Bounds on Absolute Gypsy Moth (Lymantria dispar dispar) (Lepidoptera: Erebidae) Population Density as Derived from Counts in Single Milk Carton Traps



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
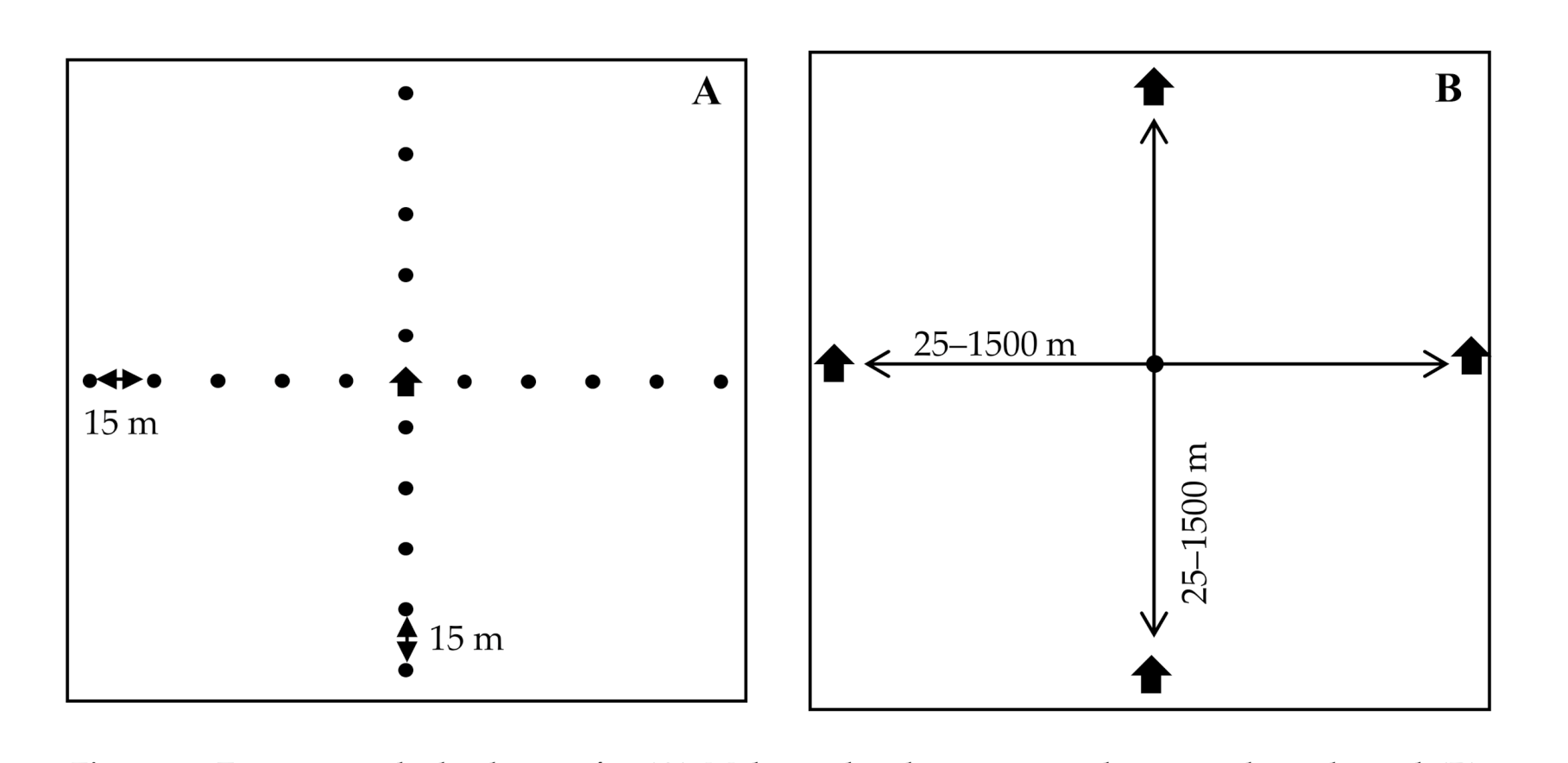
2.1. Experimental Design
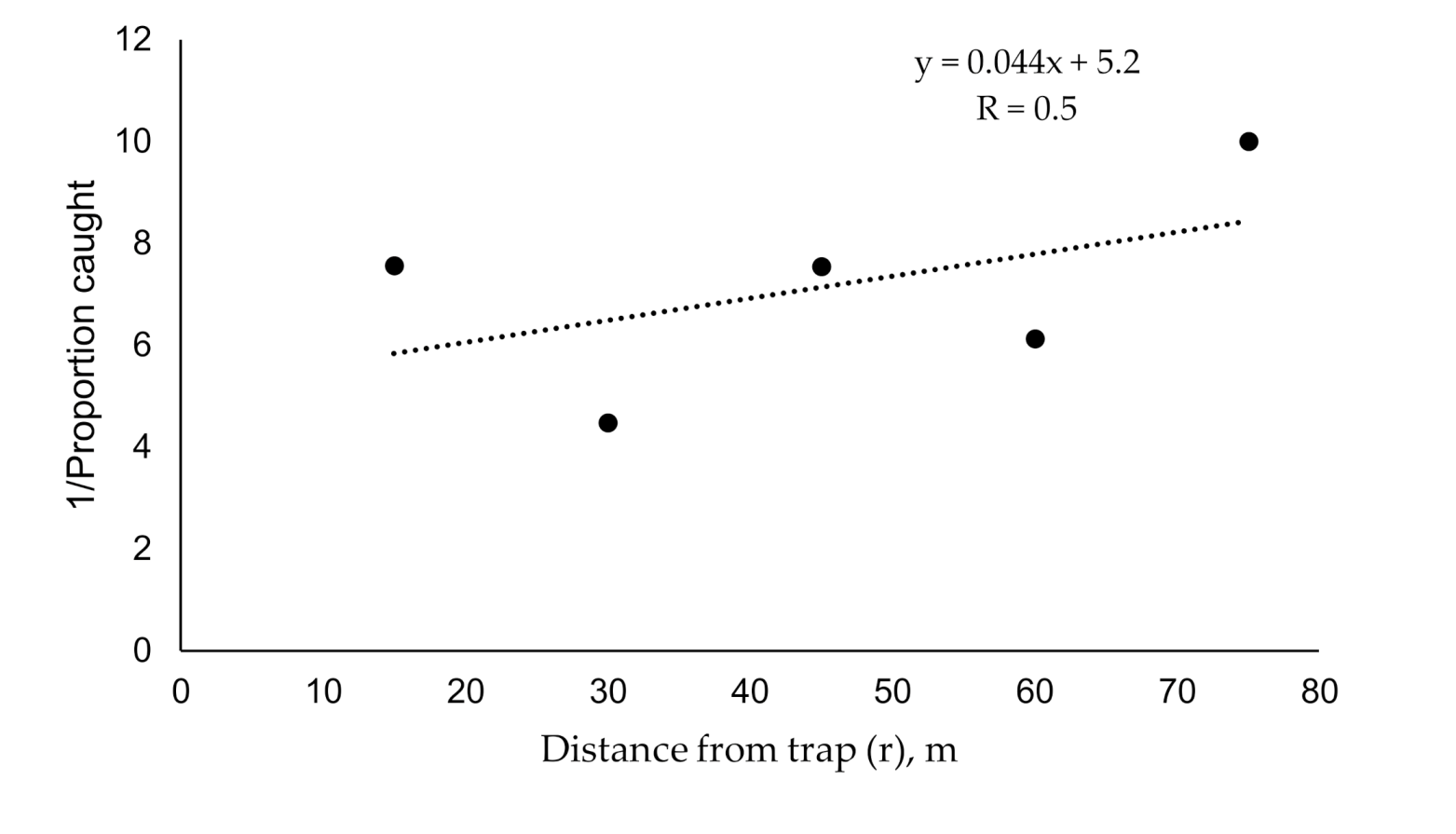
2.2. Estimating Plume Reach and Catch Probability using Existing Methods
2.3. The Novel Model and Its Derivation
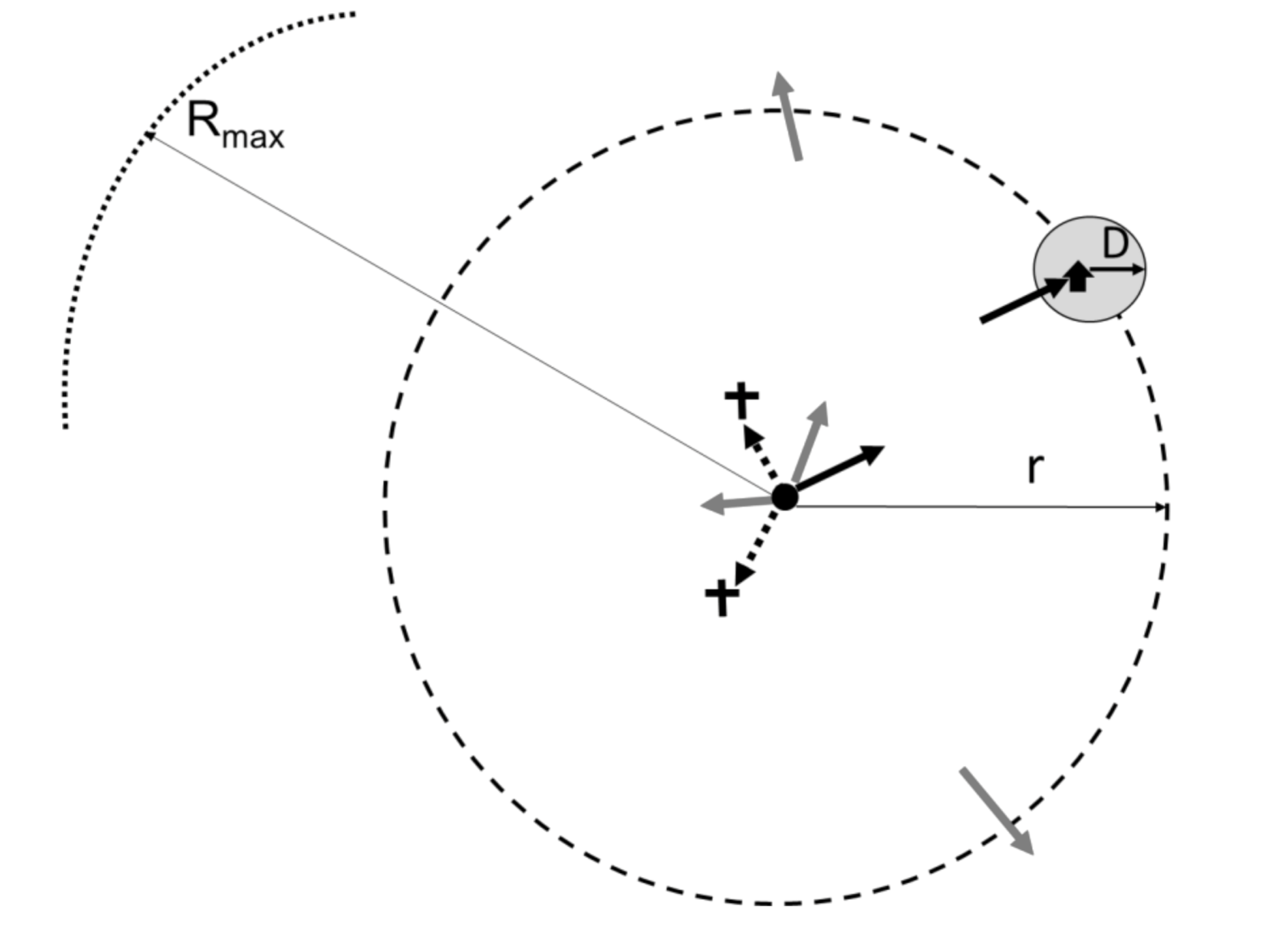
2.3.1. Preliminaries and Definitions
2.3.2. Constructing the Model
- (1)
- In the limiting case of r = 0, the male is released right next to the trap. The insect is clearly well within the plume reach, and it becomes trapped with a constant probability spTfer0, which can be measured experimentally for the given trap type.
- (2)
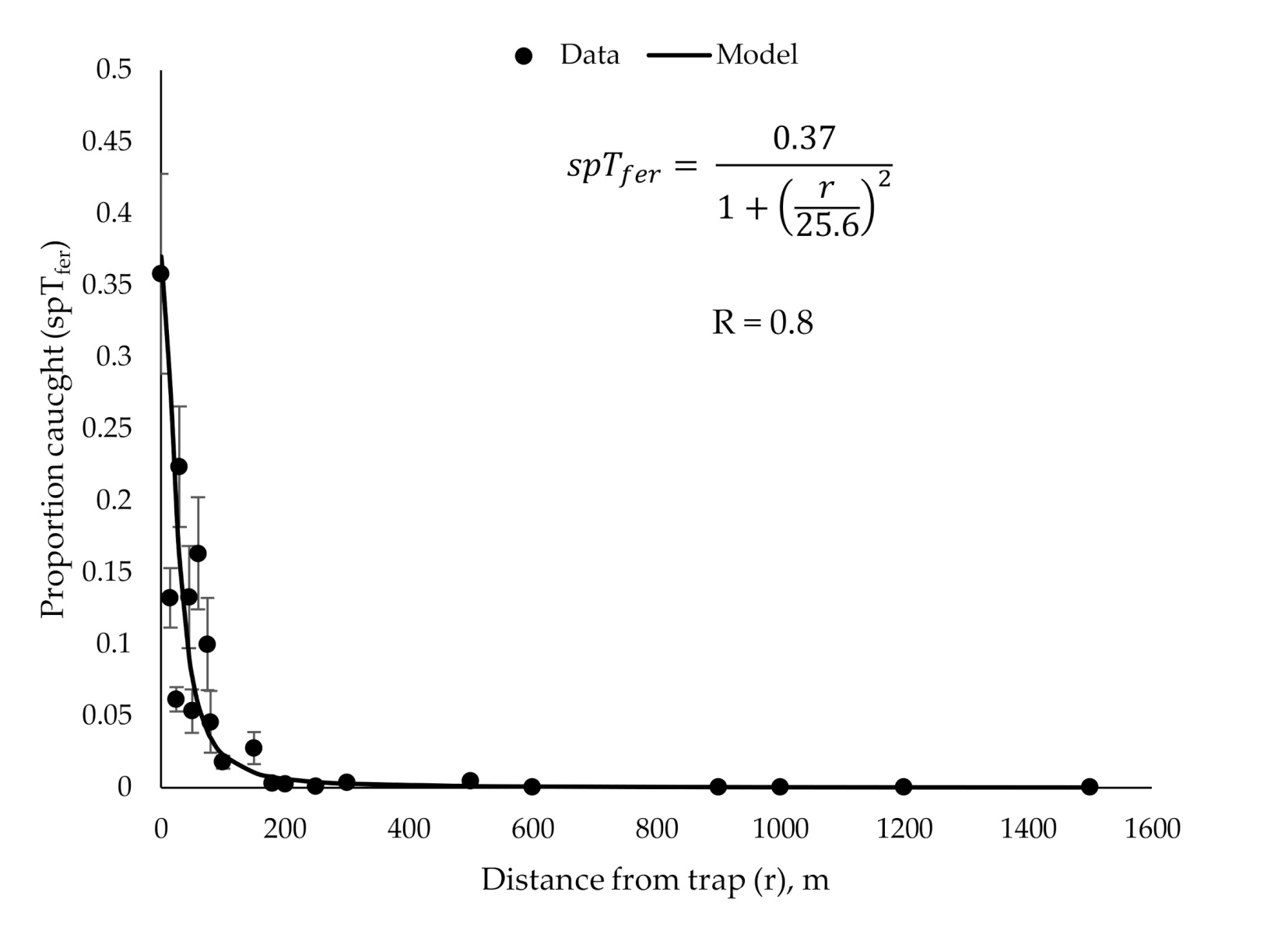
- For small, but non-zero r << D, the plume reach is nearly as strong as at r = 0, and so the over-all catch probability should not be too much smaller than spTfer(0). In this range, we expect the shape of spTfer(r) function to resemble p0 (r) of Figure 3. As the distance to the trap becomes comparable to the plume reach, r ~ D, spTfer(r) begins to decrease appreciably.
- (3)
- The most interesting and complex regime corresponds to large distances between the release point and the trap, r >> D, but still smaller than Rmax. We consider the male trajectory to be essentially 2-dimensional in this case, confined to a relatively narrow (compared to r) zone between the ground and the tree line. There are three distinct outcomes of a male trajectory at these distances (Figure 4). First, the male trajectory can enter the plume circle around the trap, and so the male becomes trapped with the probability spTfer(0). Second, the male can travel outside of the circle of radius r around the release point; we assume that for large r, its likelihood of coming back and eventually getting trapped is negligible. If these were the only two possibilities, the proportion of males trapped would be ~(), which is the ratio of the total outward insect flux 2πr to the flux through the plume reach circle 2πD. Here, flux = (number of males) × (circle perimeter). However, at large values of r, that is, at large times elapsed from the release, the males begin to die or stop the search for various reasons, that is the insect flux through the “outer” circle r is not conserved, but instead decreases with time, and, hence with the distance from the release point. As the simplest approximation, we assume that male flux falls of as 1/r. The net result is that the proportion of males trapped decreases with distance as ~ for large r (the extra “D” makes sure that the expression is dimensionless). Thus, far away from the release point, the probability of the male trapped is .
- (4)
- Finally, for r > Rmax, spTfer(r) = 0, which means males simply do not travel that far.
2.3.3. Estimating the Absolute Insect Density from Individual Trap Catch Data
3. Results
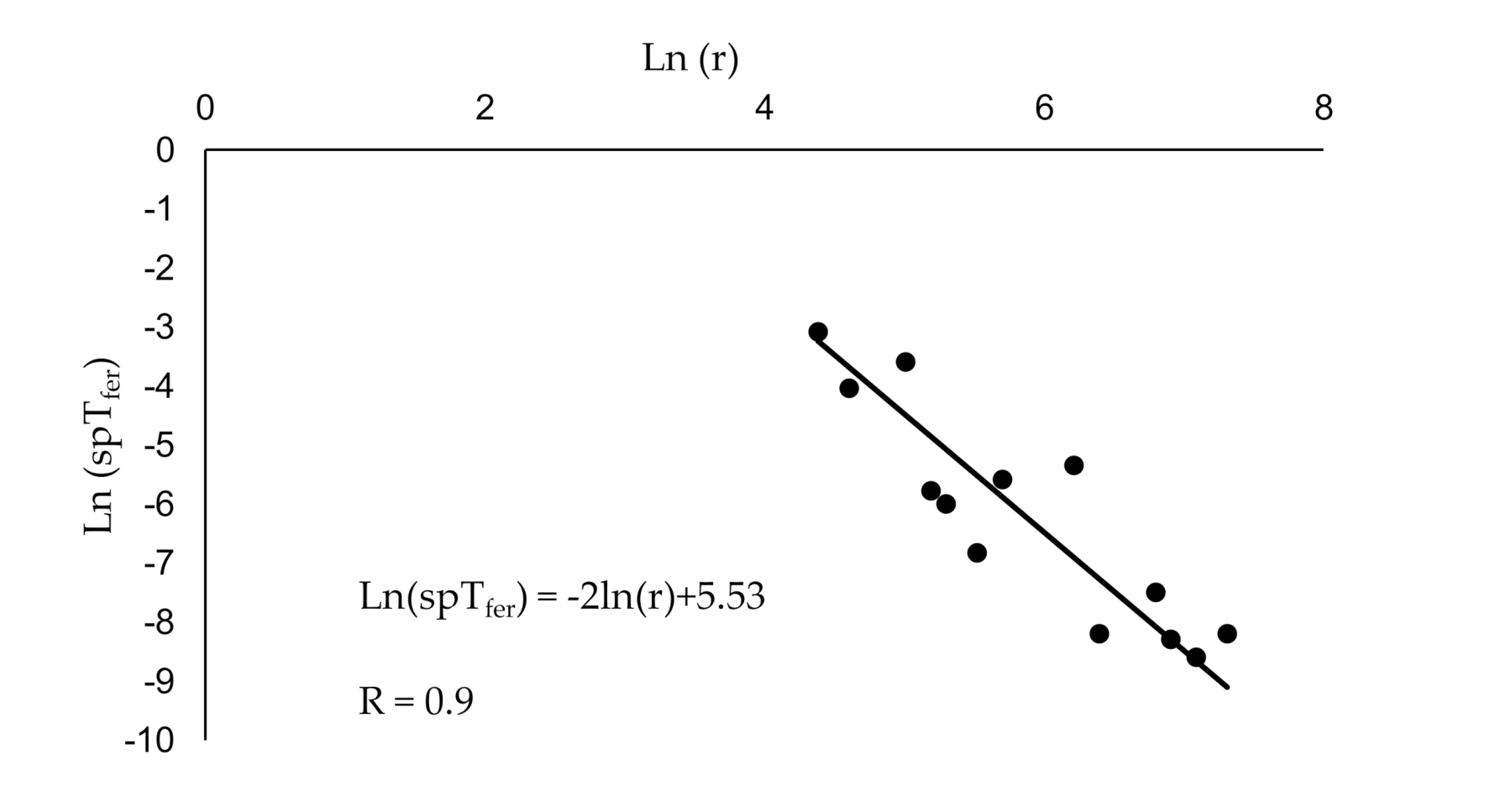
3.1. Estimating Plume Reach and Catch Probability Using Existing Methods
3.2. Validation of the New Model
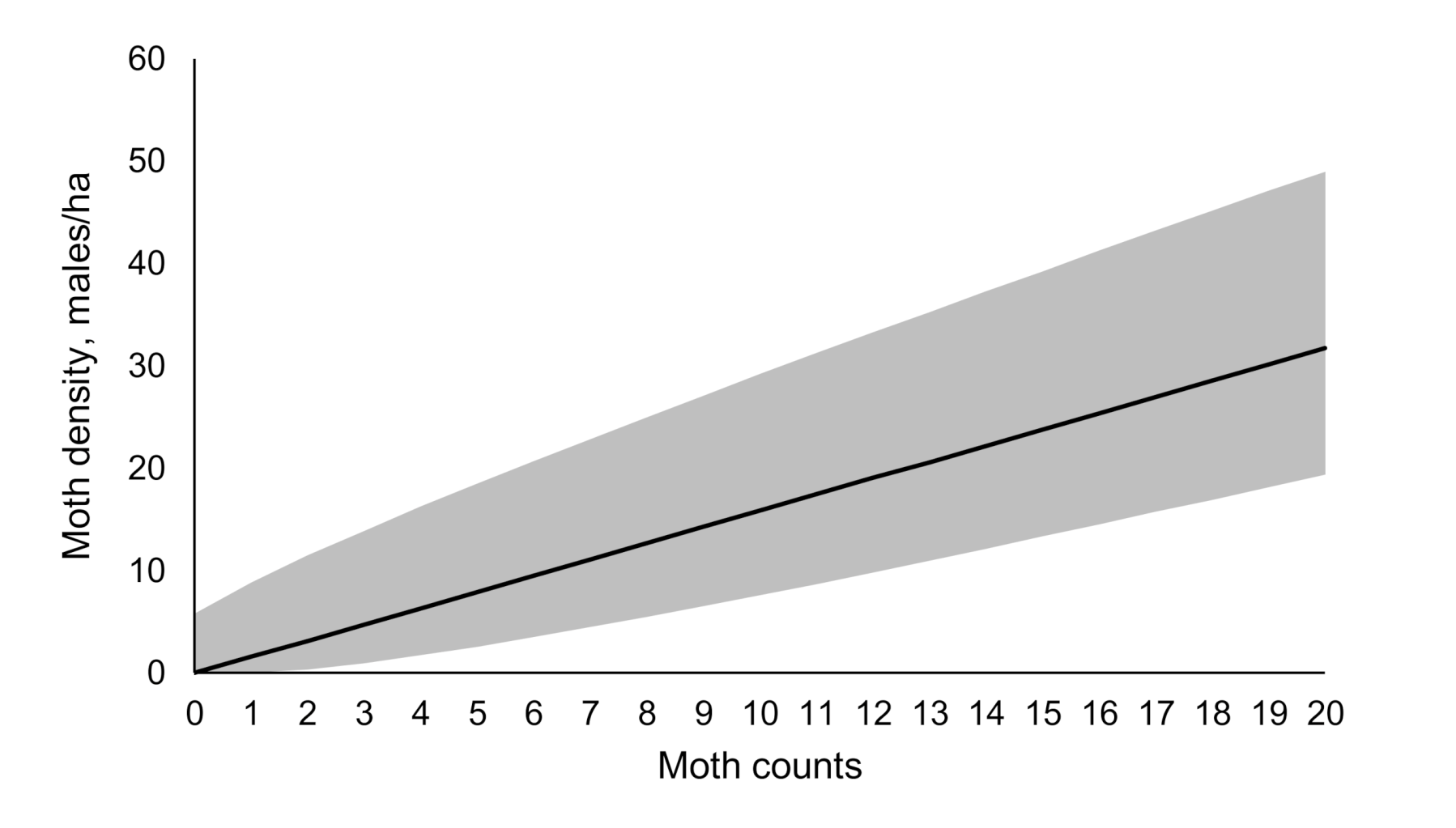
3.3. Model Application: Estimate of the Average Gypsy Moth Density from the Catch Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liebhold, A.M.; Gottschalk, K.W.; Muzika, R.-M.; Montgomery, M.E.; Young, R. Suitability of North American Tree Species to the Gypsy Moth: A Summary of Field and Laboratory Tests; NE-211; US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Madison, WI, USA, 1995.
- Gray, D.R. The gypsy moth life stage model: Landscape-wide estimates of gypsy moth establishment using a multi-generational phenology model. Ecol. Model. 2004, 176, 155–171. [Google Scholar] [CrossRef]
- Miller, J.C.; Hanson, P.E.; Dowell, R.V. Potential of gypsy moth as a pest of fruit and nut crops. Calif. Agric. 1987, 41, 10–12. [Google Scholar]
- Bigsby, K.M.; Ambrose, M.J.; Tobin, P.C.; Sills, E.O. The cost of gypsy moth sex in the city. Urban For. Urban Green. 2014, 13, 459–468. [Google Scholar] [CrossRef]
- Wirtz, R. Allergic and toxic reactions to non-stinging arthropods. Annu. Rev. Entomol. 1984, 29, 47–69. [Google Scholar] [CrossRef]
- Tobin, P.C.; Blackburn, L.M. Slow the Spread: A National Program to Manage the Gypsy Moth; General Technical Report NRS-6; USDA Forest Service: Newtown Square, PA, USA, 2007.
- Thorpe, K.W.; Ridgway, R.L.; Leonhardt, B.A. Relationship between Gypsy-Moth (Lepidoptera, Lymantriidae) Pheromone Trap Catch and Population-Density—Comparison of Traps Baited with 1 and 500 Mu-G (+)-Disparlure Lures. J. Econ. Entomol. 1993, 86, 86–92. [Google Scholar] [CrossRef]
- Carter, M.R.; Ravlin, F.W.; Mcmanus, M.L. Estimating Gypsy-Moth (Lepidoptera, Lymantriidae) egg mass density using male moths captured in pheromone-baited, milk-carton traps. Environ. Entomol. 1994, 23, 556–561. [Google Scholar] [CrossRef]
- Brown, M.W.; Rosenberger, J.L.; Cameron, E.A. Development and Evaluation of a Sampling Scheme for the Gypsy-Moth Egg Parasitoid Ooencyrtus-Kuvanae (Hymenoptera, Encyrtidae). Can. Entomol. 1981, 113, 575–584. [Google Scholar] [CrossRef]
- Abell, K.; Poland, T.M.; Cossé, A.; Bauer, L.S. Trapping techniques for emerald ash borer and its introduced parasitoids. In Biology and Control of Emerald Ash Borer; Van Driesche, R.G., Reardon, R.C., Eds.; FHTET-2014-09; U.S. Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2015; pp. 113–127. [Google Scholar]
- Elkinton, J.S.; Cardé, R.T. The Use of Pheromone Traps to Monitor the Distribution and Population Trends of the Gypsy Moth. In Management of Insect Pests with Semiochemicals; Mitchell, E.D., Ed.; Plenum: New York, NY, USA, 1981; pp. 41–55. [Google Scholar]
- Liebhold, A.M.; Tobin, P.C. Population ecology of insect invasions and their management. Ann. Rev. Entomol. 2008, 53, 387–408. [Google Scholar] [CrossRef] [Green Version]
- Sharov, A.A.; Liebhold, A.M.; Ravlin, F.W. Prediction of gypsy moth (Lepidoptera: Lymantriidae) mating success from pheromone trap counts. Environ. Entomol. 1995, 24, 1239–1244. [Google Scholar] [CrossRef]
- Tobin, P.C.; Klein, K.T.; Leonard, D.S. Gypsy moth (Lepidoptera: Lymantriidae) flight behavior and phenology based on field-deployed automated pheromone-baited traps. Environ. Entomol. 2009, 38, 1555–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byers, J.; Anderbrant, O.; Löqvist, J. Effective attraction radius. J. Chem. Ecol. 1989, 15, 749–765. [Google Scholar] [CrossRef] [PubMed]
- Byers, J. Active Space of Pheromone Plume and its Relationship to Effective Attraction Radius in Applied Models. J. Chem. Ecol. 2008, 34, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.A. Estimating insect flight densities from attractive trap catches and flight height distributions. J. Chem. Ecol. 2012, 38, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Wall, C.; Perry, J. Range of action of moth sex-attractant sources. Entomol. Exp. Appl. 1987, 44, 5–14. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Childs, R.D. Efficiency of two gypsy moth (Lepidoptera: Lymantriidae) pheromone-baited traps. Environ. Entomol. 1983, 12, 1519–1525. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Cardé, R.T. Distribution, dispersal, and apparent survival of male gypsy moths as determined by capture in pheromone-baited traps. Environ. Entomol. 1980, 9, 729. [Google Scholar] [CrossRef]
- Dufourd, C.; Weldon, C.; Anguelov, R.; Dumont, Y. Parameter identification in population models for insects using trap data. BioMath 2013, 2, 1312061. [Google Scholar] [CrossRef] [Green Version]
- Riedl, H.; Croft, B.A. A study of pheromone trap catches in relation to codling moth (Lepidoptera: Olethreutidae) damage. Can. Entomol. 1974, 106, 525–537. [Google Scholar] [CrossRef]
- Barclay, H.; Hendrichs, J. Modeling trapping of fruit flies for detection, suppression, or eradication. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 379–420. [Google Scholar]
- Gage, S.H.; Wirth, T.M.; Simmons, G.A. Predicting Regional gypsy-moth (Lymantriidae) population trends in an expanding population using pheromone trap catch and spatial-analysis. Environ. Entomol. 1990, 19, 370–377. [Google Scholar] [CrossRef]
- Taylor, R.A.J.; Mcmanus, M.L.; Pitts, C.W. The absolute eficiency of gypsy moth, Lymantria dispar (Lepidoptera, Lymantriidae), milk carton pheromone traps. Bull. Entomol. Res. 1991, 81, 111–118. [Google Scholar] [CrossRef]
- Bau, J.; Cardé, R.T. Simulation modeling to interpret the captures of moths in pheromone-baited traps used for surveillance of invasive species: The gypsy moth as a model case. J. Chem. Ecol. 2016, 42, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Östrand, F.; Anderbrant, O. From where are insects recruited? A new model to interpret catches of attractive traps. Agric. For. Entomol. 2003, 5, 163–171. [Google Scholar] [CrossRef]
- Miller, J.R.; Adams, C.G.; Weston, P.A.; Schenker, J.H. Trapping of Small Organisms Moving Randomly: Principles and Applications to Pest Monitoring and Management; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Miller, J.R. Sharpening the precision of pest management decisions: Assessing variability inherent in catch number and absolute density estimates derived from pheromone-baited traps monitoring insects moving randomly. J. Econ. Entomol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.G.; Schenker, J.H.; McGhee, P.S.; Gut, L.J.; Brunner, J.F.; Miller, J.R. Maximizing information yield from pheromone-baited monitoring traps: Estimating plume reach, trapping radius, and absolute density of Cydia pomonella (Lepidoptera: Tortricidae) in Michigan Apple. J. Econ. Entomol. 2017, 110, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, D.M.; Gut, L.J.; Miller, J.R. Estimating monitoring trap plume reach and trapping area for Drosophila suzukii (Diptera: Drosophilidae) in Michigan tart cherry. J. Econ. Entomol. 2018, 111, 1285–1289. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; Acebes-Doria, A.L.; Rice, K.B.; Short, B.D.; Adams, C.G.; Gut, L.J.; Leskey, T.C. Estimating monitoring trap plume reach and trapping area for nymphal and adult Halyomorpha halys (Hemiptera: Pentatomidae) in crop and non-crop habitats. Environ. Entomol. 2019, 48, 1104–1112. [Google Scholar] [CrossRef] [Green Version]
- Robinet, C.; Lance, D.R.; Thorpe, K.W.; Onufrieva, K.S.; Tobin, P.C.; Liebhold, A.M. Dispersion in time and space affect mating success and Allee effects in invading gypsy moth populations. J. Anim. Ecol. 2008, 77, 966–973. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Carde, R.T. Effects of Intertrap Distance and Wind Direction on the Interaction of Gypsy-Moth (Lepidoptera, Lymantriidae) Pheromone-Baited Traps. Environ. Entomol. 1988, 17, 764–769. [Google Scholar] [CrossRef]
- Tcheslavskaia, K.S.; Thorpe, K.W.; Brewster, C.C.; Sharov, A.A.; Leonard, D.S.; Reardon, R.C.; Mastro, V.C.; Sellers, P.; Roberts, E.A. Optimization of pheromone dosage for gypsy moth mating disruption. Entomol. Exp. Appl. 2005, 115, 355–361. [Google Scholar] [CrossRef]
- SAS Institute Inc. JMP®, 1989–2020; SAS Institute Inc.: Cary, NC, USA, 2020. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Elkinton, J.; Cardé, R.; Mason, C. Evaluation of time-average dispersion models for estimating pheromone concentration in a deciduous forest. J. Chem. Ecol. 1984, 10, 1081–1108. [Google Scholar] [CrossRef]
- Roberts, E.A.; Ziegler, A.H. Gypsy moth population monitoring and data collection. In Slow the Spread: A National Program to Manage the Gypsy Moth; Tobin, P., Blackbum, L.M., Eds.; USDA Forest Service Northern Research Station: Newtown Square, PA, USA, 2007. [Google Scholar]
- Sharov, A.A.; Liebhold, A.M.; Roberts, E.A. Spread of gypsy moth (Lepidoptera: Lymantriidae) in the central Appalachians: Comparison of population boundaries obtained from male moth capture, egg mass counts, and defoliation records. Environ. Entomol. 1996, 25, 783–792. [Google Scholar] [CrossRef]
- Tobin, P.C.; Sharov, A.A.; Liebhold, A.A.; Leonard, D.S.; Roberts, E.A.; Learn, M.R. Management of the gypsy moth through a decision algorithm under the STS project. Am. Entomol. 2004, 50, 200–209. [Google Scholar] [CrossRef]
- Sharov, A.A.; Liebhold, A.M.; Roberts, E.A. Spatial variation among counts of gypsy moths (Lepidoptera: Lymantriidae) in pheromone-baited traps at expanding population fronts. Environ. Entomol. 1996, 25, 1312–1320. [Google Scholar] [CrossRef]
- Sharov, A.A.; Leonard, D.; Liebhold, A.M.; Roberts, E.A.; Dickerson, W. “Slow the Spread”: A national program to contain the gypsy moth. J. For. 2002, 100, 30–35. [Google Scholar]
- Sharov, A.A.; Leonard, D.; Liebhold, A.M.; Clemens, N.S. Evaluation of preventive treatments in low-density gypsy moth populations using pheromone traps. J. Econ. Entomol. 2002, 95, 1205–1215. [Google Scholar] [CrossRef]
- Onufrieva, K.S.; Onufriev, A.V. Linear relationship between peak and season-long abundances in insects. PLoS ONE 2018, 13, e0193110. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Distance (m) | Number of Males Released | Number of Releases |
|---|---|---|---|
| 2015 | 25 | 200 | 9 |
| 50 | 200 | 3 | |
| 80 | 200 | 3 | |
| 100 | 200 | 3 | |
| 250 | 200 | 5 | |
| 500 | 200 | 5 | |
| 1000 | 200 | 5 | |
| 2016 | 25 | 200 | 8 |
| 100 | 200 | 6 | |
| 150 | 200 | 2 | |
| 180 | 200 | 2 | |
| 200 | 200 | 8 | |
| 250 | 200 | 3 | |
| 300 | 200 | 3 | |
| 2019 | 300 | 50 | 9 |
| 600 | 100 | 9 | |
| 900 | 200 | 9 | |
| 1200 | 300 | 9 | |
| 1500 | 500 | 9 |
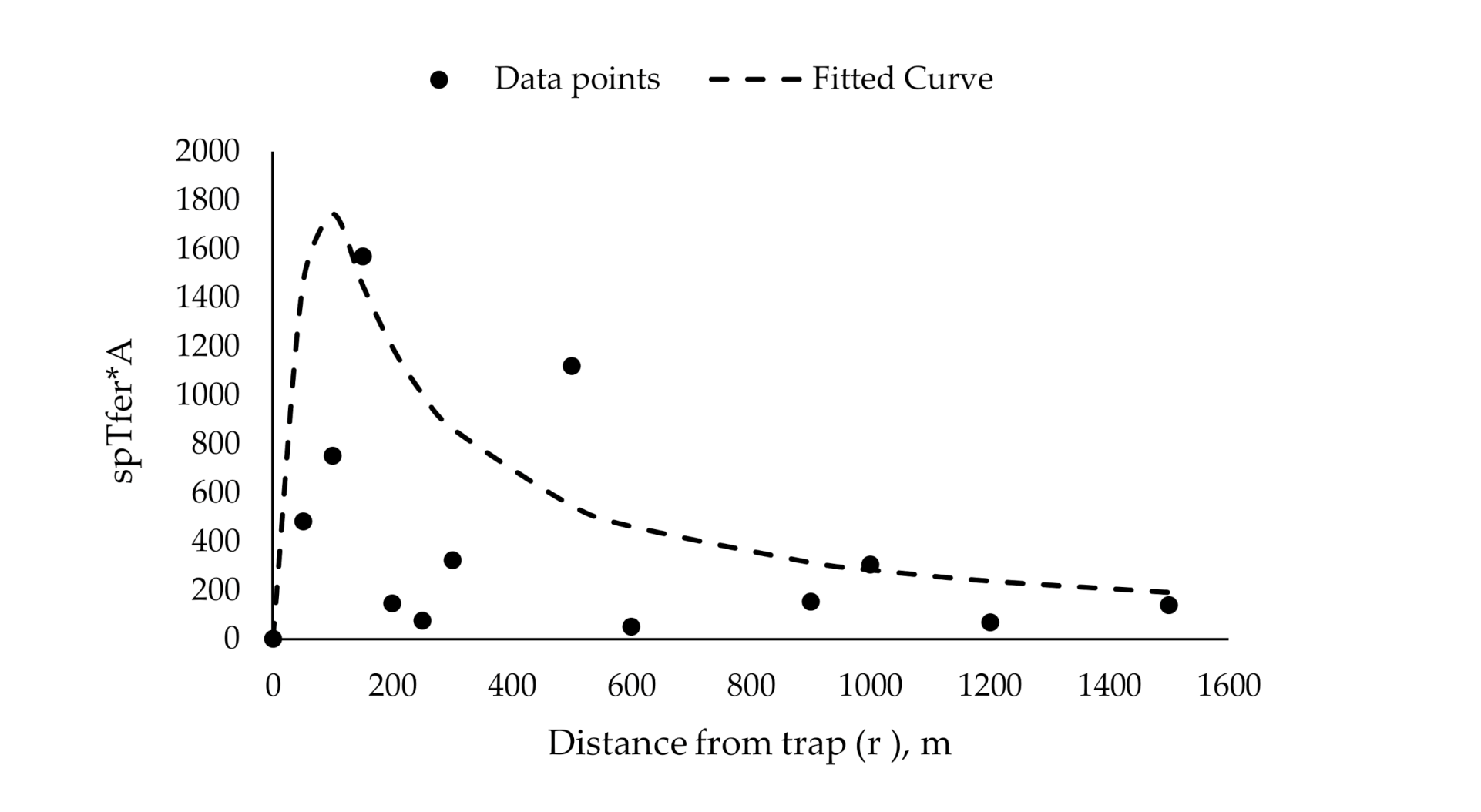
| Release Distance [r] (m) | Annulus Area [A] (m2) | Catch Probability for Each Annulus [spTfer(A)] | Annulus Area × Catch Probability [spTfer(A) × A] |
|---|---|---|---|
| 0 | 0 | 0.37 | 0 |
| 50 | 7854 | 0.24 | 1858 |
| 100 | 23,562 | 0.16 | 3855 |
| 150 | 39,270 | 0.16 | 6283 |
| 200 | 54,978 | 0.01 | 676 |
| 250 | 70,686 | 0.004 | 295 |
| 300 | 86,394 | 0.0008 | 72 |
| 500 | 149,226 | 0.03 | 4477 |
| 600 | 180,642 | 0.003 | 582 |
| 900 | 274,889 | 0.004 | 1222 |
| 1000 | 306,305 | 0.002 | 511 |
| 1200 | 369,137 | 0.007 | 2461 |
| 1500 | 463,385 | 0.01 | 4548 |
| Catch | Lower Bound | Upper Bound | Most Probable Density |
|---|---|---|---|
| 0 | 0 | 5.8 | 0 |
| 1 | 0.04 | 8.8 | 1.6 |
| 2 | 0.38 | 11.5 | 3.2 |
| 3 | 0.98 | 13.9 | 4.8 |
| 4 | 1.7 | 16.3 | 6.4 |
| 5 | 2.6 | 18.5 | 7.9 |
| 6 | 3.5 | 20.7 | 9.5 |
| 7 | 4.5 | 22.9 | 11.1 |
| 8 | 5.5 | 25 | 12.7 |
| 9 | 6.5 | 27.1 | 14.3 |
| 10 | 7.6 | 29.2 | 15.9 |
| 11 | 8.7 | 31.2 | 17.5 |
| 12 | 9.8 | 33.3 | 19 |
| 13 | 11 | 35.3 | 20.6 |
| 14 | 12.2 | 37.3 | 22.2 |
| 15 | 13.3 | 39.3 | 23.8 |
| 16 | 14.5 | 41.2 | 25.4 |
| 17 | 15.7 | 43.2 | 27 |
| 18 | 16.9 | 45.2 | 28.6 |
| 19 | 18.2 | 47.1 | 30.2 |
| 20 | 19.4 | 49 | 31.7 |
| 21 | 20.6 | 51 | 33.3 |
| 22 | 21.9 | 52.9 | 35 |
| 23 | 23.1 | 54.8 | 36.5 |
| 24 | 24.4 | 56.7 | 38.1 |
| 25 | 25.7 | 58.8 | 39.7 |
| 26 | 27 | 60.4 | 41.3 |
| 27 | 28.2 | 62.4 | 42.9 |
| 28 | 29.5 | 64.2 | 44.4 |
| 29 | 30.8 | 66.1 | 46 |
| 30 | 32.1 | 68 | 47.6 |
| 31 | 33.4 | 69.8 | 49.2 |
| 32 | 34.7 | 71.7 | 50.8 |
| 33 | 36.1 | 73.6 | 52.4 |
| 34 | 37.4 | 75.4 | 54 |
| 35 | 38.7 | 77.3 | 55.6 |
| 36 | 40 | 79.1 | 57.1 |
| 37 | 41.4 | 81 | 58.7 |
| 38 | 42.7 | 82.8 | 60.3 |
| 39 | 44 | 84.6 | 61.9 |
| 40 | 45.4 | 86.5 | 63.5 |
| 41 | 46.7 | 88.3 | 65.1 |
| 42 | 48 | 90.1 | 66.7 |
| 43 | 49.4 | 91.9 | 68.3 |
| 44 | 50.7 | 93.8 | 69.8 |
| 45 | 52.1 | 95.6 | 71.4 |
| 46 | 53.5 | 97.4 | 73 |
| 47 | 54.8 | 99.2 | 74.6 |
| 48 | 56.2 | 101 | 76.2 |
| 49 | 57.5 | 102.8 | 77.8 |
| 50 | 58.9 | 104.6 | 79.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onufrieva, K.S.; Onufriev, A.V.; Hickman, A.D.; Miller, J.R. Bounds on Absolute Gypsy Moth (Lymantria dispar dispar) (Lepidoptera: Erebidae) Population Density as Derived from Counts in Single Milk Carton Traps. Insects 2020, 11, 673. https://doi.org/10.3390/insects11100673
Onufrieva KS, Onufriev AV, Hickman AD, Miller JR. Bounds on Absolute Gypsy Moth (Lymantria dispar dispar) (Lepidoptera: Erebidae) Population Density as Derived from Counts in Single Milk Carton Traps. Insects. 2020; 11(10):673. https://doi.org/10.3390/insects11100673
Chicago/Turabian StyleOnufrieva, Ksenia S., Alexey V. Onufriev, Andrea D. Hickman, and James R. Miller. 2020. "Bounds on Absolute Gypsy Moth (Lymantria dispar dispar) (Lepidoptera: Erebidae) Population Density as Derived from Counts in Single Milk Carton Traps" Insects 11, no. 10: 673. https://doi.org/10.3390/insects11100673
APA StyleOnufrieva, K. S., Onufriev, A. V., Hickman, A. D., & Miller, J. R. (2020). Bounds on Absolute Gypsy Moth (Lymantria dispar dispar) (Lepidoptera: Erebidae) Population Density as Derived from Counts in Single Milk Carton Traps. Insects, 11(10), 673. https://doi.org/10.3390/insects11100673