A Unique Population in a Unique Area: The Alcon Blue Butterfly and Its Specific Parasitoid in the Białowieża Forest



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Studies of Adult Demography
2.3. Studies of Premature Individuals in Ant Nests
3. Results
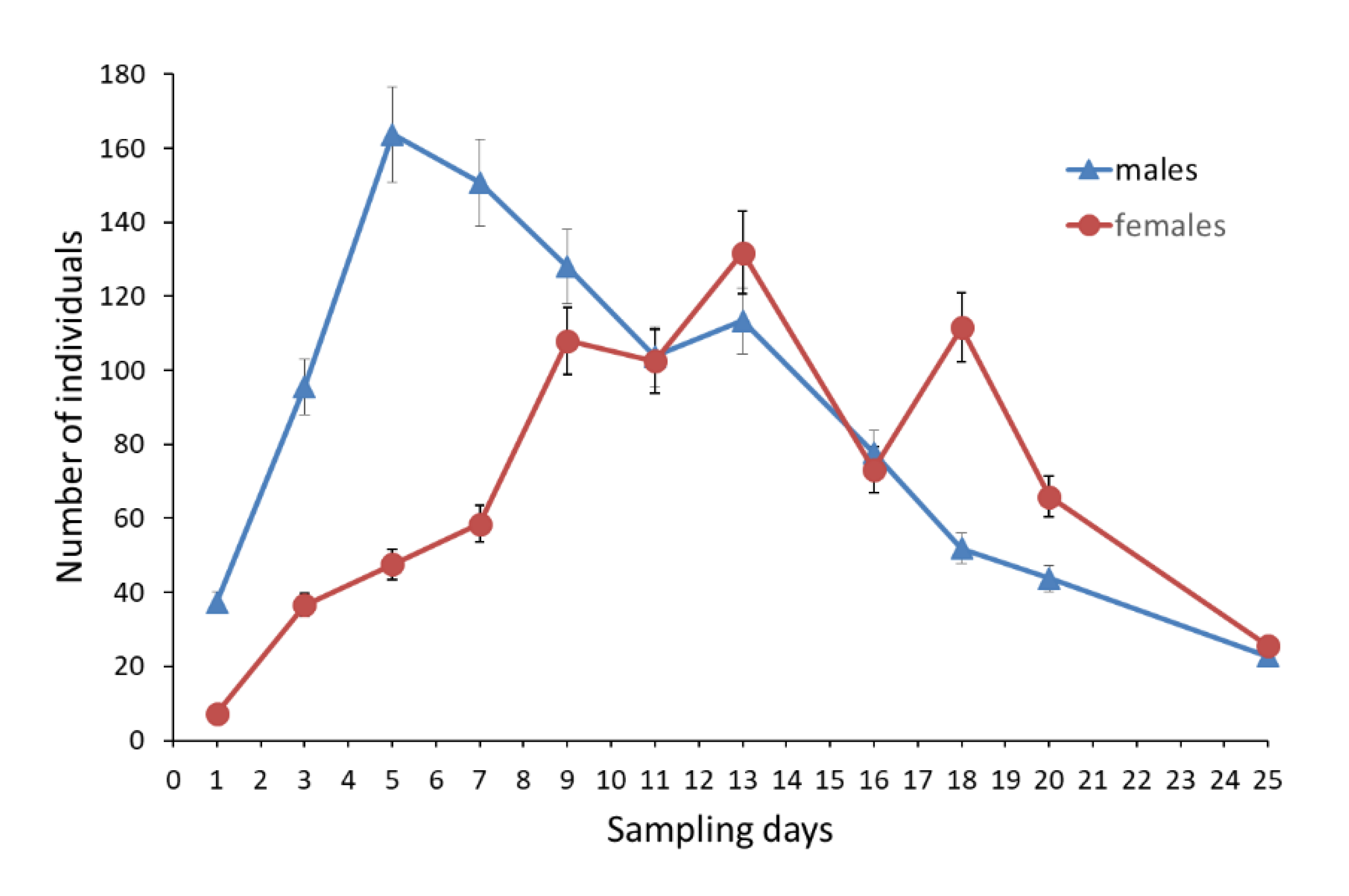
3.1. Adult Demography
3.2. Host Ants and Parasitoids
4. Discussion
4.1. The Alcon Blue Butterfly in the Białowieża Forest
4.2. Adult Demography
4.3. Host Ants and Parasitoids
4.4. Implications for Conservation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fric, Z.; Wahlberg, N.; Pech, P.; Zrzavý, J. Phylogeny and classification of the Phengaris-Maculinea clade (Lepidoptera: Lycaenidae): Total evidence and phylogenetic species concepts. Syst. Entomol. 2007, 32, 558–567. [Google Scholar] [CrossRef]
- Thomas, J.A.; Settele, J. Butterfly mimics of ants. Nature 2004, 432, 283–284. [Google Scholar] [CrossRef]
- Maes, D.; Verovnik, R.; Wiemers, M.; Brosens, D.; Beshkov, S.; Bonelli, S.; Buszko, J.; Cantú-Salazar, L.; Cassar, L.F.; Collins, S.; et al. Integrating national Red Lists for prioritising conservation actions for European butterflies. J. Insect Conserv. 2019, 23, 301–330. [Google Scholar] [CrossRef]
- Thomas, J.A. The ecology and conservation of Maculinea arion and other European species of large blue butterfly. In Ecology and Conservation of Butterflies; Pullin, A.S., Ed.; Chapman and Hall: London, UK, 1995; pp. 180–197. ISBN 978-9401045599. [Google Scholar]
- Settele, J.; Kühn, E.; Thomas, J.A. (Eds.) Studies in the Ecology and Conservation of Butterflies in Europe. Species Ecology along a European Gradient: Maculinea Butterflies as a Model, Volume 2; Pensoft Publishers: Sofia, Bulgaria, 2005; ISBN 978-9546422569. [Google Scholar]
- Thomas, J.A.; Simcox, D.J.; Clarke, R.T. Successful conservation of a threatened Maculinea butterfly. Science 2009, 325, 80–83. [Google Scholar] [CrossRef]
- Barbero, F.; Thomas, J.A.; Bonelli, S.; Balletto, E.; Schönrogge, K. Queen ants make distinctive sounds that are mimicked by a butterfly social parasite. Science 2009, 323, 782–785. [Google Scholar] [CrossRef]
- Thomas, J.A.; Elmes, G.W.; Sielezniew, M.; Stankiewicz–Fiedurek, A.; Simcox, D.J.; Settele, J.; Schönrogge, K. Mimetic host shifts in an endangered social parasite of ants. Proc. R. Soc. Lond. B 2013, 280, 2012233. [Google Scholar] [CrossRef] [Green Version]
- Casacci, L.P.; Bonelli, S.; Balletto, E.; Barbero, F. Multimodal signaling in myrmecophilous butterflies. Front. Ecol. Evol. 2019, 7, 454. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.A.; Elmes, G.W. Higher productivity at the cost of increased host-specificity when Maculinea butterfly larvae exploit ant colonies through trophallaxis rather than by predation. Ecol. Entomol. 1998, 23, 457–464. [Google Scholar] [CrossRef]
- Als, T.D.; Vila, R.; Kandul, N.P.; Nash, D.R.; Yen, S.H.; Hsu, Y.F.; Mignault, A.A.; Boomsma, J.J.; Pierce, N.E. The evolution of alternative parasitic life histories in large blue butterflies. Nature 2004, 432, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Pecsenye, K.; Bereczki, J.; Tihanyi, B.; Toth, A.; Peregovits, L.; Varga, Z. Genetic differentiation among the Maculinea species (Lepidoptera: Lycaenidae) in eastern Central Europe. Biol. J. Linn. Soc. 2007, 91, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Sielezniew, M.; Rutkowski, R.; Ponikwicka-Tyszko, D.; Ratkiewicz, M.; Dziekańska, I.; Švitra, G. Differences in genetic variability between two ecotypes of endangered myrmecophilous butterfly Phengaris (=Maculinea) alcon—The setting of conservation priorities. Insect Conserv. Divers. 2012, 5, 223–236. [Google Scholar] [CrossRef]
- Koubínová, D.; Dincă, V.; Dapporto, L.; Vodă, R.; Suchan, T.; Vila, R.; Alvarez, N. Genomics of extreme ecological specialists: Multiple convergent evolution but no genetic divergence between ecotypes of Maculinea alcon butterflies. Sci. Rep. 2017, 7, 13752. [Google Scholar] [CrossRef] [PubMed]
- Bereczki, J.; Pecsenye, K.; Varga, Z.; Tartally, A.; Tóth, J.P. Maculinea rebeli (Hirschke)—A phantom or reality? Novel contribution to a long-standing debate over the taxonomic status of an enigmatic Lycaenidae butterfly. Syst. Entomol. 2018, 43, 166–182. [Google Scholar] [CrossRef]
- Vanden Broeck, A.; Maes, D.; Kelager, A.; Wynhoff, I.; WallisDeVries, M.F.; Nash, D.R.; Oostermeijer, J.G.B.; Van Dyck, H.; Mergeay, J. Gene flow and effective population sizes of the butterfly Maculinea alcon in a highly fragmented, anthropogenic landscape. Biol. Conserv. 2017, 209, 89–97. [Google Scholar] [CrossRef]
- Maes, D.; Vanreusel, W.; Talloen, W.; Van Dyck, H. Functional conservation units for the endangered Alcon Blue butterfly Maculinea alcon in Belgium (Lepidoptera: Lycaenidae). Biol. Conserv. 2004, 120, 233–245. [Google Scholar] [CrossRef]
- WallisDeVries, M.F. A quantitative conservation approach for the endangered butterfly Maculinea alcon. Conserv. Biol. 2004, 18, 489–499. [Google Scholar] [CrossRef]
- Nowicki, P.; Pepkowska, A.; Kudlek, J.; Skórka, P.; Witek, M.; Settele, J.; Woyciechowski, M. From metapopulation theory to conservation recommendations: Lessons from spatial occurrence and abundance patterns of Maculinea butterflies. Biol. Conserv. 2007, 140, 119–129. [Google Scholar] [CrossRef]
- Nowicki, P.; Bonelli, S.; Barbero, F.; Balletto, E. Relative importance of density-dependent regulation and environmental stochasticity for butterfly population dynamics. Oecologia 2009, 161, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Radchuk, V.; WallisDeVries, M.F.; Schtickzelle, N. Spatially and financially explicit population viability analysis of Maculinea alcon in The Netherlands. PLoS ONE 2012, 7, e38684. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, P.; Deoniziak, K.; Dziekańska, I.; Kostro-Ambroziak, A.; Plazio, E.; Rutkowski, R.; Sielezniew, M. What keeps “living dead” alive: Demography of a small and isolated population of Maculinea (=Phengaris) alcon. J. Insect Conserv. 2019, 23, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Tartally, A.; Thomas, J.A.; Anton, C.; Balletto, E.; Barbero, F.; Bonelli, S.; Bräu, M.; Casacci, L.P.; Csősz, S.; Czekes, Z.; et al. Patterns of host use by brood parasitic Maculinea butterflies across Europe. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaroszewicz, B.; Cholewińska, O.; Gutowski, J.J.; Samojlik, T.; Zimny, M.; Latałowa, M. Białowieża Forest—A relic of the high naturalness of European forests. Forests 2019, 10, 849. [Google Scholar] [CrossRef] [Green Version]
- Krzywicki, M. Fauna Papilionoidea i Hesperioidea (Lepidoptera) Puszczy Białowieskiej. Ann. Zool. 1967, 25, 1–213. [Google Scholar]
- Krzywicki, M. Stan fauny motyli dziennych Lepidoptera, Diurna Puszczy Białowieskiej. Parki Nar. Rez. Przyr. 1986, 7, 69–76. [Google Scholar]
- Buszko, J.; Kokot, A.; Palik, E.; Śliwiński, Z. Motyle większe (Macrolepidoptera) Puszczy Białowieskiej. Parki Nar. Rez. Przyr. 1996, 15, 3–46. [Google Scholar]
- Jaroszewicz, B. Stan zachowania na terenie Puszczy Białowieskiej gatunków motyli z załączników II i IV Dyrektywy Siedliskowej i propozycje działań ochronnych. Parki Nar. Rez. Przyr. 2010, 29, 29–50. [Google Scholar]
- Buszko, J. Poland. In Prime Butterfly Areas in Europe. Priority Sites for Conservation; van Swaay, C.A.M., Warren, M., Eds.; National Reference Centre for Agriculture, Nature and Fisheries, Ministry of Agriculture, Nature Management and Fisheries: Wageningen, The Netherlands, 2003; pp. 399–401. ISBN 978-9072578242. [Google Scholar]
- Thomas, J.A.; Elmes, G.W. Specialized searching and the hostile use of allomones by a parasitoid whose host, the butterfly Maculinea rebeli, inhabits ant nests. Anim. Behav. 1993, 45, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Arnason, A.N.; Schwarz, C.J. Using POPAN-5 to analyse banding data. Bird Study 1999, 46, 157–168. [Google Scholar] [CrossRef]
- White, G.C.; Burnham, K.P. Program MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46, 120–138. [Google Scholar] [CrossRef]
- Schtickzelle, N.; Le Boulenge, E.; Baguette, M. Metapopulation dynamics of the bog fritillary butterfly: Demographic processes in a patchy population. Oikos 2002, 97, 349–360. [Google Scholar] [CrossRef]
- Nowicki, P.; Vrabec, V. Evidence for positive density-dependent emigration in butterfly metapopulations. Oecologia 2011, 167, 657–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osváth-Ferencz, M.; Bonelli, S.; Nowicki, P.; Peregovits, L.; Rákosy, L.; Sielezniew, M.; Kostro-Ambroziak, A.; Dziekańska, I.; Kőrösi, A. Population demography of the endangered large blue butterfly Maculinea arion in Europe. J. Insect Conserv. 2017, 21, 411–422. [Google Scholar] [CrossRef]
- Hurvich, C.M.; Tsai, C. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Nowicki, P.; Witek, M.; Skorka, P.; Settele, J.; Woyciechowski, M. Population ecology of the endangered butterflies Maculinea teleius and M. nausithous and the implications for conservation. Popul. Ecol. 2005, 47, 193–202. [Google Scholar] [CrossRef]
- Bubová, T.; Kulma, M.; Vrabec, V.; Nowicki, P. Adult longevity and its relationship with conservation status in European butterflies. J. Insect Conserv. 2016, 20, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Elmes, G.W.; Thomas, J.A.; Wardlaw, J.C.; Hochberg, M.E.; Clark, R.T.; Simcox, D.J. The ecology of Myrmica ants in relation to the conservation of Maculinea butterflies. J. Insect Conserv. 1998, 2, 67–78. [Google Scholar] [CrossRef]
- Als, T.D.; Nash, D.R.; Boomsma, J.J. Geographical variation in host-ant specificity of the parasitic butterfly Maculinea alcon in Denmark. Ecol. Entomol. 2002, 27, 403–414. [Google Scholar] [CrossRef]
- Czechowski, W.; Radchenko, A.; Czechowska, W.; Vepsäläinen, K. The Ants of Poland with Reference to the Myrmecofauna of Europe; Fauna Poloniae (New Series); Natura optima dux Foundation: Warszawa, Poland, 2012; Volume 4, ISBN 978-83-930773-4-2. [Google Scholar]
- Kulak, A.W. Phengaris alcon (Denis & Schiffermüller, 1775). In Krasnaya Kniga Respubliki Belarusʹ. Zhivotnyye, 4th ed.; Katchanovski, I.M., Nikiforov, M.E., Parfienov, W.I., Eds.; Belorusskaya Entsiklopediya im. Petrusja Brovki: Minsk, Belarus, 2015; pp. 211–212. ISBN 978-985-11-0844-8. [Google Scholar]
- Sielezniew, M.; Dziekańska, I.; Bystrowski, C. Czerwończyk fioletek Lycaena helle (Lepidoptera, Lycaenidae) w polskiej części Puszczy Białowieskiej. Parki Nar. Rez. Przyr. 2017, 36, 87–94. [Google Scholar]
- Karczewska, M.; Michalska-Hejduk, D.; Kucharski, L. The Non-Forest Land Ecosystems of Białowieża National Park; Karczewska, M., Kucharski, L., Eds.; Białowieża National Park: Białowieża, Poland, 2016; pp. 37–139. ISBN 978-8364513169. [Google Scholar]
- Nowicki, P.; Richter, A.; Glinka, U.; Holzschuh, A.; Toelke, U.; Henle, K.; Woyciechowski, M.; Settele, J. Less input same output-simplified approach for population size assessment in Lepidoptera. Popul. Ecol. 2005, 47, 203–212. [Google Scholar] [CrossRef]
- Nowicki, P. Survey precision moderates the relationship between population size and stability. Biol. Conserv. 2017, 212, 310–315. [Google Scholar] [CrossRef]
- Cerrato, C.; Lai, V.; Balletto, E.; Bonelli, S. Direct and indirect effects of weather variability in a specialist butterfly. Ecol. Entomol. 2016, 41, 263–275. [Google Scholar] [CrossRef]
- Baliteau, L. Suivi des insectes à la tourbière de la plaine des Rauzes (Aveyron) et gestion conservatoire de l’entomofaune. Master’s Thesis, Université de Toulouse, Toulouse, France, 2012. [Google Scholar]
- Timuș, N.; Craioveanu, C.; Sitaru, C.; Rus, A.; Rákosy, L. Differences in adult phenology, demography, mobility and distribution in two syntopic ecotypes of Maculinea alcon (cruciata vs. pneumonanthe) (Lepidoptera: Lycaenidae) from Transilvania (Romania). Entomol. Romanica 2013, 18, 21–30. [Google Scholar]
- Meyer-Hozak, C. Population biology of Maculinea rebeli (Lepidoptera: Lycaenidae) on chalk grasslands of Eastern Westphalia (Germany) and implications for conservation. J. Insect Conserv. 2000, 4, 63–72. [Google Scholar] [CrossRef]
- Van Dyck, H.; Regniers, S. Egg spreading in the ant-parasitic butterfly, Maculinea alcon: From individual behaviour to egg distribution pattern. Anim. Behav. 2010, 80, 621–627. [Google Scholar] [CrossRef]
- Osváth-Ferencz, M.; Czekes, Z.; Molnár, G.; Markó, B.; Vizauer, T.Z.; Rákosy, L.; Nowicki, P. Adult population ecology and egg laying strategy in the “cruciata” ecotype of the endangered butterfly Maculinea alcon (Lepidoptera: Lycaenidae). J. Insect Conserv. 2016, 20, 255–264. [Google Scholar] [CrossRef]
- Sielezniew, M.; Stankiewicz-Fiedurek, A.M. Host ant use by Phengaris (=Maculinea) alcon in Poland. Pol. J. Entomol. 2009, 78, 323–335. [Google Scholar]
- Sielezniew, M.; Stankiewicz, A. Simultaneous exploitation of Myrmica vandeli and M. scabrinodis (Hymenoptera: Formicidae) colonies by the endangered myrmecophilous butterfly Maculinea alcon (Lepidoptera: Lycaenidae). Eur. J. Entomol. 2004, 101, 693–696. [Google Scholar] [CrossRef]
- Sielezniew, M.; Bystrowski, C.; Deoniziak, K.; Da Costa, J.M. An unexpected record of Myrmica schencki Emery, 1895 as a secondary host ant of the hygrophilous form of a small and isolated population of the Alcon Blue butterfly Phengaris (=Maculinea) alcon (Denis et Schiffermüller, 1775) (Lepidoptera, Lycaenidae) in NE Poland. Pol. J. Entomol. 2015, 84, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Van Dyck, H.; Talloen, W.; Oostermeijer, J.G.B.; Feenstra, V.; Van der Hidde, A.; Wynhoff, I. Does the presence of ant nests matter for oviposition to a specialized myrmecophilous Maculinea butterfly? Proc. R. Soc. B 2000, 267, 861–866. [Google Scholar] [CrossRef] [Green Version]
- Fürst, M.; Nash, D.R. Host ant independent oviposition in the parasitic butterfly Maculinea alcon. Biol. Lett. 2010, 6, 174–176. [Google Scholar] [CrossRef] [Green Version]
- Wynhoff, I.; Bakker, R.B.; Oteman, B.; Arnaldo, P.S.; Van Langevelde, F. Phengaris (Maculinea) alcon butterflies deposit their eggs on tall plants with many large buds in the vicinity of Myrmica ants. Insect Conserv. Divers. 2015, 8, 177–188. [Google Scholar] [CrossRef]
- Tartally, A.; Somogyi, A.; Révész, T.; Nash, D. Host ant change of a socially parasitic butterfly (Phengaris alcon) through host nest take-over. Insects 2020, 11, 556. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.R.; Stefanescu, C.; van Nouhuys, S. Parasitism of European butterflies (Hesperioidea and Papilionoidea). In Ecology of Butterflies in Europe; Settele, J., Shreeve, T.G., Konvicka, M., van Dyck, H., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 130–156. ISBN 978-0521766975. [Google Scholar]
- Sielezniew, M.; Włostowski, M.; Dziekańska, I. Myrmica schencki (Hymenoptera: Formicidae) as the primary host of Phengaris (Maculinea) arion (Lepidoptera: Lycaenidae) at heathlands in eastern Poland. Sociobiology 2010, 55, 95–106. [Google Scholar]
- Anton, C.; Zeisset, I.; Musche, M.; Durka, W.; Boomsma, J.J.; Settele, J. Population structure of a large blue butterfly and its specialist parasitoid in a fragmented landscape. Mol. Ecol. 2007, 16, 3828–3838. [Google Scholar] [CrossRef] [PubMed]
- Tartally, A. Neotypus melanocephalus (Hymenoptera: Ichneumonidae): The first record of a parasitoid wasp attacking Maculinea teleius (Lycaenidae). Nota Lepidopterol. 2005, 28, 65–67. [Google Scholar]
- Thomas, J.A.; Knapp, J.J.; Akino, T.; Gerty, S.; Wakamura, S.; Simcox, D.J.; Wardlaw, J.C.; Elmes, G.W. Parasitoid secretions provoke ant warfare. Nature 2002, 417, 505–506. [Google Scholar] [CrossRef]
- Timuș, N.; Constantineanu, R.; Rákosy, L. Ichneumon balteatus (Hymenoptera: Ichneumonidae)—A new parasitoid species of Maculinea alcon butterflies (Lepidoptera: Lycaenidae). Entomol. Romanica 2013, 18, 31–35. [Google Scholar]
- Sielezniew, M.; Stankiewicz, A.M.; Górnicki, A. Modraszek Rebela Maculinea rebeli w Przemyślu. Ekologia i perspektywy ochrony populacji. Rocz. Przemys. 2006, 42, 73–88. [Google Scholar]
- Tartally, A. Myrmecophily of Maculinea butterflies in the Carpathian Basin (Lepidoptera: Lycaenidae). Ph.D. Thesis, University of Debrecen, Debrecen, Hungary, 2008. [Google Scholar]
- Tartally, A.; Rodrigues, M.C.; Brakels, P.; Arnaldo, P.S. Myrmica aloba (Hymenoptera: Formicidae) hosts isolated populations of a hoverfly, a butterfly and an ichneumon species in NE-Portugal. J. Insect Conserv. 2013, 17, 851–855. [Google Scholar] [CrossRef]
- Tartally, A.; Koschuh, A.; Varga, Z. The re-discovered Maculinea rebeli (Hirschke, 1904): Host ant usage, parasitoid and initial food plant around the type locality with taxonomical aspects (Lepidoptera, Lycaenidae). Zookeys 2014, 406, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.A.; Clarke, R.T.; Elmes, G.W.; Hochberg, M.E. Population dynamics in the genus Maculinea (Lepidoptera: Lycaenidae). In Insect Population Dynamics: In Theory and Practice; Dempster, J.P., McLean, I.F.G., Eds.; Symposia of the Royal Entomological Society 19; Chapman & Hall: London, UK, 1998; pp. 261–290. ISBN 978-0412832604. [Google Scholar]
- Tartally, A.; Nash, D.R.; Varga, Z.; Lengyel, S. Changes in host ant communities of Alcon Blue butterflies in abandoned mountain hay meadows. Insect Conserv. Divers. 2019, 12, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, M.E.; Elmes, G.W.; Thomas, J.A.; Clarke, R.T. Effects of habitat reduction on the persistence of Ichneumon eumerus (Hymenoptera: Ichneumonidae), the specialist parasitoid of Maculinea rebeli (Lepidoptera: Lycaenidae). J. Insect Conserv. 1998, 2, 59–66. [Google Scholar] [CrossRef]
- Munguira, M.L.; Martín, J. Action Plan for the Maculinea Butterflies in Europe; Nature and Environment, No. 97; Council of Europe Publishing: Strasbourg, France, 1999; ISBN 978-9287139931. [Google Scholar]
- Anton, C.; Musche, M.; Settele, J. Spatial patterns of host exploitation in a larval parasitoid of the predatory Dusky Large Blue, Maculinea nausithous. Basic Appl. Ecol. 2007, 8, 66–74. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecotype of Phengaris alcon | Site | Country | Year(s) | N | Area (ha) | Density (N/ha) | Source |
|---|---|---|---|---|---|---|---|
| Higrophilous (Gentiana pneumonanthe) | Budy | Poland | 2019 | 1460 | 1.71 | 854 | present study |
| Caselette | Italy | 1997–2010 | 1600 | 2.9 | 52–552 | [20,47] | |
| Mottarone | Italy | 2009–2010 | 346–388 | 1.2 | 288–323 | [47] | |
| Rauzes | France | 2004–2007 | 281–1222 | 13 | 22–94 | [48] | |
| Fanatele | Romania | 2010 | 1313 | 40 | 33 | [49] | |
| Bokiny | Poland | 2015 | 40 | 5.47 | 7 | [20] | |
| Xerothermophilous (Gentiana cruciata) | Talesberg | Germany | 1993 | 496 | 0.36 | 1378 | [45] |
| Rimetea | Romania | 2012 | 699 | 0.93 | 752 | [35] | |
| Schmandberg | Germany | 1990 | 108 | 0.5 | 216 | [50] | |
| Sengenberg | Germany | 1990 | 262 | 1.5 | 175 | [50] | |
| Wiedberg | Germany | 1993 | 85 | 0.58 | 147 | [45] | |
| Fanatele Domnesc | Romania | 2010 | 1073 | 40 | 27 | [49] | |
| Rieseler Berg | Germany | 1990 | 38 | 2.25 | 17 | [50] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziekańska, I.; Nowicki, P.; Pirożnikow, E.; Sielezniew, M. A Unique Population in a Unique Area: The Alcon Blue Butterfly and Its Specific Parasitoid in the Białowieża Forest. Insects 2020, 11, 687. https://doi.org/10.3390/insects11100687
Dziekańska I, Nowicki P, Pirożnikow E, Sielezniew M. A Unique Population in a Unique Area: The Alcon Blue Butterfly and Its Specific Parasitoid in the Białowieża Forest. Insects. 2020; 11(10):687. https://doi.org/10.3390/insects11100687
Chicago/Turabian StyleDziekańska, Izabela, Piotr Nowicki, Ewa Pirożnikow, and Marcin Sielezniew. 2020. "A Unique Population in a Unique Area: The Alcon Blue Butterfly and Its Specific Parasitoid in the Białowieża Forest" Insects 11, no. 10: 687. https://doi.org/10.3390/insects11100687
APA StyleDziekańska, I., Nowicki, P., Pirożnikow, E., & Sielezniew, M. (2020). A Unique Population in a Unique Area: The Alcon Blue Butterfly and Its Specific Parasitoid in the Białowieża Forest. Insects, 11(10), 687. https://doi.org/10.3390/insects11100687