Detection of Leishmania and Trypanosoma DNA in Field-Caught Sand Flies from Endemic and Non-Endemic Areas of Leishmaniasis in Southern Thailand


 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sand Fly Collection, Sample Preparation, and Identification
2.3. DNA Extraction
2.4. Detection of Leishmania and Trypanosoma DNA in the Sand Fly Samples
2.5. Molecular Cloning and Nucleotide Sequencing
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.; Clode, P.L.; Peacock, C.; Thompson, R.C.A. Chapter Two—Host–Parasite Relationships and Life Histories of Trypanosomes in Australia. In Advances in Parasitology; Rollinson, D., Stothard, J.R., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 97, pp. 47–109. [Google Scholar]
- Njagu, Z.; Mihok, S.; Kokwaro, E.; Verloo, D. Isolation of Trypanosoma brucei from the monitor lizard (Varanus niloticus) in an endemic focus of rhodesian sleeping sickness in Kenya. Acta Trop. 1999, 72, 137–148. [Google Scholar] [CrossRef]
- Ferreira, R.C.; De Souza, A.A.; Freitas, R.A.; Campaner, M.; Takata, C.S.; Barrett, T.V.; Shaw, J.J.; Teixeira, M.M. A phylogenetic lineage of closely related trypanosomes (Trypanosomatidae, Kinetoplastida) of anurans and sand flies (Psychodidae, Diptera) sharing the same ecotopes in brazilian amazonia. J. Eukaryot. Microbiol. 2008, 55, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Lemos, M.; Morais, D.H.; Carvalho, V.T.; D’Agosto, M. First record of Trypanosoma chattoni in Brazil and occurrence of other Trypanosoma species in Brazilian frogs (Anura, Leptodactylidae). J. Parasitol. 2008, 94, 148–151. [Google Scholar] [CrossRef]
- Viola, L.B.; Campaner, M.; Takata, C.S.; Ferreira, R.C.; Rodrigues, A.C.; Freitas, R.A.; Duarte, M.R.; Grego, K.F.; Barrett, T.V.; Camargo, E.P.; et al. Phylogeny of snake trypanosomes inferred by SSU rDNA sequences, their possible transmission by phlebotomines, and taxonomic appraisal by molecular, cross-infection and morphological analysis. Parasitology 2008, 135, 595–605. [Google Scholar] [CrossRef]
- Jariyapan, N.; Daroontum, T.; Jaiwong, K.; Chanmol, W.; Intakhan, N.; Sor-Suwan, S.; Siriyasatien, P.; Somboon, P.; Bates, M.D.; Bates, P.A. Leishmania (Mundinia) orientalis n. sp. (Trypanosomatidae), a parasite from Thailand responsible for localised cutaneous leishmaniasis. Parasites Vectors 2018, 11, 351. [Google Scholar] [CrossRef]
- Chiewchanvit, S.; Tovanabutra, N.; Jariyapan, N.; Bates, M.D.; Mahanupab, P.; Chuamanochan, M.; Tantiworawit, A.; Bates, P.A. Chronic generalized fibrotic skin lesions from disseminated leishmaniasis caused by Leishmania martiniquensis in two patients from northern Thailand infected with HIV. Br. J. Dermatol. 2015, 173, 663–670. [Google Scholar] [CrossRef]
- Thisyakorn, U.; Jongwutiwes, S.; Vanichsetakul, P.; Lertsapcharoen, P. Visceral leishmaniasis: The first indigenous case report in Thailand. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 23–24. [Google Scholar] [CrossRef]
- Maharom, P.; Siripattanapipong, S.; Mungthin, M.; Naaglor, T.; Sukkawee, R.; Pudkorn, R.; Wattana, W.; Wanachiwanawin, D.; Areechokchai, D.; Leelayoova, S. Visceral leishmaniasis caused by Leishmania infantum in Thailand. Southeast Asian J. Trop. Med. Public Health 2008, 39, 988–990. [Google Scholar]
- Leelayoova, S.; Siripattanapipong, S.; Manomat, J.; Piyaraj, P.; Tan-Ariya, P.; Bualert, L.; Mungthin, M. Leishmaniasis in Thailand: A Review of Causative Agents and Situations. Am. J. Trop. Med. Hyg. 2017, 96, 534–542. [Google Scholar] [CrossRef] [Green Version]
- Milocco, C.; Kamyingkird, K.; Desquesnes, M.; Jittapalapong, S.; Herbreteau, V.; Chaval, Y.; Douangboupha, B.; Morand, S. Molecular demonstration of Trypanosoma evansi and Trypanosoma lewisi DNA in wild rodents from Cambodia, Lao PDR and Thailand. Transbound. Emerg. Dis. 2013, 60, 17–26. [Google Scholar] [CrossRef]
- Sim, S.T.; Wiwanitkit, V. Emerging human trypanosomiasis in Southeast Asia: A coming new threat? J. Nat. Sci. 2015, 1, e29. [Google Scholar]
- Sarataphan, N.; Vongpakorn, M.; Nuansrichay, B.; Autarkool, N.; Keowkarnkah, T.; Rodtian, P.; Stich, R.W.; Jittapalapong, S. Diagnosis of a Trypanosoma lewisi-like (Herpetosoma) infection in a sick infant from Thailand. J. Med. Microbiol. 2007, 56, 1118–1121. [Google Scholar] [CrossRef]
- Apiwathnasorn, C.; Sucharit, S.; Rongsriyam, Y.; Leemingsawat, S.; Kerdpibule, V.; Deesin, T.; Surathin, K.; Vutikes, S.; Punavuthi, N. A brief survey of phlebotomine sandflies in Thailand. Southeast Asian J. Trop. Med. Public Health 1989, 20, 429–432. [Google Scholar]
- Apiwathnasorn, C.; Samung, Y.; Prummongkol, S.; Phayakaphon, A.; Panasopolkul, C. Cavernicolous species of phlebotomine sand flies from Kanchanaburi Province, with an updated species list for Thailand. Southeast Asian J. Trop. Med. Public Health 2011, 42, 1405–1409. [Google Scholar]
- Artemiev, M.A. Classification of the subfamily Phlebotominae. Parassitologia 1991, 33, 69–77. [Google Scholar]
- Marcondes, C.B. A proposal of generic and subgeneric abbreviations for Phlebotomine sandflies (Diptera: Psychodidae: Phlebotominae) of the World. Entomol. News 2007, 118, 351–356. [Google Scholar] [CrossRef]
- Galati, E.A.B. Classificação de Phlebotominae, in Flebotomíneos do Brasil; Rangel, E.F., Lainson, R., Eds.; Fiocruz: Rio de Janeiro, Brazil, 2003; pp. 23–51. [Google Scholar]
- Sukra, K.; Kanjanopas, K.; Amsakul, S.; Rittaton, V.; Mungthin, M.; Leelayoova, S. A survey of sandflies in the affected areas of leishmaniasis, southern Thailand. Parasitol. Res. 2013, 112, 297–302. [Google Scholar] [CrossRef]
- Kanjanopas, K.; Siripattanapipong, S.; Ninsaeng, U.; Hitakarun, A.; Jitkaew, S.; Kaewtaphaya, P.; Tan-ariya, P.; Mungthin, M.; Charoenwong, C.; Leelayoova, S. Sergentomyia (Neophlebotomus) gemmea, a potential vector of Leishmania siamensis in southern Thailand. BMC Infect. Dis. 2013, 13, 333. [Google Scholar] [CrossRef]
- Phumee, A.; Tawatsin, A.; Thavara, U.; Pengsakul, T.; Thammapalo, S.; Depaquit, J.; Gay, F.; Siriyasatien, P. Detection of an Unknown Trypanosoma DNA in a Phlebotomus stantoni (Diptera: Psychodidae) Collected From Southern Thailand and Records of New Sand Flies With Reinstatement of Sergentomyia hivernus Raynal & Gaschen, 1935 (Diptera: Psychodidae). J. Med. Entomol. 2017, 54, 429–434. [Google Scholar] [CrossRef]
- Lewis, D.J. The phlebotomine sandflies (Diptera: Psychodidae) of the oriental region. Bull. Br. Mus. (Nat. Hist.) B. Entomol. 1978, 37, 217–343. [Google Scholar] [CrossRef]
- Depaquit, J.; Vongphayloth, K.; Siriyasatien, P.; Polseela, R.; Phumee, A.; Loyer, M.; Vol, A.; Varlot, G.; Rahola, N.; Brey, P.T.; et al. On the true identity of Sergentomyia gemmea and description of a closely related species: Se. raynali n. sp. Med. Vet. Entomol. 2019. [Google Scholar] [CrossRef]
- Spanakos, G.; Piperaki, E.T.; Menounos, P.G.; Tegos, N.; Flemetakis, A.; Vakalis, N.C. Detection and species identification of old world Leishmania in clinical samples using a PCR-based method. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 46–53. [Google Scholar] [CrossRef]
- Noyes, H.A.; Stevens, J.R.; Teixeira, M.; Phelan, J.; Holz, P. A nested PCR for the ssrRNA gene detects Trypanosoma binneyi in the platypus and Trypanosoma sp. in wombats and kangaroos in Australia. Int. J. Parasitol. 1999, 29, 331–339. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bualert, L.; Charungkiattikul, W.; Thongsuksai, P.; Mungthin, M.; Siripattanapipong, S.; Khositnithikul, R.; Naaglor, T.; Ravel, C.; El Baidouri, F.; Leelayoova, S. Case Report: Autochthonous disseminated dermal and visceral leishmaniasis in an AIDS patient, southern Thailand, caused by Leishmania siamensis. Am. J. Trop. Med. Hyg. 2012, 86, 821–824. [Google Scholar] [CrossRef]
- Chusri, S.; Hortiwakul, T.; Silpapojakul, K.; Siriyasatien, P. Case Report: Consecutive cutaneous and visceral leishmaniasis manifestations involving a novel Leishmania species in two HIV patients in Thailand. Am. J. Trop. Med. Hyg. 2012, 87, 76–80. [Google Scholar] [CrossRef]
- Jungudomjaroen, S.; Phumee, A.; Chusri, S.; Kraivichian, K.; Jariyapan, N.; Payungporn, S.; Siriyasatien, P. Detection of Leishmania martiniquensis DNA in various clinical samples by quantitative PCR. Trop. Biomed. 2015, 32, 736–744. [Google Scholar]
- Noppakun, N.; Kraivichian, K.; Siriyasatien, P. Disseminated dermal leishmaniasis caused by Leishmania siamensis in a systemic steroid therapy patient. Am. J. Trop. Med. Hyg. 2014, 91, 869–870. [Google Scholar] [CrossRef]
- Osatakul, S.; Mungthin, M.; Siripattanapipong, S.; Hitakarun, A.; Kositnitikul, R.; Naaglor, T.; Leelayoova, S. Recurrences of visceral leishmaniasis caused by Leishmania siamensis after treatment with amphotericin B in a seronegative child. Am. J. Trop. Med. Hyg. 2014, 90, 40–42. [Google Scholar] [CrossRef]
- Phumee, A.; Kraivichian, K.; Chusri, S.; Noppakun, N.; Vibhagool, A.; Sanprasert, V.; Tampanya, V.; Wilde, H.; Siriyasatien, P. Detection of Leishmania siamensis DNA in saliva by polymerase chain reaction. Am. J. Trop. Med. Hyg. 2013, 89, 899–905. [Google Scholar] [CrossRef]
- Phumee, A.; Chusri, S.; Kraivichian, K.; Wititsuwannakul, J.; Hortiwakul, T.; Thavara, U.; Silpapojakul, K.; Siriyasatien, P. Multiple Cutaneous Nodules in an HIV-Infected Patient. PLoS Negl. Trop. Dis. 2014, 8, e3291. [Google Scholar] [CrossRef]
- Pothirat, T.; Tantiworawit, A.; Chaiwarith, R.; Jariyapan, N.; Wannasan, A.; Siriyasatien, P.; Supparatpinyo, K.; Bates, M.D.; Kwakye-Nuako, G.; Bates, P.A. First isolation of Leishmania from Northern Thailand: Case report, identification as Leishmania martiniquensis and phylogenetic position within the Leishmania enriettii complex. PLoS Negl. Trop. Dis. 2014, 8, e3339. [Google Scholar] [CrossRef]
- Siriyasatien, P.; Chusri, S.; Kraivichian, K.; Jariyapan, N.; Hortiwakul, T.; Silpapojakul, K.; Pym, A.M.; Phumee, A. Early detection of novel Leishmania species DNA in the saliva of two HIV-infected patients. BMC Infect. Dis. 2016, 16, 89. [Google Scholar] [CrossRef]
- Suankratay, C.; Suwanpimolkul, G.; Wilde, H.; Siriyasatien, P. Case Report: Autochthonous visceral leishmaniasis in a human immunodeficiency virus (HIV)-infected patient: The first in Thailand and review of the literature. Am. J. Trop. Med. Hyg. 2010, 82, 4–8. [Google Scholar] [CrossRef]
- Sukmee, T.; Siripattanapipong, S.; Mungthin, M.; Worapong, J.; Rangsin, R.; Samung, Y.; Kongkaew, W.; Bumrungsana, K.; Chanachai, K.; Apiwathanasorn, C.; et al. A suspected new species of Leishmania, the causative agent of visceral leishmaniasis in a Thai patient. Int. J. Parasitol. 2008, 38, 617–622. [Google Scholar] [CrossRef]
- Ready, P. Biology of phlebotomine sand flies as vectors of disease agents. Annu. Rev. Entomol. 2013, 58, 227–250. [Google Scholar] [CrossRef]
- Maia, C.; Depaquit, J. Can Sergentomyia (Diptera, Psychodidae) play a role in the transmission of mammal-infecting Leishmania? Parasite 2016, 23, 55. [Google Scholar] [CrossRef]
- Mukherjee, S.; Hassan, M.Q.; Ghosh, A.; Ghosh, K.N.; Bhattacharya, A.; Adhya, S. Leishmania DNA in Phlebotomus and Sergentomyia species during a kala-azar epidemic. Am. J. Trop. Med. Hyg. 1997, 57, 423–425. [Google Scholar] [CrossRef]
- Arvizi, P.; Amirkhani, A. Mitochondrial DNA characterization of Sergentomyia sintoni populations and finding mammalian Leishmania infections in this sandfly by using ITS-rDNA gene. Iran. J. Vet. Res. 2008, 9, 9–18. [Google Scholar]
- Campino, L.; Cortes, S.; Dionísio, L.; Neto, L.; Afonso, M.O.; Maia, C. The first detection of Leishmania major in naturally infected Sergentomyia minuta in Portugal. Mem. Inst. Oswaldo Cruz 2013, 108, 516–518. [Google Scholar] [CrossRef]
- Ayari, C.; Ben Othman, S.; Chemkhi, J.; Tabbabi, A.; Fisa, R.; Ben Salah, A.; BenAbderrazak, S. First detection of Leishmania major DNA in Sergentomyia (Sintonius) clydei (Sinton, 1928, Psychodidae: Phlebotominae), from an outbreak area of cutaneous leishmaniasis in Tunisia. Infect. Genet. Evol. 2016, 39, 241–248. [Google Scholar] [CrossRef]
- Aouadi, K.; Ghawar, W.; Salem, S.; Gharbi, M.; Bettaieb, J.; Yazidi, R.; Harrabi, M.; Hamarsheh, O.; Ben Salah, A. First report of naturally infected Sergentomyia minuta with Leishmania major in Tunisia. Parasites Vectors 2015, 8, 649. [Google Scholar] [CrossRef]
- Senghor, M.; Niang, A.; Depaquit, J.; Faye, M.; Ferté, H.; Elguero, E.; Gaye, O.; Alten, B.; Perktas, U.; Cassan, C.; et al. Transmission of Leishmania infantum in the canine leishmaniasis focus of Mont-Rolland, Senegal: Ecological, parasitological and molecular evidence for a possible role of Sergentomyia sand flies. PLoS Negl. Trop. Dis. 2016, 10, e0004940. [Google Scholar] [CrossRef]
- Chusri, S.; Thammapalo, S.; Chusri, S.; Thammapalo, S.; Silpapojakul, K.; Siriyasatien, P. Animal reservoirs and potential vectors of Leishmania siamensis in southern Thailand. Southeast Asian J. Trop. Med. Public Health 2014, 45, 13–19. [Google Scholar]
- Siripattanapipong, S.; Leelayoova, S.; Ninsaeng, U.; Mungthin, M. Detection of DNA of Leishmania siamensis in Sergentomyia (Neophlebotomus) iyengari (Diptera: Psychodidae) and Molecular Identification of Blood Meals of Sand Flies in an Affected Area, Southern Thailand. J. Med. Entomol. 2018, 55, 1277–1283. [Google Scholar] [CrossRef]
- Botero, A.; Cooper, C.; Thompson, C.K.; Clode, P.L.; Rose, K.; Thompson, R.C. Morphological and Phylogenetic Description of Trypanosoma noyesi sp. nov.: An Australian Wildlife Trypanosome within the T. cruzi Clade. Protist 2016, 167, 425–439. [Google Scholar] [CrossRef]
- Kato, H.; Uezato, H.; Sato, H.; Bhutto, A.M.; Soomro, F.R.; Baloch, J.H.; Iwata, H.; Hashiguchi, Y. Natural infection of the sand fly Phlebotomus kazeruni by Trypanosoma species in Pakistan. Parasites Vectors 2010, 3, 10. [Google Scholar] [CrossRef]
- Nzelu, C.O.; Kato, H.; Puplampu, N.; Desewu, K.; Odoom, S.; Wilson, M.D.; Sakurai, T.; Katakura, K.; Boakye, D.A. First detection of Leishmania tropica DNA and Trypanosoma species in Sergentomyia sand flies (Diptera: Psychodidae) from an outbreak area of cutaneous leishmaniasis in Ghana. PLoS Negl. Trop. Dis. 2014, 8, e2630. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
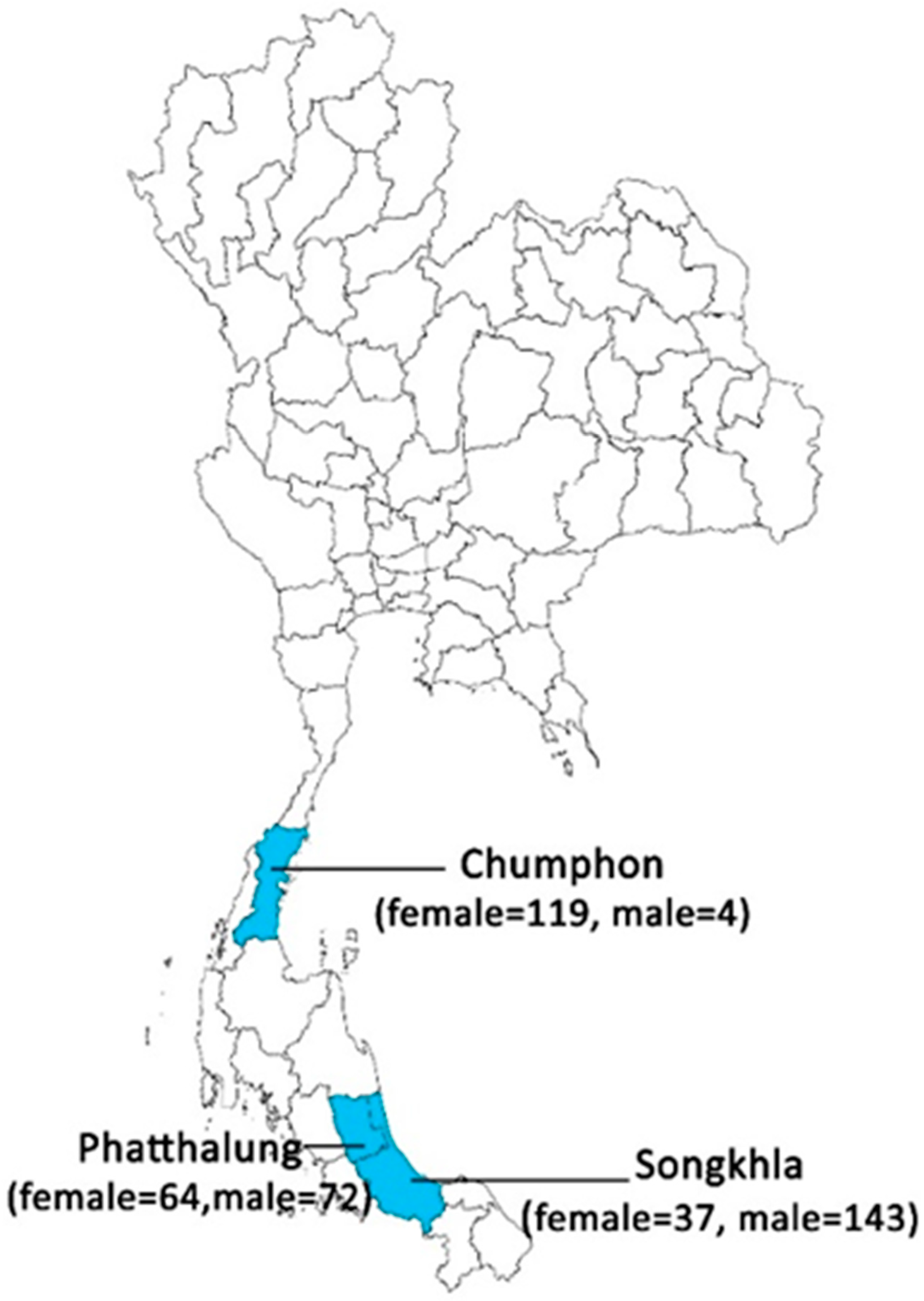
| Species | Provinces | Total | ||
|---|---|---|---|---|
| Songkhla | Phatthalung | Chumphon | ||
| Se. anodontis | 4 | 0 | 48 | 52 |
| Se. khawi | 29 | 50 | 0 | 79 |
| Se. sylvatica | 0 | 0 | 1 | 1 |
| Se. barraudi | 2 | 3 | 0 | 5 |
| Se. indica | 0 | 2 | 0 | 2 |
| Ph. stantoni | 2 | 0 | 0 | 2 |
| Ph. asperulus | 0 | 0 | 14 | 14 |
| Ph. betisi | 0 | 3 | 38 | 41 |
| Ph. kiangsuensis | 0 | 6 | 15 | 21 |
| Ph. major major | 0 | 0 | 2 | 2 |
| Ph. mascomai | 0 | 0 | 1 | 1 |
| Total | 37 | 64 | 119 | 220 |
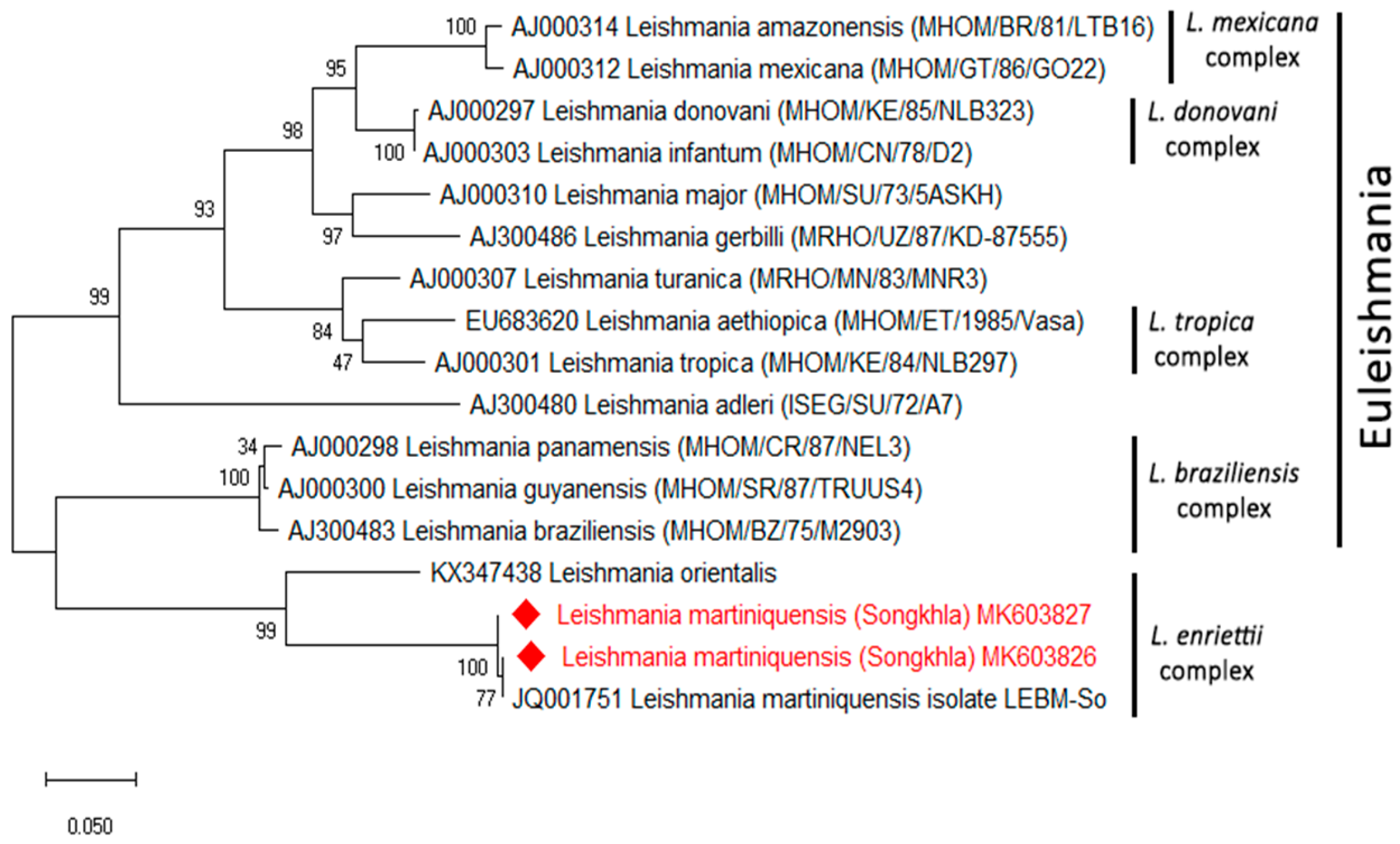
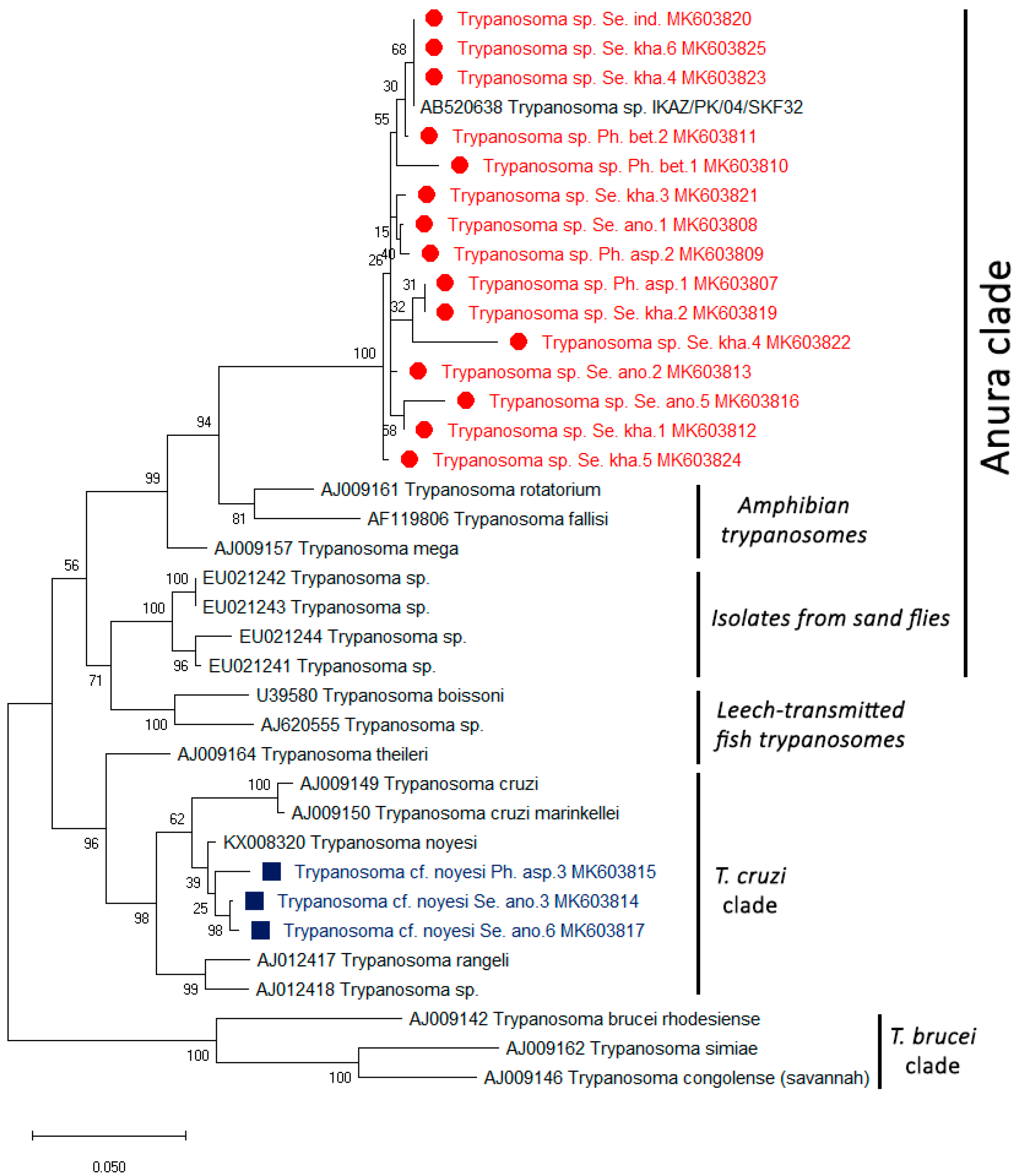
| Province | Positive Sand Fly Species | Detection of Leishmania spp. and Trypanosoma sp. | ||
|---|---|---|---|---|
| L. martiniquensis | T. noyesi | Trypanosoma sp. | ||
| Songkhla (n = 37) | Se. khawi | 2 * | ND | 4 |
| Phatthalung (n = 64) | Se. khawi | ND | ND | 1 |
| Se. indica | ND | ND | 1 | |
| Chumphon (n = 119) | Ph. asperulus | ND | 1 | 2 |
| Se. anodontis | ND | 2 | 4 | |
| Ph. betisi | ND | ND | 2 | |
| Total (n = 220) | 1 | 3 | 14 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srisuton, P.; Phumee, A.; Sunantaraporn, S.; Boonserm, R.; Sor-suwan, S.; Brownell, N.; Pengsakul, T.; Siriyasatien, P. Detection of Leishmania and Trypanosoma DNA in Field-Caught Sand Flies from Endemic and Non-Endemic Areas of Leishmaniasis in Southern Thailand. Insects 2019, 10, 238. https://doi.org/10.3390/insects10080238
Srisuton P, Phumee A, Sunantaraporn S, Boonserm R, Sor-suwan S, Brownell N, Pengsakul T, Siriyasatien P. Detection of Leishmania and Trypanosoma DNA in Field-Caught Sand Flies from Endemic and Non-Endemic Areas of Leishmaniasis in Southern Thailand. Insects. 2019; 10(8):238. https://doi.org/10.3390/insects10080238
Chicago/Turabian StyleSrisuton, Pimpilad, Atchara Phumee, Sakone Sunantaraporn, Rungfar Boonserm, Sriwatapron Sor-suwan, Narisa Brownell, Theerakamol Pengsakul, and Padet Siriyasatien. 2019. "Detection of Leishmania and Trypanosoma DNA in Field-Caught Sand Flies from Endemic and Non-Endemic Areas of Leishmaniasis in Southern Thailand" Insects 10, no. 8: 238. https://doi.org/10.3390/insects10080238
APA StyleSrisuton, P., Phumee, A., Sunantaraporn, S., Boonserm, R., Sor-suwan, S., Brownell, N., Pengsakul, T., & Siriyasatien, P. (2019). Detection of Leishmania and Trypanosoma DNA in Field-Caught Sand Flies from Endemic and Non-Endemic Areas of Leishmaniasis in Southern Thailand. Insects, 10(8), 238. https://doi.org/10.3390/insects10080238