Emigration Effects Induced by Radio Frequency Treatment to Dates Infested by Carpophilus hemipterus

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Dates and Infestation Procedure
2.3. Experimental Procedures
2.4. Insect Emigration Assessment
2.5. Statistical Analyses
3. Results
3.1. Heating Profiles
3.2. Insect Emigration
3.3. Insect Mortality
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aleid, S.M. Date fruit processing and processed products. In Dates: Postharvest Science, Processing Technology and Health Benefits, 1st ed.; Siddiq, M., Aleid, S.M., Kader, A.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 171–202. [Google Scholar] [CrossRef]
- Alhamdan, A.; Hassan, B.; Alkahtani, H.; Abdelkarim, D.; Younis, M. Freezing of fresh Barhi dates for quality preservation during frozen storage. Saudi J. Biol. Sci. 2018, 25, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, D. Date palm arthropod pests and their management in Israel. Phytoparasitica 2008, 36, 411–448. [Google Scholar] [CrossRef]
- Al-Deeb, M.A. Date palm insect and mite pests and their management. In Dates: Production, Processing, Food, and Medicinal Values, 1st ed.; Manickavasagan, A., Mohamed Essa, M., Sukumar, E., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 113–128. [Google Scholar]
- Abo-El-Saad, M.; El-Shafie, H. Insect pests of stored dates and their management. In Dates: Postharvest Science, Processing Technology and Health Benefits, 1st ed.; Siddiq, M., Aleid, S.M., Kader, A.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 81–104. [Google Scholar] [CrossRef]
- Burks, C.S.; Yasin, M.; El-Shafie, H.A.F.; Wakil, W. Pests of stored dates. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges, Sustainability in Plant and Crop Protection; Wakil, W., Romeno Faleiro, J., Miller, T.A., Eds.; Springer International Publishing: Zürich, Switzerland, 2015; pp. 237–286. [Google Scholar] [CrossRef]
- Emekci, M.; Moore, D. Sap beetles. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges, Sustainability in Plant and Crop Protection; Wakil, W., Romeno Faleiro, J., Miller, T.A., Eds.; Springer International Publishing: Zürich, Switzerland, 2015; pp. 205–235. [Google Scholar] [CrossRef]
- Bartelt, R.J.; Dowd, P.F.; Plattner, R.D.; Weisleder, D. Aggregation pheromone of Carpophilus hemipterus. Wind-tunnel bioassay and identification of two novel tetraene hydrocarbons. J. Chem. Ecol. 1990, 16, 1015–1039. [Google Scholar] [CrossRef] [PubMed]
- Yahia, E.M. Date. In The Commercial Storage of Fruits Vegetables and Florist and Nursery Stocks; Agriculture Handbook n. 66; Gross, K.C., Wang, C.Y., Saltveit, M., Eds.; USDA: Beltsville, MD, USA, 2016; pp. 311–314. [Google Scholar]
- Navarro, H.; Navarro, S. Postharvest processing of dates: Drying, disinfestation and storage. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges, Sustainability in Plant and Crop Protection; Wakil, W., Romeno Faleiro, J., Miller, T.A., Eds.; Springer International Publishing: Zürich, Switzerland, 2015; pp. 391–409. [Google Scholar] [CrossRef]
- Navarro, S.; Donahaye, J.E.; Rindner, M.; Azrieli, A. Storage of dates under carbon dioxide atmosphere for quality preservation. In Proceedings of the International Conference Controlled Atmosphere and Fumigation in Stored Products, Fresno, CA, USA, 29 October–3 November 2000; Donahaye, E.J., Navarro, S., Leesch, J.G., Eds.; Executive Printing Services: Clovis, CA, USA, 2001; pp. 231–239. [Google Scholar]
- El Mohandes, M.A. Nonchemical methyl bromide alternatives in dates’ processing sector. In Dates: Production, Processing, Food, and Medicinal Values, 1st ed.; Manickavasagan, A., Mohamed Essa, M., Sukumar, E., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 227–253. [Google Scholar]
- Dhouibi, M.H.; Lagha, A.; Bensalem, A.; Hammami, Y. Palm dates fumigation in Tunisia: Efficiency of phosphine and CO2 mixtures, at different temperatures, as an alternative to methyl bromide. Int. J. Agric. Innov. Res. 2015, 6, 1697–1702. [Google Scholar]
- Taylor, R.W.D. Methyl bromide—Is there any future for this noteworthy fumigant? J. Stored Prod. Res. 1994, 30, 253–260. [Google Scholar] [CrossRef]
- Yahia, E.M.; Lobo, M.G.; Kader, A.A. Harvesting and postharvest technology of dates. In Dates: Postharvest Science, Processing Technology and Health Benefits, 1st ed.; Siddiq, M., Aleid, S.M., Kader, A.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 105–135. [Google Scholar] [CrossRef]
- Jiao, S.; Sun, W.; Yang, T.; Zou, Y.; Zhu, X.; Zhao, Y. Investigation of the Feasibility of Radio Frequency Energy for Controlling Insects in Milled Rice. Food Bioprocess Technol. 2017, 10, 781–788. [Google Scholar] [CrossRef]
- Wang, S.; Tiwari, G.; Jiao, S.; Johnson, J.A.; Tang, J. Developing postharvest disinfestation treatments for legumes using radio frequency energy. Biosyst. Eng. 2010, 105, 341–349. [Google Scholar] [CrossRef]
- Jiao, S.; Johnson, J.A.; Tang, J.; Wang, S. Industrial-scale radio frequency treatments for insect control in lentils. J. Stored Prod. Res. 2012, 48, 143–148. [Google Scholar] [CrossRef]
- Wang, S.; Tang, J.; Cavalieri, R.P.; Davis, D.C. Differential heating of insects in dried nuts and fruits associated with radio frequency and microwave treatments. Trans. ASAE 2003, 46, 1175–1182. [Google Scholar] [CrossRef]
- Wang, S.; Tang, J.; Sun, T.; Mitcham, E.J.; Koral, T.; Birla, S.L. Considerations in design of commercial radio frequency treatments for postharvest pest control in in-shell walnuts. J. Food Eng. 2006, 77, 304–312. [Google Scholar] [CrossRef]
- Wang, S.; Tang, J.; Johnson, J.A.; Cavalieri, R.P. Heating uniformity and differential heating of insects in almonds associated with radio frequency energy. J. Stored Prod. Res. 2013, 55, 15–20. [Google Scholar] [CrossRef]
- Macana, R.J.; Baik, O.D. Disinfestation of insect pests in stored agricultural materials using microwave and radio frequency heating: A review. Food Rev. Int. 2018, 34, 483–510. [Google Scholar] [CrossRef]
- Jiao, S.; Luan, D.; Tang, J. Principles of Radio-Frequency and Microwave heating. In Radio-Frequency heating in food processing: Principles and applications; Awuah, G.B., Ramaswamy, H.S., Tang, J., Eds.; CRC Press: Boca Raton, USA, 2014; pp. 3–20. [Google Scholar]
- Nelson, S.O. Dielectric Properties of Agricultural Materials and Their Applications, 1st ed.; Academic Press: Amsterdam, The Netherlands, 2015; p. 292. [Google Scholar]
- Nelson, S.O.; Trabelsi, S. Factors influencing the dielectric properties of agricultural and food products. J. Microw. Power Electromagn. Energy 2012, 46, 93–107. [Google Scholar] [CrossRef]
- Marra, F.; Zhang, L.; Lyng, J.G. Radio frequency treatment of foods: Review of recent advances. J. Food Eng. 2009, 91, 497–508. [Google Scholar] [CrossRef]
- Tiwari, G.; Tang, J. Radio-Frequency heating of dry foods and agricultural commodities. In Radio-Frequency Heating in Food Processing: Principles and Applications; Awuah, G.B., Ramaswamy, H.S., Tang, J., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 281–302. [Google Scholar] [CrossRef]
- Sosa-Morales, M.E.; Villa-Rojas, R.; Rojas-Laguna, R.; López-Malo, A. RF deinfestation (pest control) of agricultural products. In Radio-Frequency Heating in Food Processing: Principles and Applications; Awuah, G.B., Ramaswamy, H.S., Tang, J., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 337–353. [Google Scholar] [CrossRef]
- Wang, S.; Ikediala, J.; Tang, J.; Hansen, J.; Mitcham, E.; Mao, R.; Swanson, B. Radio frequency treatments to control codling moth in in-shell walnuts. Postharvest Biol. Technol. 2001, 22, 29–38. [Google Scholar] [CrossRef]
- Garbati Pegna, F.; Sacchetti, P.; Canuti, V.; Trapani, S.; Bergesio, C.; Belcari, A.; Zanoni, B.; Meggiolaro, F. Radio frequency treatment for postharvest disinfestation of dates. Chem. Eng. Trans. 2015, 44, 19–24. [Google Scholar]
- Garbati Pegna, F.; Sacchetti, P.; Canuti, V.; Trapani, S.; Bergesio, C.; Belcari, A.; Zanoni, B.; Meggiolaro, F. Radio frequency irradiation treatment of dates in a single layer to control Carpophilus hemipterus. Biosyst. Eng. 2017, 155, 1–11. [Google Scholar] [CrossRef]
- Navarro, S.; Finkelman, S.; Rindner, M.; Dias, R. Emigration and control of Nitidulid beetles from dates using heat. IOBC/WPRS Bull. 2004, 27, 219–225. [Google Scholar]
- Finkelman, S.; Navarro, S.; Rindner, M.; Dias, R. Use of heat for disinfestation and control of insects in dates: Laboratory and field trials. Phytoparasitica 2006, 34, 37–48. [Google Scholar] [CrossRef]
- Rafaeli, A.; Kostukovsky, M.; Carmeli, D. Successful disinfestations of sap-beetle contaminations from organically grown dates using heat treatment: A case study. Phytoparasitica 2006, 34, 204–212. [Google Scholar] [CrossRef]
- Wang, S.; Monzon, M.; Johnson, J.; Mitcham, E.; Tang, J. Industrial-scale radio frequency treatments for insect control in walnuts. I: heating uniformity and energy efficiency. Postharvest Biol. Technol. 2007, 45, 240–246. [Google Scholar] [CrossRef]
- Turinek, M.; Bavec, F.; Repič, M.; Bavec, M.; Athanassiou, C.G.; Turinek, M.; Leitner, E.; Trematerra, P.; Trdan, S. Mortality, progeny production and preference of Sitophilus zeamais adults to wheat from integrated and alternative production systems. Acta Agric. Scand. Sect. B Plant Soil Sci. 2016, 66, 443–451. [Google Scholar]
- Andrade, J.; Estévez-Pérez, M.; Perez, G.E. Statistical comparison of the slopes of two regression lines: A tutorial. Anal. Chim. Acta 2014, 838, 1–12. [Google Scholar] [CrossRef]
- Šidák, Z. Rectangular Confidence Regions for the Means of Multivariate Normal Distributions. J. Am. Stat. Assoc. 1967, 62, 626–633. [Google Scholar] [CrossRef]
- Sheskin, D.J. Handbook of Parametric Nonparametric Statistical Procedures, 3rd ed.; Chapman and Hall/CRC: New York, NY, USA, 2004; pp. 757–780. [Google Scholar]
- Social Science Statistics. Available online: https://www.socscistatistics.com/tests/chisquare2/default2.aspx (accessed on 6 May 2019).
- Marascuilo, L.A. Large-sample multiple comparisons. Psychol. Bull. 1966, 65, 280–290. [Google Scholar] [CrossRef]
- Wagh, S.T.; Razvi, N.A. Marascuilo method of multiple comparisons (An analytical study of Caesarean section delivery). IJCMR 2016, 3, 1137–1140. [Google Scholar]
- Lurie, S. Postharvest heat treatments. Postharvest Biol. Technol. 1998, 14, 257–269. [Google Scholar] [CrossRef]
- Neven, L.G. Physiological responses of insects to heat. Postharvest Biol. Technol. 2000, 21, 103–111. [Google Scholar] [CrossRef]
- Abram, P.K.; Boivin, G.; Moiroux, J.; Brodeur, J. Behavioural effects of temperature on ectothermic animals: Unifying thermal physiology and behavioural plasticity. Biol. Rev. 2017, 92, 1859–1876. [Google Scholar] [CrossRef]
- Donahaye, E.; Navarro, S.; Rindner, M.; Dias, R. The influence of different treatments causing emigration of Nitidulid beetles. Phytoparasitica 1991, 19, 273–282. [Google Scholar] [CrossRef]
- Donahaye, E.; Navarro, S.; Rindner, M.; Dias, R. Artificial Feeding Site to Investigate Emigration of Nitidulid Beetles from Dried Fruits. J. Econ. Entomol. 1992, 85, 1990–1993. [Google Scholar] [CrossRef]
- Ben-Amor, R.; De Miguel-Gómez, M.D.; Martínez-Sánchez, A.; Aguayo, E. Effect of hot air on Deglet Noor palm quality parameters and on Ectomyelois ceratoniae. J. Stored Prod. Res. 2016, 68, 1–8. [Google Scholar] [CrossRef]
- Wang, S.; Monzon, M.; Johnson, J.A.; Mitcham, E.J.; Tang, J. Industrial-scale radio frequency treatments for insect control in walnuts II: Insect mortality and product quality. Postharvest Biol. Technol. 2007, 45, 247–253. [Google Scholar] [CrossRef]
- Johnson, J.A.; Wang, S.; Tang, J. Thermal Death Kinetics of Fifth-instar Plodia interpunctella (Lepidoptera: Pyralidae). J. Econ. Entomol. 2003, 96, 519–524. [Google Scholar] [CrossRef]
- Wang, S.; Tang, J.; Hansen, J.D. Experimental and simulation methods of insect thermal death kinetics. In Heat Treatments for Postharvest Pest Control: Theory and Practice; Tang, J., Mitcham, E., Wang, S., Lurie, S., Eds.; CAB International: Oxon, UK, 2007; pp. 105–132. [Google Scholar] [CrossRef]
- Ben-Ialli, A.; Méot, J.-M.; Bohuon, P.; Collignan, A. Survival kinetics of Ephestia kuehniella eggs during 46–75 °C heat treatment. J. Stored Prod. Res. 2009, 45, 206–211. [Google Scholar] [CrossRef]
- Hou, L.; Wu, Y.; Wang, S. Thermal Death Kinetics of Cryptolestes pusillus (Schonherr), Rhyzopertha dominica (Fabricius), and Tribolium confusum (Jacquelin du Val) Using a Heating Block System. Insects 2019, 10, 119. [Google Scholar] [CrossRef]
- Tang, J.; Ikediala, J.; Wang, S.; Hansen, J.; Cavalieri, R.; Cavalieri, R. High-temperature-short-time thermal quarantine methods. Postharvest Biol. Technol. 2000, 21, 129–145. [Google Scholar] [CrossRef]
- Wang, S.; Ikediala, J.N.; Tang, J.; Hansen, J.D. Thermal death kinetics and heating rate effects for fifth-instar Cydia pomonella (L.) (Lepidoptera: Tortricidae). J. Stored Prod. Res. 2002, 38, 441–453. [Google Scholar] [CrossRef]
- Arlene-Christina, G.; Jayas, D.; Fields, P.; Jian, F.; White, N.; Alagusundaram, K. Movement of Cryptolestes ferrugineus out of wheat kernels and their mortalities under elevated temperatures. J. Stored Prod. Res. 2014, 59, 292–298. [Google Scholar] [CrossRef]
- Hou, L.; Ling, B.; Wang, S. Development of thermal treatment protocol for disinfesting chestnuts using radio frequency energy. Postharvest Biol. Technol. 2014, 98, 65–71. [Google Scholar] [CrossRef]
- Mitcham, E.; Veltman, R.; Feng, X.; De Castro, E.; Johnson, J.; Simpson, T.; Biasi, W.; Wang, S.; Tang, J. Application of radio frequency treatments to control insects in in-shell walnuts. Postharvest Biol. Technol. 2004, 33, 93–100. [Google Scholar] [CrossRef]
- Al-Azawi, A.F.; El-Haidari, H.S.; Aziz, F.M.; Murad, A.K.; Al-Saud, H.M. The effect of high temperatures on the dried fruit beetle Carpophilus hemipterus L., a pest of stored dates in Iraq. Date Palm J. 1984, 3, 327–336. [Google Scholar]
- Nelson, S.O.; Rhine, L.E.S.A.J.J. Factors Influencing Effectiveness of Radio-Frequency Electric Fields for Stored-Grain Insect Control. Trans. ASAE 1966, 9, 809–817. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Experiment Name | Irradiation Voltage (V) | Exposure Time (s) | Replicates (n) | Dates (n) | Insects (n) |
|---|---|---|---|---|---|
| RF0 | 0 | 600 | 6 | 90 | 324 |
| 5000 RF-180s | RF 5000 V | 180 | 2 | 30 | 94 |
| 5000 RF-300s | RF 5000 V | 300 | 2 | 30 | 102 |
| 3500 RF-210s | RF 3500 V | 210 | 3 | 45 | 82 |
| 3000 RF-330s | RF 3000 V | 330 | 6 | 90 | 253 |
| 3000 RF-390s | RF 3000 V | 390 | 6 | 99 | 360 |
| 2500 RF-450s | RF 2500 V | 450 | 6 | 90 | 299 |
| 2500 RF-600s | RF 2500 V | 600 | 6 | 84 | 335 |
| Experiment | Model | R² | SEE | Parameters ± SE | t | Slope (ti) |
|---|---|---|---|---|---|---|
| 5000 RF-180s | β0 | 0.94 | 2.03 | 18.087 ± 0.6156 | 29.378 | 0.217 a |
| (n = 126) | β1 | 0.2179 ± 0.0149 | 14.608 | |||
| β2 | −0.0003 ± 0.0001 | −4.345 | ||||
| 5000 RF-300s | β0 | 0.92 | 2.88 | 14.8638 ± 0.6436 | 23.095 | 0.166 a |
| (n = 210) | β1 | 0.1631 ± 0.0096 | 17.038 | |||
| β2 | −0.0002 ± 0.0000 | −5.347 | ||||
| 3500 RF-210s | β0 | 0.87 | 2.62 | 17.2123 ± 0.5488 | 31.363 | 0.148 a |
| (n = 252) | β1 | 0.1478 ± 0.0115 | 12.867 | |||
| β2 | −0.0002 ± 0.0001 | −3.066 | ||||
| 3000 RF-330s | β0 | 0.92 | 2.04 | 19.6108 ± 0.2372 | 82.672 | 0.118 b |
| (n = 741) | β1 | 0.1184 ± 0.0032 | 36.809 | |||
| β2 | −0.0001 ± 0.0000 | −14.888 | ||||
| 3000 RF-390s | β0 | 0.93 | 1.99 | 20.5970 ± 0.2310 | 89.178 | 0.100 b,c |
| (n = 759) | β1 | 0.1005 ± 0.0027 | 37.759 | |||
| β2 | −0.0001 ± 0.0000 | −14.925 | ||||
| 2500 RF-450s | β0 | 0.92 | 1.92 | 21.2550 ± 0.2410 | 88.186 | 0.087 c |
| (n = 630) | β1 | 0.0868 ± 0.0024 | 35.911 | |||
| β2 | −0.0001 ± 0.0000 | −15.663 | ||||
| 2500 RF-600s | β0 | 0.93 | 1.95 | 22.8967 ± 0.1709 | 133.956 | 0.081 c |
| (n = 1260) | β1 | 0.0814 ± 0.0013 | 62.921 | |||
| β2 | −0.0001 ± 0.0000 | −33.752 |
| Experiment | N | Temperature °C (Mean ± SD) 1 | |
|---|---|---|---|
| 5000 RF-180 | 7 | 46.67 ± 2.77 | a,b |
| 5000 RF-300 | 7 | 49.83 ± 4.72 | a |
| 3500 RF-210 | 10 | 41.31 ± 3.77 | b |
| 3000 RF-330 | 23 | 44.83 ± 2.69 | b |
| 3000 RF-390 | 22 | 45.46 ± 2.10 | a,b |
| 2500 RF-450 | 16 | 44.68 ± 1.84 | b |
| 2500 RF-600 | 23 | 47.94 ± 3.20 | a |
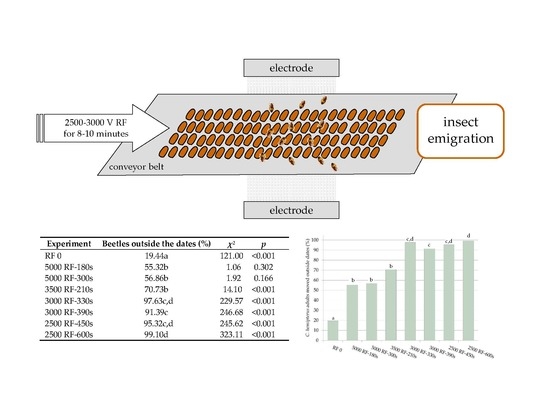
| Experiment | Beetles Outside the Dates (%) | χ2 | p |
|---|---|---|---|
| RF 0 | 19.44 a | 121.00 | <0.001 |
| 5000 RF-180 s | 55.32 b | 1.06 | 0.302 |
| 5000 RF-300 s | 56.86 b | 1.92 | 0.166 |
| 3500 RF-210 s | 70.73 b | 14.10 | <0.001 |
| 3000 RF-330 s | 97.63 c,d | 229.57 | <0.001 |
| 3000 RF-390 s | 91.39 c | 246.68 | <0.001 |
| 2500 RF-450 s | 95.32 c,d | 245.62 | <0.001 |
| 2500 RF-600 s | 99.10 d | 323.11 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosi, M.C.; Garbati Pegna, F.; Nencioni, A.; Guidi, R.; Bicego, M.; Belcari, A.; Sacchetti, P. Emigration Effects Induced by Radio Frequency Treatment to Dates Infested by Carpophilus hemipterus. Insects 2019, 10, 273. https://doi.org/10.3390/insects10090273
Rosi MC, Garbati Pegna F, Nencioni A, Guidi R, Bicego M, Belcari A, Sacchetti P. Emigration Effects Induced by Radio Frequency Treatment to Dates Infested by Carpophilus hemipterus. Insects. 2019; 10(9):273. https://doi.org/10.3390/insects10090273
Chicago/Turabian StyleRosi, Marzia Cristiana, Francesco Garbati Pegna, Anita Nencioni, Roberto Guidi, Michele Bicego, Antonio Belcari, and Patrizia Sacchetti. 2019. "Emigration Effects Induced by Radio Frequency Treatment to Dates Infested by Carpophilus hemipterus" Insects 10, no. 9: 273. https://doi.org/10.3390/insects10090273
APA StyleRosi, M. C., Garbati Pegna, F., Nencioni, A., Guidi, R., Bicego, M., Belcari, A., & Sacchetti, P. (2019). Emigration Effects Induced by Radio Frequency Treatment to Dates Infested by Carpophilus hemipterus. Insects, 10(9), 273. https://doi.org/10.3390/insects10090273