Validating Morphometrics with DNA Barcoding to Reliably Separate Three Cryptic Species of Bombus Cresson (Hymenoptera: Apidae)

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Determining Malar and Flagellar Segment Ratios
2.3. DNA Barcoding
2.4. Statistical Analyses
3. Results
3.1. Malar Ratio Measurements & DNA Barcoding
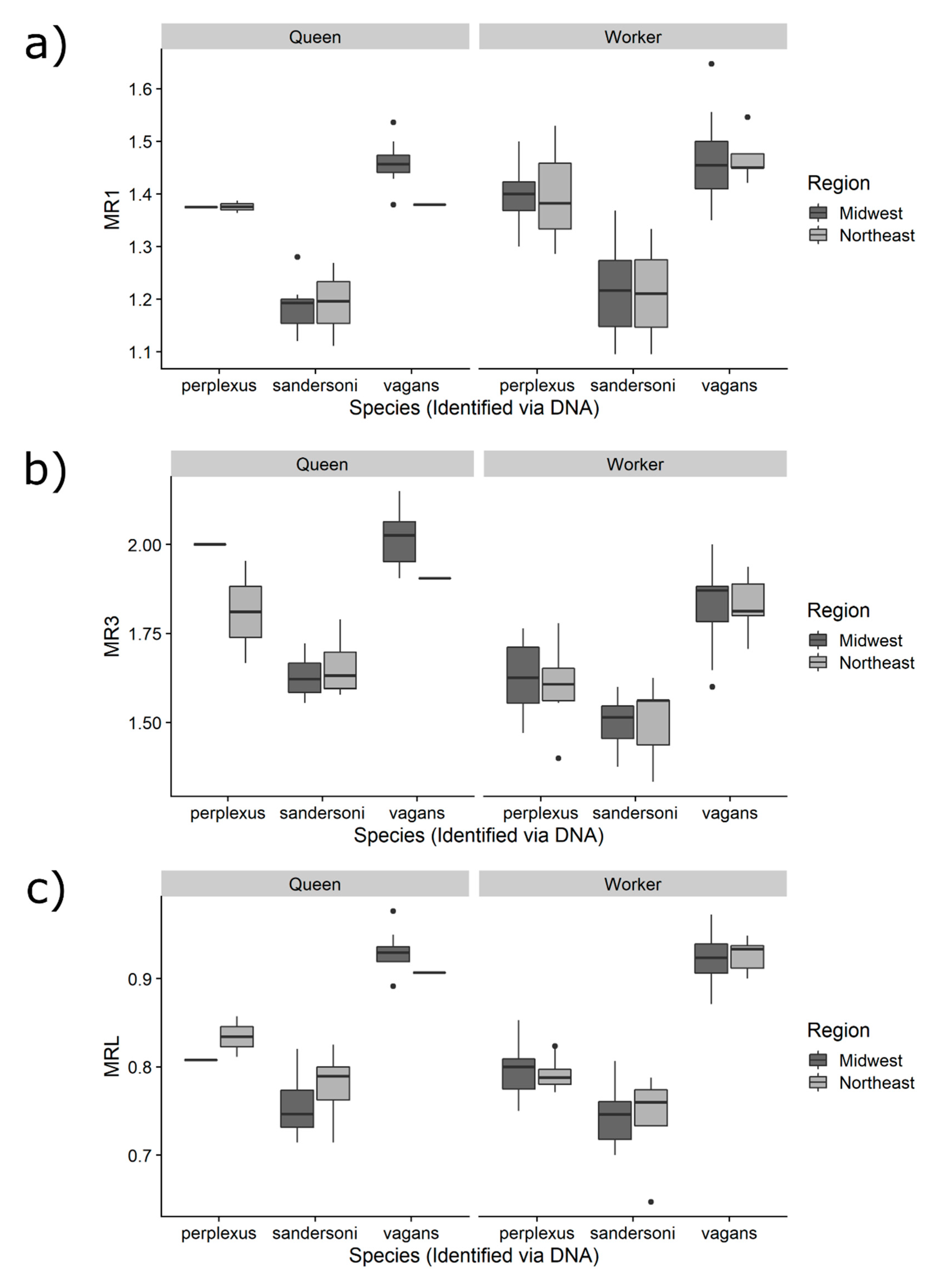
3.2. Comparing the Three Measurements among Species
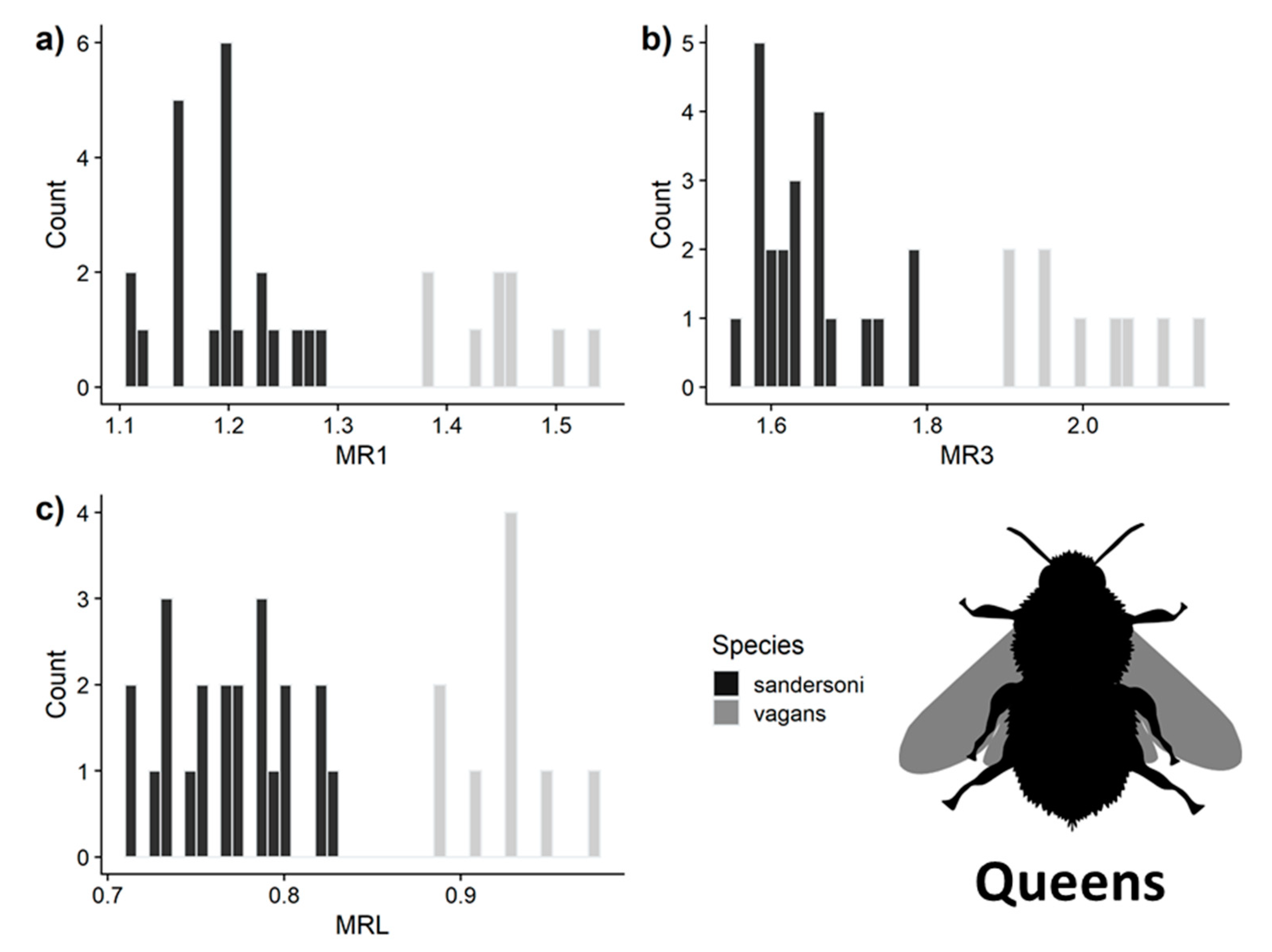
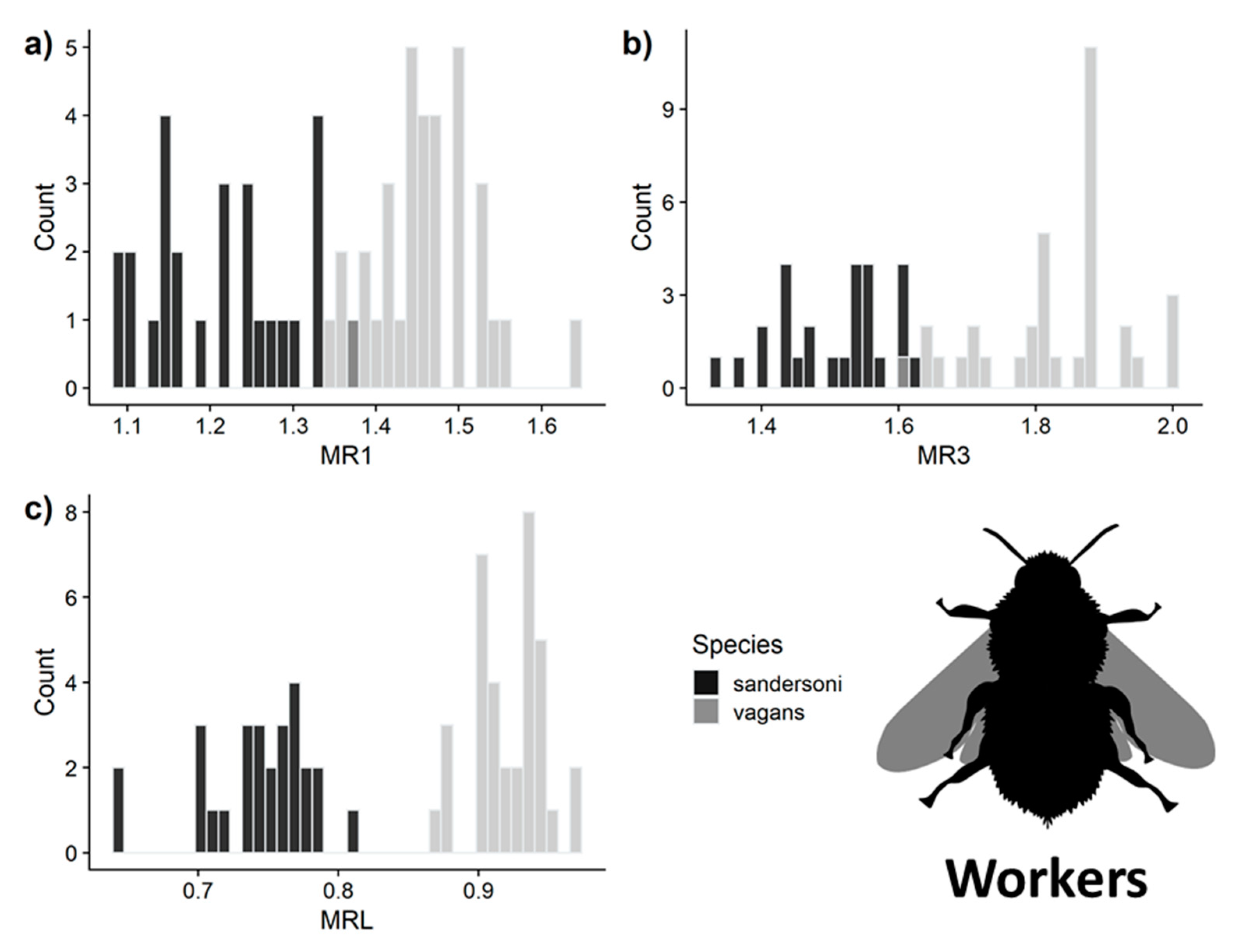
3.3. Predictive Accuracy of the Three Measurements for Separating B. vagans and B. sandersoni
3.4. Correlation within and between All Observers
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Approval
Consent to Participate
Consent for Publication
Availability of Data and Material
Code Availability
Data Deposition
Appendix A

{kind=link}
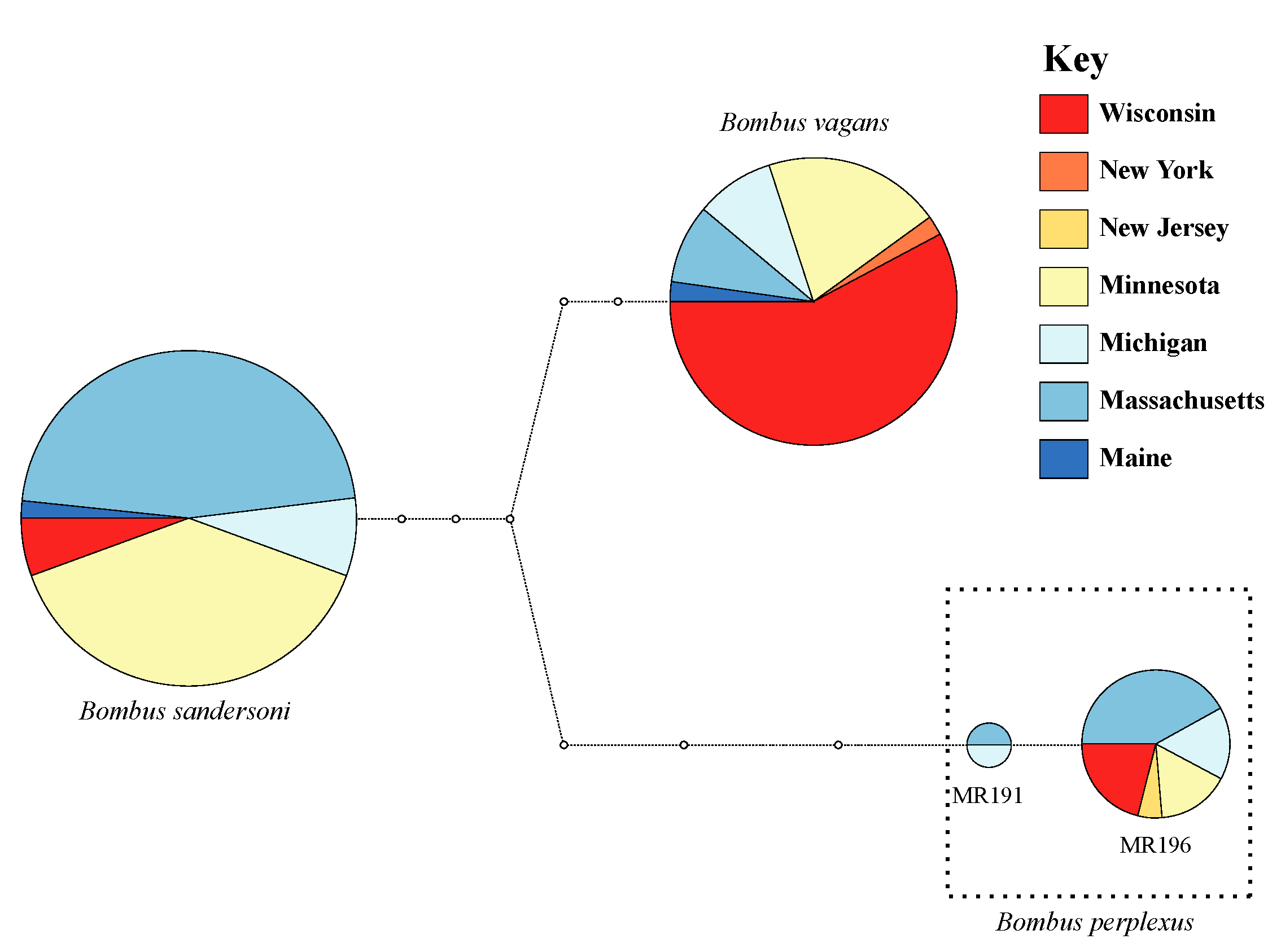{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Project ID Number | Caste | Coll. Date m/d/yr. | State Collected | Latitude | Longitude | DNA Results | GenBank Accession Numbers |
|---|---|---|---|---|---|---|---|
| MR_003 | w | 8/23/2018 | WI | 45.1385 | −88.4728 | vagans | MT951529 |
| MR_004 | w | 7/25/2018 | WI | 45.2704 | −88.3468 | vagans | MT951530 |
| MR_076 | w | 7/23/2018 | WI | 45.2002 | −88.5869 | vagans | MT951532 |
| MR_077 | w | 7/26/2018 | WI | 45.2996 | −88.3899 | vagans | MT951533 |
| MR_079 | w | 7/26/2018 | WI | 45.2996 | −88.3899 | vagans | MT951534 |
| MR_080 | w | 7/23/2018 | WI | 45.2002 | −88.5869 | vagans | MT951535 |
| MR_082 | w | 7/18/2018 | WI | 45.1682 | −88.3119 | vagans | MT951536 |
| MR_084 | w | 7/9/2018 | WI | 45.3415 | −88.4195 | vagans | MT951537 |
| MR_085 | w | 7/9/2018 | WI | 45.3198 | −88.4079 | vagans | MT951538 |
| MR_091 | w | 7/23/2018 | WI | 45.3199 | −88.4072 | vagans | MT951539 |
| MR_092 | w | 7/23/2018 | WI | 45.3199 | −88.4072 | vagans | MT951540 |
| MR_094 | w | 7/18/2018 | WI | 45.1682 | −88.3119 | vagans | MT951541 |
| MR_097 | Q | 6/8/2018 | MI | 46.5008 | −90.0185 | vagans | MT951542 |
| MR_102 | w | 6/26/2014 | NY | 43.7356 | −73.8508 | vagans | MT951543 |
| MR_105 | w | 8/15/2018 | MI | 46.5365 | −89.0134 | vagans | MT951544 |
| MR_118 | Q | 7/25/2018 | WI | 45.2704 | −88.3468 | vagans | MT991562 |
| MR_119 | Q | 8/30/2018 | WI | 45.9942 | −88.4572 | vagans | MT991563 |
| MR_121 | w | 8/14/2018 | MN | 47.7686 | −90.8927 | vagans | MT951545 |
| MR_124 | w | 7/25/2018 | WI | 45.2704 | −88.3468 | vagans | MT951546 |
| MR_127 | w | 8/15/2018 | MI | 46.5759 | −88.8877 | vagans | MT951547 |
| MR_133 | w | 8/14/2018 | MN | 47.7856 | −90.8823 | vagans | MT951548 |
| MR_134 | w | 8/14/2018 | MN | 47.7856 | −90.8823 | vagans | MT951549 |
| MR_136 | w | 8/14/2018 | MN | 47.7856 | −90.8823 | vagans | MT951550 |
| MR_144 | w | 7/19/2018 | MA | 42.6806 | −72.1117 | vagans | MT951551 |
| MR_146 | w | 7/18/2018 | MA | 42.6614 | −72.1083 | vagans | MT951552 |
| MR_151 | w | 7/10/2018 | MN | 47.7697 | −90.3107 | vagans | MT951553 |
| MR_154 | w | 7/2/2018 | MN | 47.4935 | −91.9518 | vagans | MT951554 |
| MR_160 | w | 8/14/2018 | MN | 47.7686 | −90.8927 | vagans | MT951555 |
| MR_165 | w | 8/14/2018 | MN | 47.7686 | −90.8927 | vagans | MT951556 |
| MR_167 | Q | 7/23/2018 | WI | 45.2719 | −88.6892 | vagans | MT991564 |
| MR_168 | w | 7/18/2018 | WI | 45.1682 | −88.3119 | vagans | MT951557 |
| MR_180 | w | 7/11/2014 | MA | 42.5312 | −72.3220 | vagans | MT951558 |
| MR_184 | w | 8/22/2018 | WI | 45.3215 | −88.4050 | vagans | MT951559 |
| MR_185 | Q | 5/15/2018 | WI | 45.3215 | −88.4050 | vagans | MT951560 |
| MR_186 | w | 8/8/2018 | WI | 46.0250 | −88.8744 | vagans | MT951561 |
| MR_187 | w | 8/8/2018 | WI | 46.0250 | −88.8744 | vagans | MT951562 |
| MR_199 | Q | 7/10/2018 | WI | 45.1690 | −88.3338 | vagans | MT991565 |
| MR_200 | Q | 7/10/2018 | WI | 45.1690 | −88.3338 | vagans | MT991567 |
| MR_204 | w | 8/7/2018 | WI | 45.8141 | −88.6540 | vagans | MT951563 |
| MR_207 | w | 7/19/2018 | WI | 45.2951 | −88.5143 | vagans | MT951564 |
| MR_212 | w | 8/21/2018 | MI | 46.3424 | −89.4668 | vagans | MT951565 |
| MR_215 | Q | 7/10/2018 | WI | 45.1690 | −88.3338 | vagans | MT951566 |
| MR_219 | w | 7/10/2015 | MA | 42.5225 | −72.3233 | vagans | MT951567 |
| MR_220 | Q | 5/25/2014 | ME | 43.8978 | −69.7676 | vagans | MT991566 |
| MR_001 | w | 7/11/2018 | MA | 42.3481 | −72.2324 | sandersoni | MT951476 |
| MR_011 | w | 7/11/2018 | MA | 42.3481 | −72.2324 | sandersoni | MT951477 |
| MR_013 | w | 7/11/2018 | MA | 42.3466 | −72.2272 | sandersoni | MT951478 |
| MR_015 | w | 7/11/2018 | MA | 42.3446 | −72.2275 | sandersoni | MT951479 |
| MR_017 | w | 7/11/2018 | MA | 42.3481 | −72.2324 | sandersoni | MT951480 |
| MR_040 | w | 6/5/2018 | MN | 47.8405 | −90.7564 | sandersoni | MT951483 |
| MR_042 | w | 6/8/2018 | MN | 47.5978 | −90.8237 | sandersoni | MT951484 |
| MR_043 | Q | 6/5/2018 | MN | 47.7689 | −90.8913 | sandersoni | MT951485 |
| MR_047 | w | 7/9/2018 | MN | 47.5977 | −90.8242 | sandersoni | MT951486 |
| MR_048 | w | 7/10/2018 | MN | 47.7697 | −90.3107 | sandersoni | MT951487 |
| MR_052 | w | 7/9/2018 | MN | 47.5977 | −90.8242 | sandersoni | MT951488 |
| MR_053 | w | 7/2/2018 | MN | 47.4935 | −91.9518 | sandersoni | MT951489 |
| MR_055 | w | 7/9/2018 | MN | 47.5977 | −90.8242 | sandersoni | MT951490 |
| MR_057 | w | 7/9/2018 | MN | 47.5977 | −90.8242 | sandersoni | MT951491 |
| MR_058 | w | 7/9/2018 | MN | 47.5977 | −90.8242 | sandersoni | MT951492 |
| MR_060 | w | 7/10/2018 | MN | 47.7697 | −90.3107 | sandersoni | MT951493 |
| MR_061 | w | 7/10/2018 | MN | 47.7697 | −90.3107 | sandersoni | MT951494 |
| MR_062 | w | 7/10/2018 | MN | 47.7697 | −90.3107 | sandersoni | MT951495 |
| MR_065 | w | 7/10/2018 | MN | 47.7697 | −90.3107 | sandersoni | MT951496 |
| MR_067 | Q | 8/13/2018 | MN | 47.5977 | −90.8242 | sandersoni | MT951497 |
| MR_087 | w | 7/17/2014 | MA | 42.5020 | −72.3690 | sandersoni | MT951499 |
| MR_088 | Q | 5/6/2014 | MA | 42.4310 | −72.2500 | sandersoni | MT951500 |
| MR_095 | Q | 7/23/2018 | MI | 46.2213 | −86.6676 | sandersoni | MT951501 |
| MR_099 | Q | 5/23/2018 | MN | 48.0534 | −90.0562 | sandersoni | MT951503 |
| MR_103 | w | 6/9/2010 | ME | 44.3000 | −68.3500 | sandersoni | MT951504 |
| MR_108 | Q | 5/5/2014 | MA | 42.4182 | −72.2535 | sandersoni | MT951505 |
| MR_109 | Q | 5/5/2014 | MA | 42.4187 | −72.2439 | sandersoni | MT951506 |
| MR_113 | Q | 5/5/2014 | MA | 42.4312 | −72.2501 | sandersoni | MT951507 |
| MR_139 | Q | 6/12/2018 | MA | 42.3426 | −72.2357 | sandersoni | MT951508 |
| MR_142 | Q | 7/11/2018 | MA | 42.3504 | −72.2274 | sandersoni | MT951509 |
| MR_143 | w | 7/18/2018 | MA | 42.6590 | −72.1068 | sandersoni | MT951510 |
| MR_178 | Q | 5/5/2014 | MA | 42.4141 | −72.2546 | sandersoni | MT951511 |
| MR_179 | Q | 5/5/2014 | MA | 42.4141 | −72.2546 | sandersoni | MT951512 |
| MR_181 | w | 7/9/2014 | MA | 42.4407 | −72.2490 | sandersoni | MT951513 |
| MR_214 | w | 6/20/2018 | MI | 46.3424 | −89.4668 | sandersoni | MT951514 |
| MR_216 | Q | 5/6/2015 | MA | 42.4187 | −72.2439 | sandersoni | MT951515 |
| MR_217 | w | 7/6/2015 | MA | 42.4188 | −72.2438 | sandersoni | MT951516 |
| MR_218 | w | 7/16/2015 | MA | 42.5225 | −72.3233 | sandersoni | MT951517 |
| MR_221 | Q | 5/3/2015 | MA | 42.5005 | −72.2691 | sandersoni | MT951518 |
| MR_224 | w | 6/20/2018 | MI | 46.3424 | −89.4668 | sandersoni | MT951519 |
| MR_225 | w | 6/6/2017 | MN | 47.7964 | −90.9315 | sandersoni | MT951520 |
| MR_229 | Q | 5/2/2015 | MA | 42.5312 | −72.3219 | sandersoni | MT951521 |
| MR_231 | w | 6/6/2017 | WI | 47.7660 | −88.9701 | sandersoni | MT951522 |
| MR_232 | w | 6/5/2017 | WI | 45.7384 | −88.5829 | sandersoni | MT951523 |
| MR_235 | Q | 5/2/2015 | MA | 42.5022 | −72.3697 | sandersoni | MT951524 |
| MR_237 | Q | 6/2/2018 | WI | 45.1311 | −88.3738 | sandersoni | MT991568 |
| MR_238 | Q | 6/6/2017 | MN | 47.2935 | −91.9503 | sandersoni | MT951525 |
| MR_239 | Q | 6/6/2017 | MN | 47.2935 | −91.9503 | sandersoni | MT951526 |
| MR_241 | Q | 6/6/2017 | MN | 47.2935 | −91.9503 | sandersoni | MT951527 |
| MR_009 | w | 6/20/2018 | WI | 45.9363 | −88.9506 | perplexus | MT951454 |
| MR_039 | w | 6/7/2017 | MI | 44.1351 | −85.9340 | perplexus | MT951455 |
| MR_050 | w | 8/23/2018 | WI | 45.1385 | −88.4728 | perplexus | MT951456 |
| MR_071 | w | 8/13/2018 | MN | 47.7856 | −90.8823 | perplexus | MT951457 |
| MR_115 | w | 5/24/2012 | MA | 42.3063 | −72.6946 | perplexus | MT951458 |
| MR_117 | w | 5/24/2012 | MA | 42.3063 | −72.6946 | perplexus | MT951459 |
| MR_130 | w | 6/20/2018 | WI | 45.9363 | −88.9506 | perplexus | MT951460 |
| MR_189 | w | 6/20/2018 | WI | 45.9363 | −88.9506 | perplexus | MT951461 |
| MR_190 | w | 8/13/2018 | MN | 47.7695 | −90.3098 | perplexus | MT951462 |
| MR_191 | w | 8/8/2018 | MI | 46.2715 | −89.4930 | perplexus | MT951463 |
| MR_192 | w | 8/8/2018 | MI | 46.2715 | −89.4930 | perplexus | MT951464 |
| MR_193 | w | 6/5/2018 | MN | 47.7689 | −90.8913 | perplexus | MT951465 |
| MR_194 | Q | 8/18/2018 | MI | 46.19618 | −89.1594 | perplexus | MT951466 |
| MR_195 | w | 8/15/2017 | MI | 46.3375 | −89.4753 | perplexus | MT951467 |
| MR_197 | w | 7/11/2018 | MA | 42.3399 | −72.2330 | perplexus | MT951469 |
| MR_198 | w | 7/16/2015 | MA | 42.4940 | −72.2706 | perplexus | MT951470 |
| MR_247 | w | 7/1/2019 | MA | 42.5824 | −72.5301 | perplexus | MT951471 |
| MR_248 | w | 7/3/2019 | MA | 42.5824 | −72.5301 | perplexus | MT951472 |
| MR_249 | w | 7/3/2019 | MA | 42.5824 | −72.5301 | perplexus | MT951473 |
| MR_250 | w | 7/3/2019 | MA | 42.5824 | −72.5301 | perplexus | MT951474 |
| MR_251 | Q | 7/3/2019 | MA | 42.5824 | −72.5301 | perplexus | MT951475 |
| MR_255 | Q | 4/25/2019 | MA | 42.3928 | −72.5309 | perplexus | MT991569 |
Appendix B
Appendix B.1. Equipment Required
- A stereo microscope, preferably with a 45–50× zoom capability.
- A 10× eyepiece with a reticle for measuring with at least one axis with 100 divisions in increments of 10 is preferred but other reticles work as long as the scale is large enough to measure the malar length and width.
- Reticles can be acquired for most, but not all, microscope eyepieces. Reticles regularly require a diopter adjustment to focus the reticle properly to ensure that the image of the reticle divisions are sharp and clear. It is helpful to first perform this focus exercise on the microscope stage without a specimen. The diopter adjustment compensates for differences between the observers’ eyes so it should be re-adjusted for each user to compensate for variability among observers. Instructions for diopter adjustments can be found online for microscopes and binoculars.
- Specimens must be oriented so that the body part being measured is oriented perpendicular to the axis of observation (e.g, place the specimen in a flat, horizontal position). A specimen manipulator allows for a pinned specimen to be viewed from multiple angles while remaining in a central viewing area under the microscope while allowing the viewer to maintain relative focus while twisting or turning a specimen to show all sides. A large cork can also be used, but it requires more patience to properly orient the specimen for measurement on a flat plane.
- To make the measurements, either rotate eyepiece to orient the reticle in a horizontal or vertical position or move the specimen. Try both methods to see which works best. Make one or more measurements to confirm that they are the same value.
- It is extremely important not to change the zoom between measurements. The specimen can be moved, and the focus can be adjusted between the length and width measurements, but do not change the zoom power. Each time you zoom up or down in power, the actual magnification level may be slightly different. Changing the zoom level between measurements means that you risk making your measurements at different magnification levels and the resulting ratios will be incorrect. The power of the zoom used for making measurements is not important as long as both measurements are made at the same magnification.
- Take care to ensure that both ends of the of the body part to be measured are in focus.
- Good lighting is important in making accurate determinations. A cool white LED ring lamp works well.
- Be careful to make sure that you are not 5 or 10 divisions off when reading the reticle measurement. This is a common error.
Appendix B.2. How to Measure the MRL (Malar Length to Width)
- Two measurements must be made to determine the malar ratio (MRL), i.e., the malar length and the malar width as shown in Figure 1a. The measurements of the malar length and width are made with the reticle and recorded as the number of reticle divisions. The malar length to width ratio is calculated using the following equation.MRL = malar length/malar width
- Partial divisions are not estimated, rather the measurement is taken as the number closest to the endpoint.
- The malar length is measured as the shortest distance from the base of the eye to the edge of the cheek using the definition from Williams et.al. [23]. The malar width is defined as the measurement from the outside of the mandible condyle to the outside of the cheek condyle (Figure 1a). This is equivalent to “cheek breadth” [23] or to the “basal width of the mandible” in Mitchell [27]. See Figure 1b,c for separate views of the cheek and mandible.
- Running a pin tip along the bottom edge of the cheek (away from the eye) helps to locate the bottom end point to make the measurement.
- We suggest measuring the length of the mandible at least twice, once on the horizontal axis and again on the vertical axis, to ensure that these values are equal.
Appendix B.3. How to Measure the MR1 and MR3 (Malar Length to Flagellar Segment 1 and Malar Length to Flagellar Segment 3 Length)
- Measure the malar length as described above for MRL measurements.
- Measure the length of flagellomer-1 along the horizontal axis. Place the zero end of the reticle where the textured part of the segment begins (Figure 1d).
- Measure flagellomer-3 on the horizontal axis (Figure 1d).
- Calculations for malar length to flagellar segments 1 and 3 ratios are as followsMR1 = malar length/flagellomer − 1MR3 = malar length/flagellomer − 3
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Bayo, F.; Wyckhuys, K.A. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Simmons, B.I.; Balmford, A.; Bladon, A.J.; Christie, A.P.; De Palma, A.; Dicks, L.V.; Gallego-Zamorano, J.; Johnston, A.; Martin, P.A.; Purvis, A.; et al. Worldwide insect declines: An important message, but interpret with caution. Ecol. Evol. 2019, 9, 3678–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, D.L. Insect declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M.; Winfree, R. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. USA 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.H. The distribution and decline of British bumble bees (Bombus Latr.). J. Apic. Res. 1982, 21, 236–3245. [Google Scholar] [CrossRef]
- Colla, S.R.; Packer, L. Evidence for decline in eastern North American bumblebees (Hymenoptera: Apidae), with special focus on Bombus affinis Cresson. Biodiver. Conserv. 2008, 17, 1379–1391. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Williams, P.H.; Osborne, J.L. Bumblebee vulnerability and conservation worldwide. Apidologie 2009, 40, 367–387. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, M.M.; Tucker, E.M.; Mathiasson, M.E.; Rehan, S.M. Decline of bumble bees in northeastern North America, with special focus on Bombus terricola. Biol. Conserv. 2018, 217, 437–445. [Google Scholar] [CrossRef]
- Williams, P.H.; Jepsen, S. Bumblebee specialist Group report 2018. Rep., Bumblebee Spec. Group, Int. Union Conserv. Nat., Gland, Switz. 2018. Available online: https://bumblebeespecialistgroup.org/wp-content/uploads/2020/06/BBSG-Annual-Report-2018.pdf (accessed on 28 August 2020).
- Richardson, L.L.; McFarland, K.P.; Zahendra, S.; Hardy, S. Bumble bee (Bombus) distribution and diversity in Vermont, USA: A century of change. J. Insect. Conserv. 2019, 23, 45–62. [Google Scholar] [CrossRef]
- Cameron, S.A.; Sadd, B.M. Global trends in bumble bee health. Annu. Rev. Entomol. 2020, 65, 209–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winfree, R.; Bartomeus, I.; Cariveau, D.P. Native pollinators in anthropogenic habitats. Annu. Rev. Ecolo. Evol. System. 2011, 42, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.T.; Pindar, A.; Galpern, P.; Packer, L.; Potts, S.G.; Roberts, S.M.; Rasmont, P.; Schweiger, O.; Colla, S.R.; Richardson, L.L.; et al. Climate change impacts on bumblebees converge across continents. Science 2015, 349, 177–180. [Google Scholar] [CrossRef]
- Schweiger, O.; Biesmeijer, J.C.; Bommarco, R.; Hickler, T.; Hulme, P.E.; Klotz, S.; Kühn, I.; Moora, M.; Nielsen, A.; Ohlemüller, R.; et al. Multiple stressors on biotic interactions: How climate change and alien species interact to affect pollination. Biol. Rev. 2010, 85, 777–795. [Google Scholar] [CrossRef]
- Arbetman, M.P.; Gleiser, G.; Morales, C.L.; Williams, P.; Aizen, M.A. Global decline of bumblebees is phylogenetically structured and inversely related to species range size and pathogen incidence. Proc. Roy. Soc. B 2017, 284, 2017024. [Google Scholar] [CrossRef] [Green Version]
- Soroye, P.; Newbold, T.; Kerr, J. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef]
- Cresson, E.T.; Edwards, W.H. List of the North American species of Bombus and Aporthus. Proc. Entomol. Soc. Philad. 1863, 2, 69. Available online: https://digitalcommons.usu.edu/bee_lab_co (accessed on 19 September 2020).
- Milliron, H.E. Revised classification of the bumble bees—A synopsis (Hymenoptera: Apidae). J. Kans. Entomol. Soc. 1961, 34, 49–61. [Google Scholar]
- Williams, P.H.; An, J.; Brown, M.J.F.; Carolan, J.C.; Goulson, D.; Huang, J.; Ito, M. Cryptic bumblebee species: Consequences for conservation and the trade in greenhouse pollinators. PLoS ONE 2012, 7, e32992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.H.; Thorp, R.; Richardson, L.; Colla, S. Bumble Bees of North America; University Press: Princeton, NJ, USA, 2014. [Google Scholar]
- Ellis, J.S.; Knight, M.E.; Goulson, D. Delineating species for conservation using mitochondrial sequence data: The taxonomic status of two problematic Bombus species (Hymenoptera: Apidae). J. Insect. Conserv. 2005, 9, 75–83. [Google Scholar] [CrossRef]
- Franklin, H.J. The Bombidae of the New World. Trans. Am. Entomol. Soc. 1912, 38, 177–486. [Google Scholar]
- Plath, O.E. Bumble Bees and Their Ways; MacMillan Co.: New York, NY, USA, 1934. [Google Scholar]
- Mitchell, T.B. Bees of the Eastern United States; The North Carolina Agricultural Experiment Station, The National Science Foundation: Raleigh, NC, USA, 1962; Volume II. [Google Scholar]
- Plowright, R.C.; Pallett, M.J. A morphometric study of the taxonomic status of Bombus sandersoni (Hymenoptera: Apidae). Can. Entomol. 1978, 110, 647–654. [Google Scholar] [CrossRef]
- Williams, P.H. The distribution of bumblebee colour patterns worldwide: Possible significance for thermoregulation, crypsis, and warning mimicry. Biol. J. Linn. Soc. 2007, 92, 97–118. [Google Scholar] [CrossRef] [Green Version]
- Carolan, J.C.; Murray, T.E.; Fitzpatrick, U.; Crossley, J.; Schmidt, H.; Cederberg, B.; McNally, L.; Paxton, R.J.; Williams, P.H.; Brown, M.J. Colour patterns do not diagnose species: Quantitative evaluation of a DNA barcoded cryptic bumblebee complex. PLoS ONE 2012, 7, e29251. [Google Scholar] [CrossRef] [Green Version]
- Richards, O.W. The subgeneric divisions of the genus Bombus Latreille (Hymenoptera: Apidae). Bull. Brit. Mus. (Nat. Hist.) 1968, 22, 209–276. [Google Scholar] [CrossRef]
- Waters, J.O.E.; Darvill, B.E.N.; Lye, G.C.; Goulson, D. Niche differentiation of a cryptic bumblebee complex in the Western Isles of Scotland. Insect Conserv. Div. 2011, 4, 46–52. [Google Scholar] [CrossRef]
- Frison, T.H. Additional descriptions, synonymy and records of North American bumblebees (Hymenoptera: Bremidae). Trans. Am. Ent. Coc. 1929, 55, 103–118. [Google Scholar] [CrossRef]
- Krombein, K.V.; Burks, B.D. Hymenoptera of America North of Mexico. Synoptic Catalog (Agriculture Monograph No. 2); Snd Supplement; United States Government Printing Office: Washington, DC, USA, 1967. [Google Scholar]
- Plowright, R.C.; Jay, S.C. Caste differentiation in bumblebees (Bombus Latr.: Hym.) I.—The determination of female size. Insect Soc. 1968, 15, 171–192. [Google Scholar] [CrossRef]
- Couvillon, M.J.; Jandt, J.M.; Duong, N.H.; Dornhaus, A. Ontogeny of worker body size distribution in bumble bee (Bombus impatiens) colonies. Ecol. Entomol. 2010, 35, 424–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndungu, N.N.; Nkoba, K.; Sole, C.L.; Pirk, C.W.; Abdullahi, A.Y.; Raina, S.K.; Masiga, D.K. Resolving taxonomic ambiguity and cryptic speciation of Hypotrigona species through morphometrics and DNA barcoding. J. Apic. Res. 2018, 57, 354–363. [Google Scholar] [CrossRef]
- Ascher, J.S.; Pickering, J. Discover Life Bee Species Guide and World Checklist (Hymenoptera: Apoidea: Anthophila) Draft-54. Available online: http://www.discoverlife.org/mp/20q?guide=Apoidea_species (accessed on 19 September 2020).
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith-Freedman, C.J.; Andersen, J.C.; Grifin, B.P.; Schick, K.; Elkinton, J.S. Rise and fall of an oak gall wasp (Hymenoptera: Cynipidae) outbreak on Cape Cod, Martha’s Vineyard, and Nantucket. Environ. Entomol. 2019, 2, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.C.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.; Ieno, E.N.; Smith, G.M. Analyzing Ecological Data; Springer: New York, NY, USA, 2007. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.4.4. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 19 September 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S. Fourth Edition; Springer: New York, NY, USA, 2002. [Google Scholar]
- Kuhn, M.; Caret: Classification and Regression Training. R Package Version 6.0-84. 2019. Available online: https://CRAN.R-project.org/package=caret (accessed on 19 September 2020).
- Canty, A.; Ripley, B. boot: Bootstrap R (S-Plus) Functions; R Package Version 1.3-24. 2019. Available online: https://cran.r-project.org/web/packages/boot/ (accessed on 19 September 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 19 September 2020).
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- MacPhail, V.J.; Gibson, S.D.; Hatfield, R.; Colla, S.R. Using Bumble Bee Watch to investigate the accuracy and perception of bumble bee (Bombus spp.) identification by community scientists. PeerJ 2020, 8, e9412. [Google Scholar] [CrossRef]
- Suzuki-Ohno, Y.; Yokoyama, J.; Nakashizuka, T.; Kawata, M. Utilization of photographs taken by citizens for estimating bumblebee distributions. Sci. Rep. 2017, 7, 11215. [Google Scholar] [CrossRef]
- IUCN The IUCN Red List of Threatened Species. Version 2017-2. Available online: http://www.iucnredlist.org (accessed on 28 August 2020).
- Colla, S.R.; Gadallah, F.; Richardson, L.; Wagner, D.; Gall, L. Assessing declines of North American bumble bees (Bombus spp.) using museum specimens. Biodivers. Conserv. 2012, 21, 3585–3595. [Google Scholar] [CrossRef]
- Colla, S.R. Status, threats, and conservation recommendations for wild bumble bees (Bombus spp.) in Ontario, Canada: A review for policymakers and practitioners. Nat. Areas. J. 2016, 36, 412–426. [Google Scholar] [CrossRef]
- Grixti, J.C.; Wong, L.T.; Cameron, S.A.; Favret, C. Decline of bumble bees (Bombus) in the North American Midwest. Biol. Conserv. 2009, 142, 7584. [Google Scholar] [CrossRef]
- Franklin, H.J. Cranberry insects in Massachusetts. Part V Insects and other animals beneficial in cranberry growing. Massa. Agric. Exp. St. Bull. 1950, 445, 55–76. [Google Scholar]
- Goldstein, P.Z.; Ascher, J.S. Taxonomic and behavioral composition of an island fauna: A survey of bees (Hymenoptera: Apoidea: Anthophila) on Martha’s Vineyard, Massachusetts. Proc. Entomolo. Soc. Wash. 2016, 118, 37–92. [Google Scholar] [CrossRef]
- Cane, J.H.; Tepedino, V.J. Causes and extent of declines among native North American invertebrate pollinators: Detection, evidence, and consequences. Conserv. Ecol. 2001, 5, 1. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carre, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Mono. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 45–353. [Google Scholar] [CrossRef]
- Williams, N.M.; Minckley, R.L.; Silveira, F.A. Variation in native bee faunas and its implications for detecting community changes. Conserv. Ecol. 2001, 5, 7. [Google Scholar] [CrossRef]
- Carvell, C.; Roy, D.B.; Smart, S.M.; Pywell, R.F.; Preston, C.D.; Goulson, D. Declines in forage availability for bumblebees at a national scale. Biol. Conserv. 2006, 132, 481–489. [Google Scholar] [CrossRef]
- Frankie, G.W.; Thorp, R.W.; Hernandez, J.; Rizzardi, M.; Ertter, B.; Pawelek, J.C.; Witt, S.L.; Schindler, M.; Coville, R.; Wojcik, V.A. Native bees are a rich natural resource in urban California gardens. Calif. Agric. 2009, 63, 113–120. [Google Scholar] [CrossRef]
- Morales, C.L.; Arbetman, M.P.; Cameron, S.A.; Aizen, M.A. Rapid ecological replacement of a native bumble bee by invasive species. Front. Ecol. Environ. 2013, 11, 529–534. [Google Scholar] [CrossRef]
- Packer, L.; Gibbs, J.; Sheffield, C.; Hanner, R. DNA barcoding and the mediocrity of morphology. Mol. Ecol. Res. 2009, 9, 42–50. [Google Scholar] [CrossRef] [PubMed]





| Response | Variable | Levels | β ± SE | df | F | p† |
|---|---|---|---|---|---|---|
| MR1 | Species | 2110 | 183.18 | <0.0001 | ||
| perplexus-sandersoni | 0.18 ± 0.02 | 110 | <0.0001 | |||
| perplexus-vagans | −0.07 ± 0.02 | 110 | 0.0009 | |||
| sandersoni-vagans | −0.25 ± 0.01 | 110 | <0.0001 | |||
| Caste | Worker | 0.02 ± 0.01 | 1110 | 2.10 | 0.15 | |
| Region | Northeast | 0.00 ± 0.01 | 1110 | 0.01 | 0.94 | |
| MR3 | Species | 2110 | 121.26 | <0.0001 | ||
| perplexus-sandersoni | 0.14 ± 0.02 | 110 | <0.0001 | |||
| perplexus-vagans | −0.20 ± 0.03 | 110 | <0.0001 | |||
| sandersoni-vagans | −0.34 ± 0.02 | 110 | <0.0001 | |||
| Caste | Worker | −0.17 ± 0.02 | 1110 | 67.99 | <0.0001 | |
| Region | Northeast | −0.00 ± 0.02 | 1110 | 0.05 | 0.83 | |
| MRL | Species | 2110 | 362.45 | <0.0001 | ||
| perplexus-sandersoni | 0.05 ± 0.01 | 110 | <0.0001 | |||
| perplexus-vagans | −0.13 ± 0.01 | 110 | <0.0001 | |||
| sandersoni-vagans | −0.18 ± 0.01 | 110 | <0.0001 | |||
| Caste | Worker | −0.02 ± 0.01 | 1110 | 9.55 | 0.003 | |
| Region | Northeast | 0.01 ± 0.01 | 1110 | 1.22 | 0.27 |
| Species | Caste | MR1 (Mean ± SE, 95% CI) | MR3(Mean ± SE, 95% CI) | MRL (Mean ± SE, 95% CI) |
|---|---|---|---|---|
| perplexus | Worker | 1.40 ± 0.02, 1.37–1.42 | 1.63 ± 0.02, 1.59–1.67 | 0.80 ± 0.01, 0.78–0.81 |
| Queen | 1.38 ± 0.02, 1.34–1.41 | 1.79 ± 0.03, 1.74–1.85 | 0.82 ± 0.01, 0.80–0.83 | |
| sandersoni | Worker | 1.21 ± 0.01, 1.19–1.24 | 1.49 ± 0.02, 1.46–1.53 | 0.75 ± 0.01, 0.74–0.76 |
| Queen | 1.19 ± 0.01, 1.17–1.22 | 1.66 ± 0.02, 1.62–1.69 | 0.77 ± 0.01, 0.75–0.78 | |
| vagans | Worker | 1.46 ± 0.01, 1.44–1.48 | 1.83 ± 0.02, 1.80–1.86 | 0.92 ± 0.01, 0.91–0.93 |
| Queen | 1.44 ± 0.02, 1.41–1.47 | 2.00 ± 0.02, 1.95–2.04 | 0.94 ± 0.01, 0.93–0.96 |
| Species | Caste | N | Mesipisternum Hair Color | Presence of Black Hairs on Scutum | Presence of Yellow Hairs on T5 | MR1 † | MR3 † | MRL † |
|---|---|---|---|---|---|---|---|---|
| perplexus | Worker | 19 | dark-light | none-few | none-few | 1.29–1.53 (1.24–1.56) | 1.40–1.78 (1.37–1.86) | 0.75–0.85 (0.73–0.85) |
| sandersoni | Worker | 27 | light | few-many | none-few-many | 1.10–1.37 (1.00–1.42) | 1.33–1.63 (1.30–1.70) | 0.65–0.81 (0.64–0.84) |
| vagans | Worker | 35 | light | many | none-few-many | 1.35-1.65 (1.30–1.62) | 1.60–2.00 (1.57–2.08) | 0.87–0.97 (0.86–0.99) |
| perplexus | Queen | 3 | dark | none-few | none | 1.36-1.39 (1.35–1.40) | 1.67–2.00 (1.54–2.20) | 0.81–0.86 (0.78–0.87) |
| sandersoni | Queen | 22 | light | few-many | none-few-many | 1.11–1.28 (1.07–1.31) | 1.56–1.79 (1.48–1.81) | 0.71–0.83 (0.68–0.85) |
| vagans | Queen | 9 | light | many | none-few-many | 1.38–1.54 (1.33–1.57) | 1.90–2.15 (1.80–2.22) | 0.89–0.98 (0.86–0.99) |
| Comparison | Measurement | R | t | df | p |
|---|---|---|---|---|---|
| Within Obs1 | MR1 | 0.95 | 33.08 | 113 | <0.001 |
| MR3 | 0.94 | 28.48 | 113 | <0.001 | |
| MRL | 0.92 | 25.50 | 113 | <0.001 | |
| Within Obs2 | MR1 | 0.92 | 25.68 | 113 | <0.001 |
| MR3 | 0.90 | 21.61 | 113 | <0.001 | |
| MRL | 0.86 | 18.12 | 113 | <0.001 | |
| Between experienced observers (Obs1 vs. Obs2) | MR1 | 0.92 | 24.91 | 113 | <0.001 |
| MR3 | 0.91 | 22.72 | 113 | <0.001 | |
| MRL | 0.93 | 26.55 | 113 | <0.001 | |
| Between experienced and inexperienced observers (Obs1 vs. Obs3) | MR1 | 0.85 | 8.69 | 29 | <0.001 |
| MR3 | 0.81 | 7.39 | 29 | <0.001 | |
| MRL | NA | NA | NA | NA | |
| Between SD and the mean (Obs1 vs. Obs2) | MR1 | 0.02 | 0.20 | 113 | 0.84 |
| MR3 | 0.11 | 1.22 | 113 | 0.23 | |
| MRL | −0.18 | −1.90 | 113 | 0.06 | |
| Between SD and the mean (Obs1 vs. Obs3) | MR1 | 0.02 | 0.12 | 29 | 0.90 |
| MR3 | −0.02 | −0.09 | 29 | 0.93 | |
| MRL | NA | NA | NA | NA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milam, J.; Johnson, D.E.; Andersen, J.C.; Fassler, A.B.; Narango, D.L.; Elkinton, J.S. Validating Morphometrics with DNA Barcoding to Reliably Separate Three Cryptic Species of Bombus Cresson (Hymenoptera: Apidae). Insects 2020, 11, 669. https://doi.org/10.3390/insects11100669
Milam J, Johnson DE, Andersen JC, Fassler AB, Narango DL, Elkinton JS. Validating Morphometrics with DNA Barcoding to Reliably Separate Three Cryptic Species of Bombus Cresson (Hymenoptera: Apidae). Insects. 2020; 11(10):669. https://doi.org/10.3390/insects11100669
Chicago/Turabian StyleMilam, Joan, Dennis E. Johnson, Jeremy C. Andersen, Aliza B. Fassler, Desiree L. Narango, and Joseph S. Elkinton. 2020. "Validating Morphometrics with DNA Barcoding to Reliably Separate Three Cryptic Species of Bombus Cresson (Hymenoptera: Apidae)" Insects 11, no. 10: 669. https://doi.org/10.3390/insects11100669
APA StyleMilam, J., Johnson, D. E., Andersen, J. C., Fassler, A. B., Narango, D. L., & Elkinton, J. S. (2020). Validating Morphometrics with DNA Barcoding to Reliably Separate Three Cryptic Species of Bombus Cresson (Hymenoptera: Apidae). Insects, 11(10), 669. https://doi.org/10.3390/insects11100669