The Complete Mitochondrial Genome of Four Hylicinae (Hemiptera: Cicadellidae): Structural Features and Phylogenetic Implications




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Mtgenome Sequences Acquisition and Bioinformatic Analyses
2.3. Phylogenetic Analyses
3. Results and Discussion
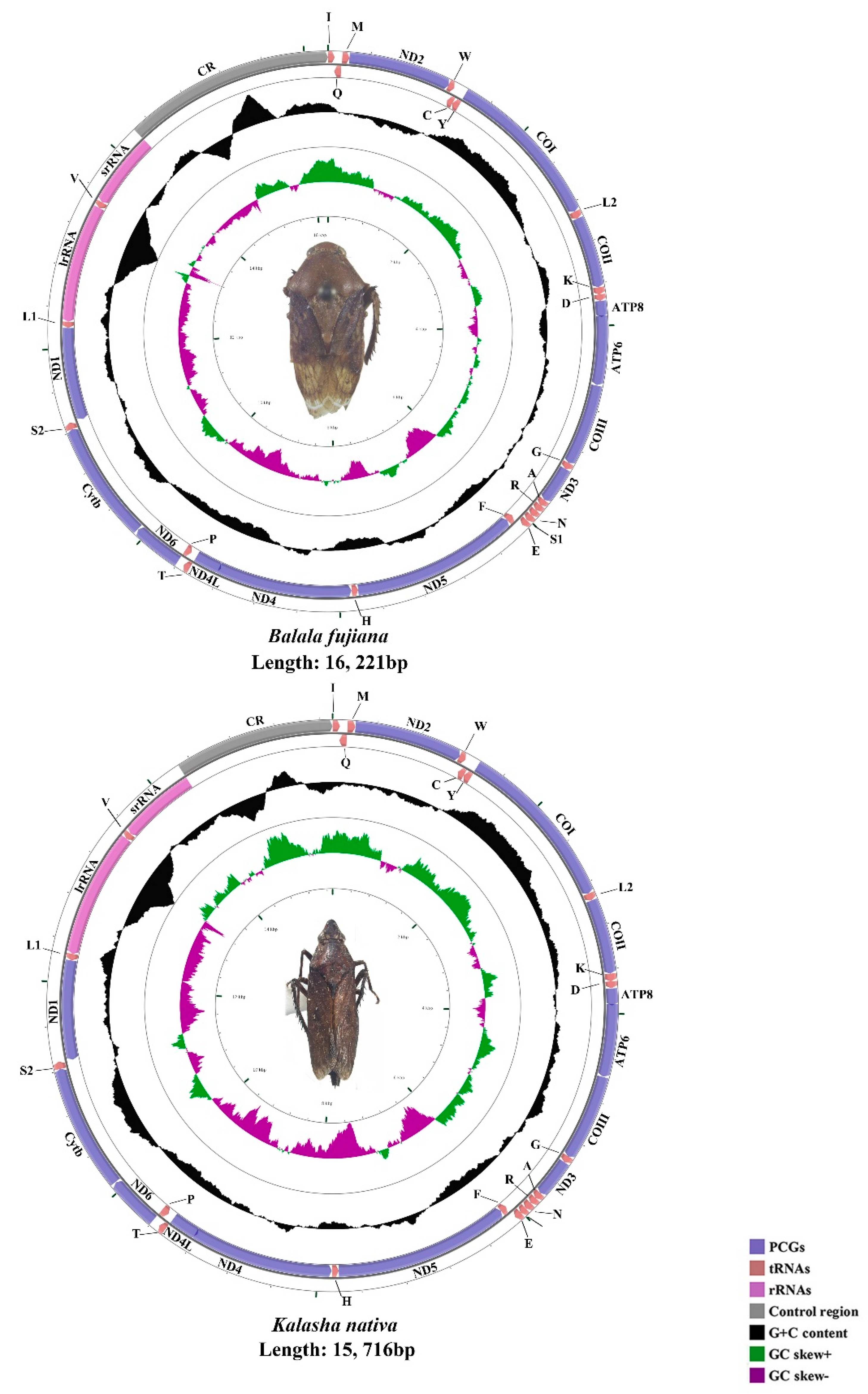
3.1. Genome Organization and Base Composition
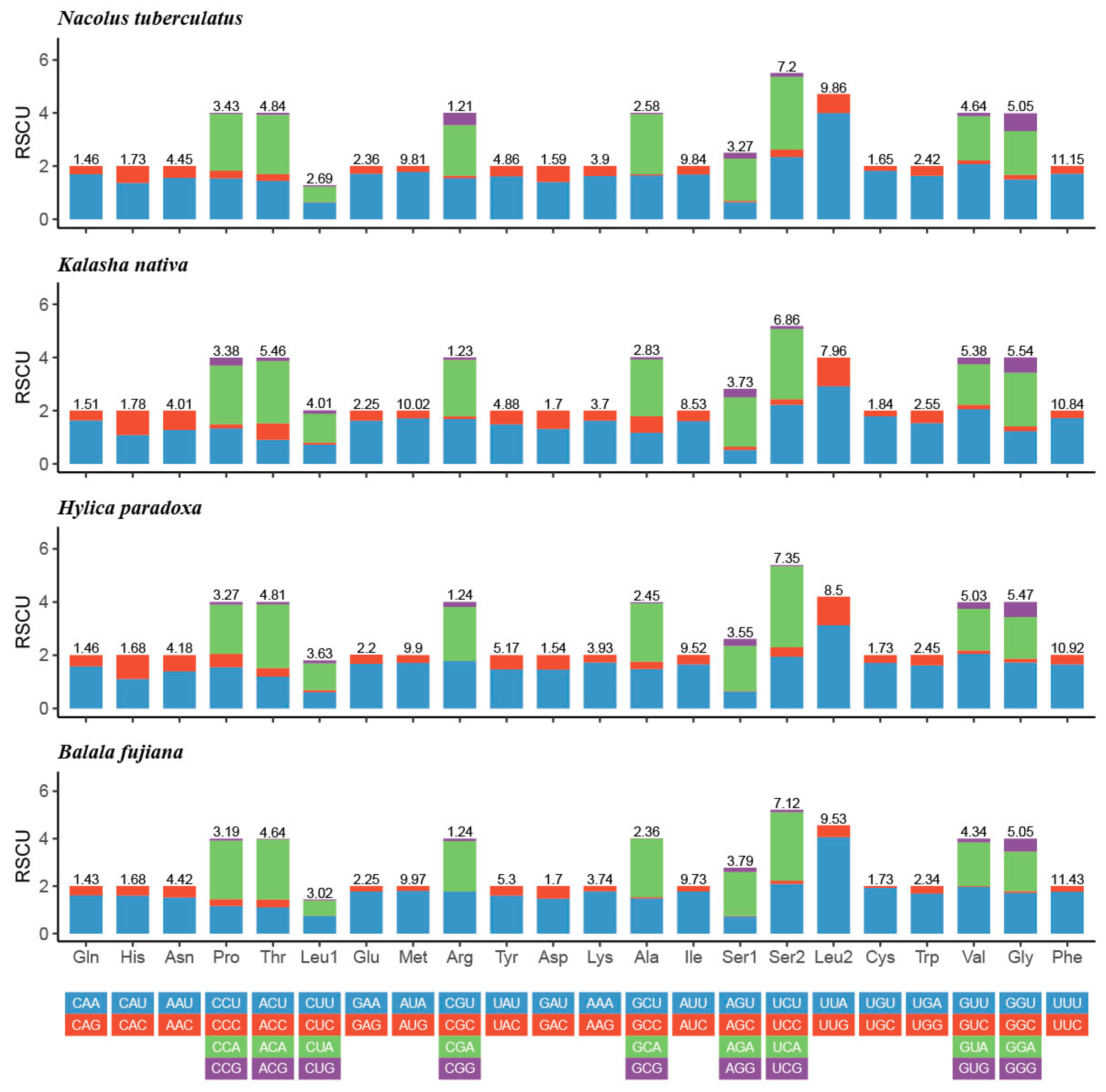
3.2. Protein-Coding Genes and Codon Usage
3.3. Transfer and Ribosomal RNA Genes
3.4. Control Region
3.5. Pi values and Selection Pressures
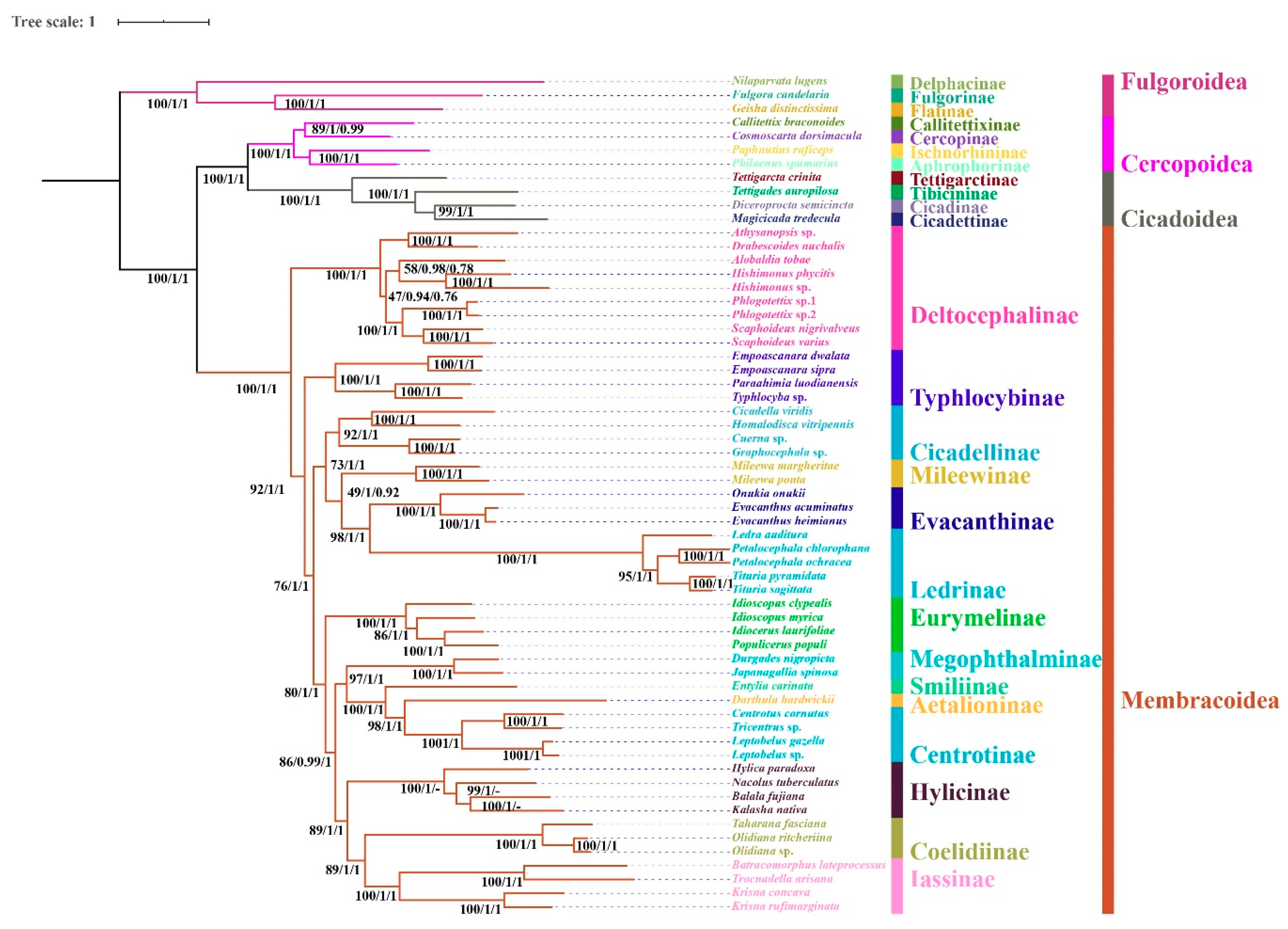
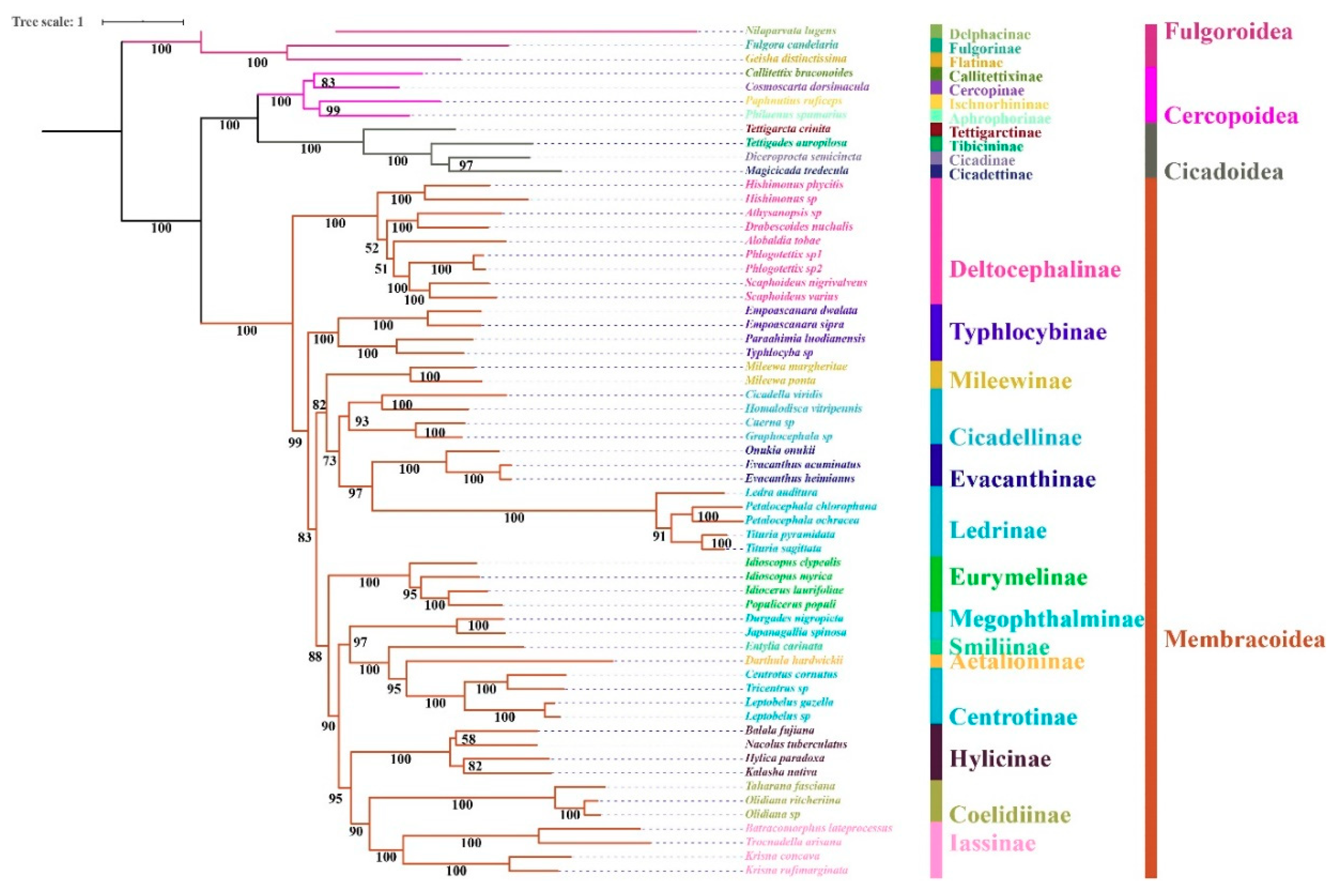
3.6. Phylogenetic Relationships
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dietrich, C.H. Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). Fla. Entomol. 2005, 88, 502–517. [Google Scholar] [CrossRef]
- Camisao, B.M.; Cavichioli, R.R.; Takiya, D.M. Eight new species of Oragua Melichar, 1926 (Insecta: Hemiptera: Cicadellidae) from Amazonas State, Brazil, with description of the female terminalia of Oragua jurua Young, 1977, and new records for the genus. Zootaxa 2014, 3841, 501–527. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.G.A. Classification, morphology and phylogeny of the family Cicadellidae (Rynchota: Homoptera). In Proceedings of the 1st Internal Workshop on Leafhoppers and Planthoppers of Economic Importance; Commonwealth Institute of Entomology: London, UK, 1983; pp. 15–37. [Google Scholar]
- Evans, J.W. A natural classification of leaf-hoppers (Homoptera, Jassoidea). Part 2: Aetalionidae, Hylicidae, Eurymelidae. Trans. R. Entomol. Soc. Lond. 1946, 97, 39–54. [Google Scholar] [CrossRef]
- Dietrich, C.H.; Rakitov, R.A.; Holmes, J.L.; Black, W.C. Phylogeny of the major lineages of Membracoidea (Insecta: Hemiptera: Cicadomorpha) based on 28S rDNA sequences. Mol. Phylogenet. Evol. 2001, 18, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Oman, P.W.; Knight, W.J.; Nielson, M.W. Leafhoppers (Cicadellidae): A Bibliography, Generic Check-list, and Index to the World Literature 1956–1985; CAB International Institute of Entomology: Wallingford, CT, USA, 1990; p. 368. [Google Scholar]
- Tang, J.; Zhang, Y. Review of the oar-head leafhopper genus Nacolus Jacobi (Hemiptera: Cicadellidae: Hylicinae). Zootaxa 2019, 4571, 58–72. [Google Scholar] [CrossRef]
- Metcalf, Z.P. General Catalogue of the Homoptera; Fascicle VI. Cicadelloidea. Part 2. Hylicidae; United States Department of Agriculture-Agricultural Research Service: Washington, DC, USA, 1962; pp. 1–18.
- Kramer, J.P. A review of the African leafhopper genus Wolfella Spinola (Homoptera: Cicadellidae). Trans. Am. Entomol. Soc. 1965, 91, 167–180. [Google Scholar]
- Viraktamath, C.A.; Webb, M.D. A review of the Indian hylicine leafhopper genus Traiguma Dist., with description of a new species (Insecta, Homoptera, Auchenorrhyncha: Cicadellidae). Reichenbachia 1991, 28, 123–133. [Google Scholar]
- Nielson, M.W. The Leafhopper Vectors of Phytopathogenic Viruses (Homoptera, Cicadellidae): Taxonomy, Biology, and Virus Transmission; No. 1382; US Department of Agriculture: Washington, DC, USA, 1968.
- Linnavuori, R. Revisional studies on African leafhoppers (Homoptera Cicadelloidea). Rev. Zool. Bot. Afr. 1972, 86, 195–251. [Google Scholar]
- Kramer, J.P. A review of the Oriental leafhopper genus Sudra Distant (Homoptera: Cicadellidae: Hylicinae). Proc. Biol. Soc. Wash. 1964, 77, 47–52. [Google Scholar]
- Shen, L.; Zhang, Y. A New species of the genus Assiringia Stål (Homoptera: Cicadellidae: Hylicinae) from Tibet, China. Entomotaxonomia 1995, 17, 106–108. [Google Scholar]
- Shen, L.; Zhang, Y. A new species and a new Chinese record of the genus Kalasha Distant (Homoptera: Cicadellidae: Hylicinae). Entomotaxonomia 1995, 17, 185–188. [Google Scholar]
- Shen, L.; Zhang, Y. Two new species of the genus Balala Distant (Homoptera: Cicadellidae: Hylicinae) from China. Entomotaxonomia 1995, 17, 271–275. [Google Scholar]
- Dietrich, C.H.; Allen, J.M.; Lemmon, A.R.; Lemmon, E.M.; Takiya, D.M.; Evangelista, O.; Walden, K.K.O.; Grady, P.G.S.; Johnson, K.P. Anchored hybrid enrichment-based phylogenomics of leafhoppers and treehoppers (Hemiptera: Cicadomorpha: Membracoidea). Insect Syst. Divers 2017, 1, 57–72. [Google Scholar] [CrossRef]
- Boore, L.J. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.-X.; Hewitt, G.M. Insect mitochondrial control region: A review of its structure, evolution and usefulness in evolutionary studies. Biochem. Syst. Ecol. 1997, 25, 99–120. [Google Scholar] [CrossRef]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating molecular evolution into phylogenetic analysis, and a new compilation of conserved polymerase chain reaction primers for animal mitochondrial DNA. Annu. Rev. Ecol. Evol. S 2006, 37, 545–579. [Google Scholar] [CrossRef] [Green Version]
- Gissi, C.; Iannelli, F.; Pesole, G. Evolution of the mitochondrial genome of Metazoa as exemplified by comparison of congeneric species. Heredity 2008, 101, 301–320. [Google Scholar] [CrossRef] [Green Version]
- Shao, R.F.; Dowton, M.; Murrell, A.; Barker, S.C. Rates of gene rearrangement and nucleotide substitution are correlated in the mitochondrial genomes of insects. Mol. Biol. Evol. 2003, 20, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.Y.; Hasegawa, H.; Cooley, J.R.; Simon, C.; Yoshimura, J.; Cai, W.Z.; Sota, T.; Li, H. Mitochondrial genomics reveals shared phylogeographic patterns and demographic history among three periodical cicada species groups. Mol. Biol. Evol. 2019, 36, 1187–1200. [Google Scholar] [CrossRef]
- Li, H.; Leavengood, J.M.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.P.; Zhou, X.G.; Cai, W.Z. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. R. Soc. B Biol. Sci. 2017, 284, 10. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Liang, A.P.; Bu, C.P. A molecular phylogeny of Hemiptera inferred from mitochondrial genome sequences. PLoS ONE 2012, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, S.L.; Lambkin, C.L.; Barker, S.C.; Whiting, M.F. A mitochondrial genome phylogeny of Diptera: Whole genome sequence data accurately resolve relationships over broad timescales with high precision. Syst. Entomol. 2007, 32, 40–59. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Li, H.; Song, F.; Zhao, Y.S.; Wilson, J.J.; Cai, W.Z. Higher-level phylogeny and evolutionary history of Pentatomomorpha (Hemiptera: Heteroptera) inferred from mitochondrial genome sequences. Syst. Entomol. 2019, 44, 810–819. [Google Scholar] [CrossRef]
- Xu, D.; Yu, T.; Zhang, Y. Characterization of the complete mitochondrial genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and their phylogenetic implications. Insects 2020, 11, 534. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, Y. Characterization of two complete mitochondrial genomes of Ledrinae (Hemiptera: Cicadellidae) and phylogenetic analysis. Insects 2020, 11, 609. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, Y. Review of the leafhopper genus Hylica Stal (Hemiptera: Cicadellidae: Hylicinae) with description of one new species. Zootaxa 2018, 4388, 526–536. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, Y. Review of the leafhopper genus Kalasha Distant (Hemiptera: Cicadellidae: Hylicinae). Zootaxa 2019, 4545, 408–418. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, Y. Review of the Oriental leafhopper genus Balala Distant, with new species and new records (Hemiptera: Cicadellidae: Hylicinae). Zootaxa 2020, 4731, 23–42. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Juhling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Putz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.L.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Song, N.; Liang, A. The complete mitochondrial genome sequence of Geisha distinctissima (Hemiptera: Flatidae) and comparison with other hemipteran insects. Acta Biochim. Biophys. Sin. 2009, 41, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.J.; Zhu, W.C.; Rong, X.; Zhang, Y.K.; Ding, X.L.; Liu, J.; Chen, D.S.; Du, Y.; Hong, X.Y. The complete mitochondrial genomes of two rice planthoppers, Nilaparvata lugens and Laodelphax striatellus: Conserved genome rearrangement in Delphacidae and discovery of new characteristics of atp8 and tRNA genes. BMC Genom. 2013, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Bu, C.; Wipfler, B.; Liang, A. Comparative analysis of the mitochondrial genomes of Callitettixini spittlebugs (Hemiptera: Cercopidae) confirms the overall high evolutionary speed of the AT-rich region but reveals the presence of short conservative elements at the tribal level. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liang, A.P. The complete mitochondrial genome of spittlebug Paphnutius ruficeps (Insecta: Hemiptera: Cercopidae) with a fairly short putative control region. Acta Biochim. Biophys. Sin. 2013, 45, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.B.; Beckenbach, A.T. Insect mitochondrial genomics: The complete mitochondrial genome sequence of the meadow spittlebug Philaenus spumarius (Hemiptera: Auchenorrhyncha: Cercopoidae). Genome 2005, 48, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Lukasik, P.; Chong, R.A.; Nazario, K.; Matsuura, Y.; Bublitz, A.C.; Campbell, M.A.; Meyer, M.C.; Van Leuven, J.T.; Pessacq, P.; Veloso, C.; et al. One hundred mitochondrial genomes of cicadas. J. Hered. 2019, 110, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Liang, A.P.; Gao, J.; Zhao, X. Characterization of the complete mitochondrial genome of the treehopper Darthula hardwickii (Hemiptera: Aetalionidae). Mitochondrial DNA Part A 2016, 27, 3291–3292. [Google Scholar] [CrossRef]
- Zhao, X.; Liang, A.P. Complete DNA sequence of the mitochondrial genome of the treehopper Leptobelus gazella (Membracoidea: Hemiptera). Mitochondrial DNA Part A 2016, 27, 3318–3319. [Google Scholar] [CrossRef]
- Song, N.; Cai, W.Z.; Li, H. Insufficient power of mitogenomic data in resolving the auchenorrhynchan monophyly. Zool. J. Linn. Soc. 2018, 183, 776–790. [Google Scholar] [CrossRef]
- Song, N.; Cai, W.; Li, H. Deep-level phylogeny of Cicadomorpha inferred from mitochondrial genomes sequenced by NGS. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Mao, M.; Yang, X.S.; Bennett, G. The complete mitochondrial genome of Entylia carinata (Hemiptera: Membracidae). Mitochondrial DNA Part B Resour. 2016, 1, 662–663. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Li, H.; Dai, R.H. Complete mitochondrial genome of Taharana fasciana (Insecta, Hemiptera: Cicadellidae) and comparison with other Cicadellidae insects. Genetica 2017, 145, 593–602. [Google Scholar] [CrossRef]
- Wang, X.Y.; Wang, J.J.; Fan, Z.H.; Dai, R.H. Complete mitogenome of Olidiana ritcheriina (Hemiptera: Cicadellidae) and phylogeny of Cicadellidae. PeerJ 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.F.; Dai, R.H.; Zhan, H.P.; Qu, L. Complete mitochondrial genome of Drabescoides nuchalis (Hemiptera: Cicadellidae). Mitochondrial DNA Part A 2016, 27, 3626–3627. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Yang, M.F.; Dai, R.H.; Li, H. Complete mitochondrial genome of Evacanthus heimianus (Hemiptera: Cicadellidae: Evacanthinae) from China. Mitochondrial DNA Part B Resour. 2019, 4, 284–285. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.W.; Yang, X.; Li, C.; Song, Y.H. The complete mitochondrial genome of the leafhopper Evacanthus acuminatus (Hemiptera: Cicadellidae: Evacanthinae). Mitochondrial DNA Part B Resour. 2019, 4, 3866–3867. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Zhang, H.; Zhao, T. Insights into the phylogeny of Hemiptera from increased mitogenomic taxon sampling. Mol. Phylogenet. Evol. 2019, 137, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Yang, M.F.; Dai, R.H.; Li, H.; Wang, X.Y. Characterization and phylogenetic implications of the complete mitochondrial genome of Idiocerinae (Hemiptera: Cicadellidae). Int. J. Biol. Macromol. 2018, 120, 2366–2372. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.H.; Wang, J.J.; Yang, M.F. The complete mitochondrial genome of the leafhopper Idioscopus clypealis (Hemiptera: Cicadellidae: Idiocerinae). Mitochondrial DNA Part B Resour. 2018, 3, 32–33. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Wu, Y.F.; Dai, R.H.; Yang, M.F. Comparative mitogenomes of six species in the subfamily Iassinae (Hemiptera: Cicadellidae) and phylogenetic analysis. Int. J. Biol. Macromol. 2020, 149, 1294–1303. [Google Scholar] [CrossRef]
- Li, D.-F.; Dai, R.-H. The complete mitochondrial genome of Tituria pyramidata (Hemiptera: Cicadellidae: Ledrinae) from China. Mitochondrial DNA Part B Resour. 2020, 5, 1757–1758. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Li, D.F.; Li, H.; Yang, M.F.; Dai, R.H. Structural and phylogenetic implications of the complete mitochondrial genome of Ledra auditura. Sci. Rep. 2019, 9, 11. [Google Scholar] [CrossRef]
- He, H.; Yang, M. The mitogenome of Mileewa margheritae (Hemiptera: Cicadellidae: Mileewinae). Mitochondrial DNA Part B Resour. 2020, 5, 3181–3182. [Google Scholar] [CrossRef]
- He, H.; Yang, M. Characterization and phylogenetic analysis of the mitochondrial genome of Mileewa ponta (Hemiptera: Cicadellidae: Mileewinae). Mitochondrial DNA Part B Resour. 2020, 5, 2994–2995. [Google Scholar] [CrossRef]
- Wang, J.J.; Dai, R.H.; Li, H.; Zhan, H.P. Characterization of the complete mitochondrial genome of Japanagallia spinosa and Durgades nigropicta (Hemiptera: Cicadellidae: Megophthalminae). Biochem. Syst. Ecol. 2017, 74, 33–41. [Google Scholar] [CrossRef]
- Tan, C.; Chen, X.; Li, C.; Song, Y. The complete mitochondrial genome of Empoascanara sipra (Hemiptera: Cicadellidae: Typhlocybinae) with phylogenetic consideration. Mitochondrial DNA Part B Resour. 2019, 5, 260–261. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.H.; Yuan, X.W.; Li, C. The mitochondrial genome of Paraahimia luodianensis (Hemiptera: Cicadellidae: Typhlocybinae), a new genus and species from China. Mitochondrial DNA Part B Resour. 2020, 5, 1351–1352. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeier, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; Institute of Electrical and Electronics Engineers (IEEE): New Orleans, LA, USA, 2010; Volume 14, pp. 1–8. [Google Scholar]
- Clary, D.O.; Wolstenholme, D.R. The mitochondrial DNA molecular of Drosophila yakuba: Nucleotide sequence, gene organization, and genetic code. J. Mol. Evol. 1985, 22, 252–271. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.M.; Zhang, C.N.; Dietrich, C.H.; Zhang, Y.L.; Dai, W. Characterization of the complete mitochondrial genomes of Maiestas dorsalis and Japananus hyalinus (Hemiptera: Cicadellidae) and comparison with other Membracoidea. Sci. Rep. 2017, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, M.; Yang, X.S.; Bennett, G. The complete mitochondrial genome of Macrosteles quadrilineatus (Hemiptera: Cicadellidae). Mitochondrial DNA Part B Resour. 2017, 2, 173–175. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.P.; Dietrich, C.H.; Friedrich, F.; Beutel, R.G.; Wipfler, B.; Peters, R.S.; Allen, J.M.; Petersen, M.; Donath, A.; Walden, K.K.O.; et al. Phylogenomics and the evolution of hemipteroid insects. Proc. Natl. Acad. Sci. USA 2018, 115, 12775–12780. [Google Scholar] [CrossRef] [Green Version]
- Skinner, R.K.; Dietrich, C.H.; Walden, K.K.O.; Gordon, E.; Sweet, A.D.; Podsiadlowski, L.; Petersen, M.; Simon, C.; Takiya, D.M.; Johnson, K.P. Phylogenomics of Auchenorrhyncha (Insecta: Hemiptera) using transcriptomes: Examining controversial relationships via degeneracy coding and interrogation of gene conflict. Syst. Entomol. 2020, 45, 85–113. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superfamily | Family/Subfamily | Species | Accession Number | Reference |
|---|---|---|---|---|
| Outgroup | ||||
| Fulgoroidea | Fulgoridae/Fulgorinae | Fulgora candelaria | NC_019576 | [26] |
| Flatidae/Flatinae | Geisha distinctissima | NC_012617 | [42] | |
| Delphacidae/Delphacinae | Nilaparvata lugens | NC_021748 | [43] | |
| Cercopoidea | Cercopidae/Cercopinae | Cosmoscarta dorsimacula | NC_040115 | Unpublished |
| Cercopidae/Callitettixinae | Callitettix braconoides | NC_025497 | [44] | |
| Cercopidae/Ischnorhininae | Paphnutius ruficeps | NC_021100 | [45] | |
| Aphrophoridae/Aphrophorinae | Philaenus spumarius | NC_005944 | [46] | |
| Cicadoidea | Cicadidae/Cicadinae | Diceroprocta semicincta | KM000131 | Unpublished |
| Cicadidae/Cicadettinae | Magicicada tredecula | MH937705 | [24] | |
| Cicadidae/Tibicininae | Tettigades auropilosa | KM000129 | Unpublished | |
| Tettigarctidae/Tettigarctinae | Tettigarcta crinita | MG737758 | [47] | |
| Ingroup | ||||
| Membracoidea | Aetalionidae/Aetalioninae | Darthula hardwickii | NC_026699 | [48] |
| Membracidae/Centrotinae | Leptobelus gazella | NC_023219 | [49] | |
| Membracidae/Centrotinae | Centrotus cornutus | KX437728 | [50] | |
| Membracidae/Centrotinae | Tricentrus sp. | KY039115 | [51] | |
| Membracidae/Centrotinae | Leptobelus sp. | JQ910984 | [25] | |
| Membracidae/Smiliinae | Entylia carinata | NC_033539 | [52] | |
| Cicadellidae/Cicadellinae | Cuerna sp. | KX437741 | [50] | |
| Cicadellidae/Cicadellinae | Graphocephala sp. | KX437740 | [50] | |
| Cicadellidae/Cicadellinae | Cicadella viridis | KY752061 | Unpublished | |
| Cicadellidae/Cicadellinae | Homalodisca vitripennis | NC_006899 | Unpublished | |
| Cicadellidae/Coelidiinae | Taharana fasciana | NC_036015 | [53] | |
| Cicadellidae/Coelidiinae | Olidiana sp. | KY039119 | Unpublished | |
| Cicadellidae/Coelidiinae | Olidiana_ritcheriina | MK738125 | [54] | |
| Cicadellidae/Deltocephalinae | Hishimonus phycitis | KX437727 | [50] | |
| Cicadellidae/Deltocephalinae | Hishimonus sp. | KX437735 | [50] | |
| Cicadellidae/Deltocephalinae | Phlogotettix sp. 1 | KY039135 | [51] | |
| Cicadellidae/Deltocephalinae | Phlogotettix sp. 2 | KX437721 | [50] | |
| Cicadellidae/Deltocephalinae | Scaphoideus varius | KY817245 | [51] | |
| Cicadellidae/Deltocephalinae | Scaphoideus nigrivalveus | KY817244 | [51] | |
| Cicadellidae/Deltocephalinae | Alobaldia tobae | KY039116 | [51] | |
| Cicadellidae/Deltocephalinae | Athysanopsis sp. | KX437726 | [50] | |
| Cicadellidae/Deltocephalinae | Drabescoides nuchalis | NC_028154 | [55] | |
| Cicadellidae/Evacanthinae | Evacanthus heimianus | MG813486 | [56] | |
| Cicadellidae/Evacanthinae | Evacanthus acuminatus | MK948205 | [57] | |
| Cicadellidae/Evacanthinae | Onukia onukii | MK251119 | [58] | |
| Cicadellidae/Eurymelinae | Populicerus populi | NC_039427 | [59] | |
| Cicadellidae/Eurymelinae | Idiocerus laurifoliae | NC_039741 | [59] | |
| Cicadellidae/Eurymelinae | Idioscopus clypealis | NC_0396f42 | [60] | |
| Cicadellidae/Eurymelinae | Idioscopus myrica | MH492317 | [59] | |
| Cicadellidae/Hylicinae | Kalasha nativa | MW218662 | This study | |
| Cicadellidae/Hylicinae | Nacolus tuberculatus | MW218663 | This study | |
| Cicadellidae/Hylicinae | Balala fujiana | MW218661 | This study | |
| Cicadellidae/Hylicinae | Hylica paradoxa | MW218660 | This study | |
| Cicadellidae/Iassinae | Trocnadella arisana | NC036480 | [61] | |
| Cicadellidae/Iassinae | Krisna rufimarginata | NC046068 | [61] | |
| Cicadellidae/Iassinae | Krisna concava | MN577635 | [61] | |
| Cicadellidae/Iassinae | Batracomorphus lateprocessus | MG813489 | [61] | |
| Cicadellidae/Ledrinae | Tituria pyramidata | MN920440 | [62] | |
| Cicadellidae/Ledrinae | Ledra auditura | MK387845 | [63] | |
| Cicadellidae/Ledrinae | Petalocephala ochracea | KX437734 | [50] | |
| Cicadellidae/Ledrinae | Petalocephala chlorophana | MT610899 | [30] | |
| Cicadellidae/Ledrinae | Tituria sagittata | MT610900 | [30] | |
| Cicadellidae/Mileewinae | Mileewa margheritae | MT483998 | [64] | |
| Cicadellidae/Mileewinae | Mileewa ponta | MT497465 | [65] | |
| Cicadellidae/Megophthalminae | Japanagallia spinosa | NC_035685 | [66] | |
| Cicadellidae/Megophthalminae | Durgades nigropicta | NC_035684 | [66] | |
| Cicadellidae/Typhlocybinae | Typhlocyba sp. | KY039138 | [51] | |
| Cicadellidae/Typhlocybinae | Empoascanara dwalata | MT350235 | Unpublished | |
| Cicadellidae/Typhlocybinae | Empoascanara sipra | MN604278 | [67] | |
| Cicadellidae/Typhlocybinae | Paraahimia luodianensis | NC047464 | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Huang, W.; Zhang, Y. The Complete Mitochondrial Genome of Four Hylicinae (Hemiptera: Cicadellidae): Structural Features and Phylogenetic Implications. Insects 2020, 11, 869. https://doi.org/10.3390/insects11120869
Tang J, Huang W, Zhang Y. The Complete Mitochondrial Genome of Four Hylicinae (Hemiptera: Cicadellidae): Structural Features and Phylogenetic Implications. Insects. 2020; 11(12):869. https://doi.org/10.3390/insects11120869
Chicago/Turabian StyleTang, Jiu, Weijian Huang, and Yalin Zhang. 2020. "The Complete Mitochondrial Genome of Four Hylicinae (Hemiptera: Cicadellidae): Structural Features and Phylogenetic Implications" Insects 11, no. 12: 869. https://doi.org/10.3390/insects11120869
APA StyleTang, J., Huang, W., & Zhang, Y. (2020). The Complete Mitochondrial Genome of Four Hylicinae (Hemiptera: Cicadellidae): Structural Features and Phylogenetic Implications. Insects, 11(12), 869. https://doi.org/10.3390/insects11120869