190. [Google Scholar] [CrossRef]
- Carroll, M.C.; Ginsberg, H.S.; Hyland, K.E.; Hu, R. Distributoin of Ixodes dammini (Acari: Ixodidae) in residential suburban landscape by area application of insecticides. J. Med. Entomol. 1992, 30, 107–113. [Google Scholar]
- Stafford, K.C.; Magnarelli, L.A. Spatial and temporal patterns of Ixodes scapularis (Acari: Ixodidae) in southcentral Connecticut. J. Med. Entomol. 1993, 30, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C., III. Tick Management Handbook: An Integrated Guide for Homeowners, Pest Control Operators, and Public Health Officials For The Prevention of Tick-Associated Disease, 2nd ed.; The Connecticut Agricultural Experiment Station: New Haven, CT, USA, 2007; p. 78. [Google Scholar]
- Ogden, N.H.; Lindsay, L.R. Effects of climate and climate change on vectors and vector-borne diseases: Ticks are different. Trends Parasitol. 2016, 32, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Roy-Dufresne, E.; Logan, T.; Simon, J.A.; Chmura, G.L.; Millien, V. Poleward expansion of the white-footed mouse (Peromyscus leucopus) under climate change: Implications for the spread of Lyme disease. PLoS ONE 2013, 8, e80724. [Google Scholar] [CrossRef] [PubMed]
- Pérez de León, A.A.; Teel, P.D.; Li, A.; Ponnusamy, L.; Roe, R.M. Advancing integrated tick management to mitigate burden of tick-borne diseases. Outlooks Pest Manag. 2014, 25, 382–389. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Stafford, K.C., III. Management of ticks and tick-borne diseases. In Tickborne Diseases of Humans; Goodman, J.L., Dennis, D.T., Sonenshine, D.E., Eds.; ASM Press: Washington, DC, USA, 2005; pp. 65–86. [Google Scholar]
- Stafford, K.C., III. Forum: Management of Lyme Disease (Integrated Pest Management for Ixodes scapularis: Principals and Prospects). In Ecology and Environmental Management of Lyme Disease; Ginsberg, H.S., Ed.; Rutgers University Press: New Brunswick, NJ, USA, 1993; pp. 178–181. [Google Scholar]
- Ginsberg, H.S. Integrated pest management and allocation of control efforts for vector-borne diseases. J. Vector Ecol. 2001, 26, 32–38. [Google Scholar] [PubMed]
- Ginsberg, H.S. Transmission risk of Lyme disease and implications for tick management. Am. J. Epidemiol. 1993, 138, 65–73. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
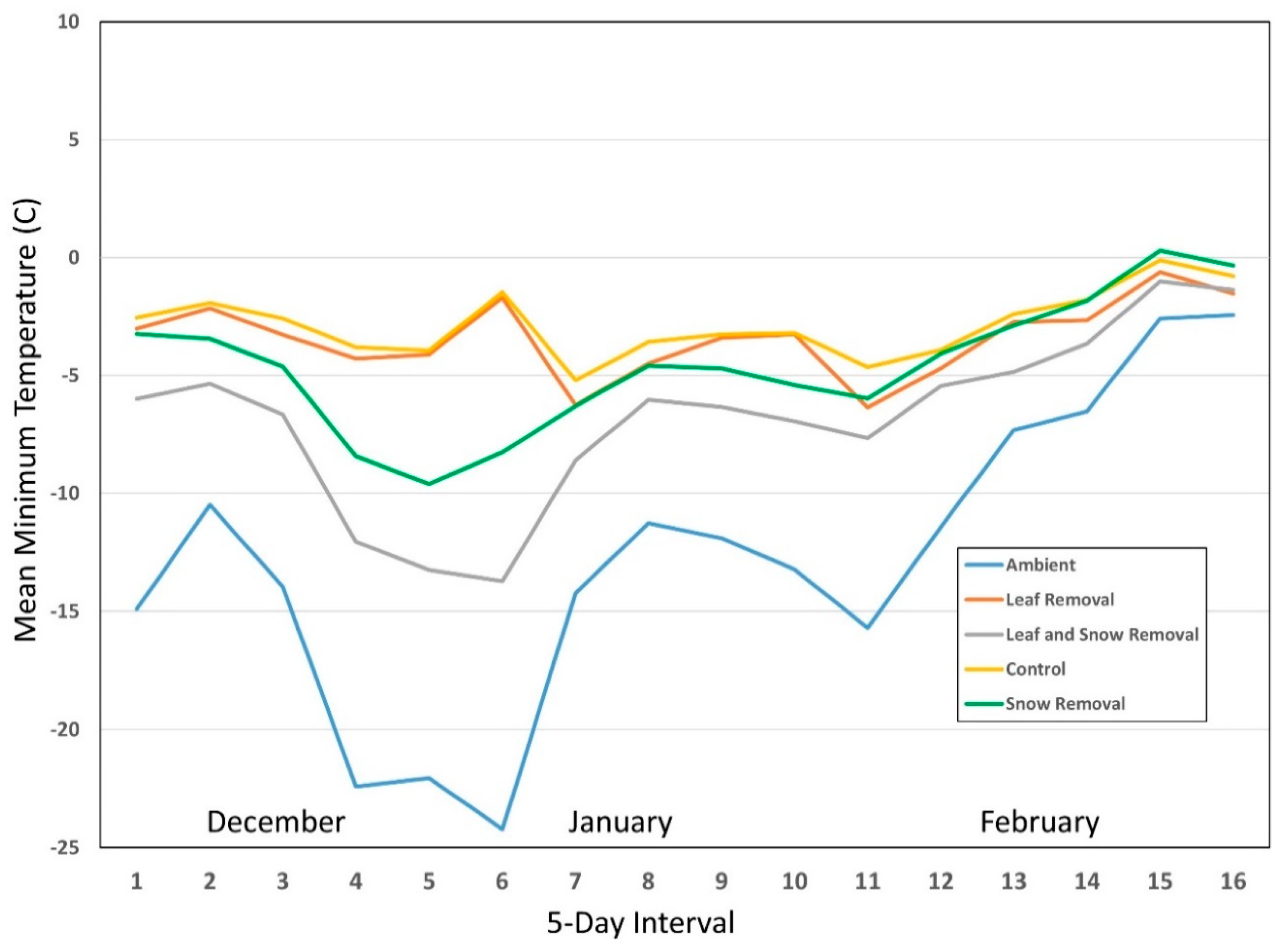{kind=link}
| Treatments | Y1 | Y2 | Y3 | Total | |||
|---|---|---|---|---|---|---|---|
| CT | ME | CT | ME | CT | ME | ||
| Control | 94% 249 | 21% 56 | 61% 166 | 53% 145 | 54% 142 | 74% 167 | 59% A 925 |
| SR | 86% 239 | 23% 62 | 63% 153 | 28% 72 | 47% 123 | 33% 98 | 46% AB 747 |
| LR | 86% 226 | 3% 7 | 53% 138 | 24% 63 | 44% 111 | 52% 139 | 44% B 684 |
| LRSR | 77% 207 | 3% 7 | 40% 106 | 17% 44 | 52% 132 | 31% 84 | 36% B 580 |
| Treatment | N | Y1 | Y2 | Y3 |
|---|---|---|---|---|
| Control | 12 | −1.4 A | −1.7 A | −2.9 A |
| SR | 12 | −2.4 AB | −2.3 A | −4.6 AB |
| LR | 12 | −2.9 B | −2.7 AB | −3.3 A |
| LRSR | 12 | −3.8 BC | −4.5 BC | −6.2 BC |
| Ambient | 6 | −8.9 C | −9.8 C | −12.6 C |
| Treatment | N | Y1 | Y2 | Y3 |
|---|---|---|---|---|
| Control | 12 | 0.7 A | 0.4 A | −2.1 A |
| SR | 12 | 1.0 AB | 0.9 A | 0.1 AB |
| LR | 12 | 1.0 AB | 0.8 AB | −0.6 A |
| LRSR | 12 | 1.4 B | 1.1 B | 0.3 AB |
| Ambient | 6 | 1.2 B | 1.0 B | −0.2 B |
| Location | Year | Month | # of Non-Snow Days | # of Snow Days | Avg. Monthly Snow Accumulation (cm) |
|---|---|---|---|---|---|
| CT | 1 | Dec | 16 | 0 | 0.0 |
| CT | 1 | Jan | 25 | 6 | 8.5 |
| CT | 1 | Feb | 14 | 15 | 12.9 |
| CT | 2 | Dec | 13 | 2 | 1.0 |
| CT | 2 | Jan | 26 | 5 | 9.2 |
| CT | 2 | Feb | 16 | 13 | 18.2 |
| CT | 3 | Dec | 11 | 4 | 2.0 |
| CT | 3 | Jan | 16 | 15 | 11.4 |
| CT | 3 | Feb | 25 | 4 | 7.2 |
| Total | 162 | 64 | 7.8 | ||
| ME | 1 | Dec | 16 | 0 | 0.0 |
| ME | 1 | Jan | 0 | 31 | 9.6 |
| ME | 1 | Feb | 3 | 26 | 10.9 |
| ME | 2 | Dec | 0 | 16 | 10.7 |
| ME | 2 | Jan | 6 | 25 | 26.4 |
| ME | 2 | Feb | 0 | 28 | 45.6 |
| ME | 3 | Dec | 0 | 16 | 20.2 |
| ME | 3 | Jan | 0 | 31 | 57.4 |
| ME | 3 | Feb | 0 | 28 | 16.1 |
| Total | 25 | 201 | 21.9 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linske, M.A.; Stafford, K.C., III; Williams, S.C.; Lubelczyk, C.B.; Welch, M.; Henderson, E.F. Impacts of Deciduous Leaf Litter and Snow Presence on Nymphal Ixodes scapularis (Acari: Ixodidae) Overwintering Survival in Coastal New England, USA. Insects 2019, 10, 227. https://doi.org/10.3390/insects10080227
Linske MA, Stafford KC III, Williams SC, Lubelczyk CB, Welch M, Henderson EF. Impacts of Deciduous Leaf Litter and Snow Presence on Nymphal Ixodes scapularis (Acari: Ixodidae) Overwintering Survival in Coastal New England, USA. Insects. 2019; 10(8):227. https://doi.org/10.3390/insects10080227
Chicago/Turabian StyleLinske, Megan A., Kirby C. Stafford, III, Scott C. Williams, Charles B. Lubelczyk, Margret Welch, and Elizabeth F. Henderson. 2019. "Impacts of Deciduous Leaf Litter and Snow Presence on Nymphal Ixodes scapularis (Acari: Ixodidae) Overwintering Survival in Coastal New England, USA" Insects 10, no. 8: 227. https://doi.org/10.3390/insects10080227
APA StyleLinske, M. A., Stafford, K. C., III, Williams, S. C., Lubelczyk, C. B., Welch, M., & Henderson, E. F. (2019). Impacts of Deciduous Leaf Litter and Snow Presence on Nymphal Ixodes scapularis (Acari: Ixodidae) Overwintering Survival in Coastal New England, USA. Insects, 10(8), 227. https://doi.org/10.3390/insects10080227